











 text new page (beta)
text new page (beta) Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
Permalink
El conocimiento de la distribución geográfica de las especies es fundamental para comprender los patrones y los procesos que explican la biodiversidad del planeta, así como para proponer estrategias efectivas de conservación (Rosauer & Jetz 2015, Harrison & Noss 2017, Smyčka et al. 2023). Por lo tanto, esta información es clave para plantear hipótesis sobre los procesos ecológicos, geográficos o geológicos que determinan los patrones de riqueza y endemismo de la diversidad biológica (Harrison & Noss 2017).
Los patrones, formas y tamaños de la distribución a diferentes escalas geográficas han sido descritos y analizados para diversos grupos biológicos en América, incluyendo mamíferos (Escalante et al. 2002, Escalante 2013), reptiles (Flores-Villela & García-Vázquez 2014), anfibios (Parra-Olea et al. 2014), árboles tropicales (Pennington & Sarukhán 2005), aves (Navarro-Sigüenza et al. 2014) y angiospermas (Rodríguez et al. 2018). Dentro de las angiospermas, el estudio de la distribución geográfica de familias con alta diversidad taxonómica, como Asteraceae, Fabaceae, Lamiaceae, Orchidaceae y Poaceae, permite inferir los procesos ecológicos, evolutivos y geológicos que han modelado los patrones de la riqueza de especies y endemismo en distintas regiones del continente (Villaseñor et al. 2007, Lenzner et al. 2017, Villaseñor 2018, Estrada-Márquez et al. 2021, Roeble et al. 2024). En particular, Asteraceae ha sido utilizada como indicador de la biodiversidad, principalmente por ser una de las familias de angiospermas más diversas, con aproximadamente 30,000 especies a nivel mundial. Además, sus especies exhiben una notable variación en formas de crecimiento, morfología y distribución geográfica (Villaseñor 2018). Dentro de la familia Asteraceae, destacan géneros como Ageratina Spach (167 especies), Brickellia Elliott (100), Stevia A. Gray (230) y Verbesina L. (325), los cuales presentan un alto número de especies en ambientes templados y áridos del continente americano (Schilling et al. 2015b, Soejima et al. 2017, Hinojosa-Espinosa et al. 2019, Moreira et al. 2023).
Brickellia es uno de los géneros más representativos de la tribu Eupatorieae (Asteraceae), y sus especies se distinguen morfológicamente por presentar cipselas con 8 a 10 costillas, vilano formado por cerdas capilares, corolas relativamente estrechas y un número cromosómico básico de x = 9 (Schilling et al. 2015a). El género es endémico del continente americano y comprende más de 100 especies distribuidas desde el sur de Canadá hasta el norte de Argentina (Schilling et al. 2015a, Villaseñor 2016). México representa un importante centro de diversificación para Brickellia, con alrededor de 85 especies registradas, de las cuales 57 son endémicas (Villaseñor 2018). Las especies del género habitan principalmente en bosques templados como bosque de pino-encino, así como en ambientes áridos y semiáridos, como el matorral xerófilo y el bosque tropical caducifolio (Schilling et al. 2015b).
De acuerdo con Schilling et al. (2015a), Brickellia constituye un grupo monofilético que se originó hace aproximadamente nueve millones de años, durante la transición del Mioceno al Plioceno, con altas tasas de diversificación durante el Pleistoceno. No obstante, la delimitación taxonómica del género ha sido objeto de controversia (King 1987, Turner 1997, Schilling et al. 2013, 2015b). Schilling et al. (2015a) reconstruyeron una filogenia molecular de Brickellia, identificando nueve clados principales. Aunque varios de estos clados habían sido previamente reconocidos como géneros independientes, en su estudio fueron propuestos como secciones taxonómicas dentro del género: Barroetea, Brickellia, Coleosanthus, Gemmipedium, Kuhnia, Leptanthodium, Microphyllae, Phanerostylis y Xerobrickellia.
El género Brickellia ha sido estudiado desde diversos enfoques, incluyendo el taxonómico y filogenético (King 1987, Turner 1997, Schilling et al. 2013, 2015a, 2015b, Morales-Garduño et al. 2023), anatómico y morfológico (Rivera et al. 2019, Morales-Garduño & Villaseñor 2024), ecológico (Kula et al. 2005, Pacheco-Hernández et al. 2021, Duell et al. 2024) y fitoquímico (Palacios-Espinosa et al. 2008, Zhu et al. 2008, Escandón-Rivera et al. 2012, Jiménez-Arellanes et al. 2020, Trevino-Moreno et al. 2023). Sin embargo, hasta la fecha no se han realizado estudios que proporcionen estimaciones detalladas sobre la riqueza de especies y endemismo del género a lo largo de su distribución geográfica. Por lo tanto, el objetivo de este estudio fue analizar la distribución geográfica de las especies de Brickellia, con el fin de identificar áreas o regiones con alta concentración de especies y endemismos, tanto a una escala fina (celdas de 1° latitud y longitud) como a una escala más amplia, basada en provincias biogeográficas.
Materiales y métodos
Base de datos. Se construyó una base de datos para las 109 especies reconocidas del género Brickellia. La lista de especies se elaboró con base en la clasificación propuesta por Schilling et al. (2015a,b) y en el Checklist de las plantas vasculares nativas de México (Villaseñor 2016). Los datos fueron recopilados a partir de la información disponible en ejemplares de herbario y bases de datos en línea. La base de datos resultante contiene información taxonómica (familia, género, epíteto específico y autor) y datos geográficos (país, estado, municipio, localidad, latitud y longitud).
La información sobre las especies de Brickellia presentes en México proviene de la revisión de ejemplares realizada por el especialista en Asteraceae José Luis Villaseñor. Dicha revisión incluyó materiales resguardados en los siguientes herbarios: CHAPA, CIIDIR, CODAGEM, ENCB, FEZA, GUADA, HUAA, HUMO, IBUG, LL, MEXU, MICH, QMEX, RSA, SD, SLPM, TEX y XAL (acrónimos según Thiers 2024). Además, se incorporaron registros documentados en la literatura científica provenientes de ejemplares depositados en colecciones como F, G, GBH, GH, IB, LL, SMU, UAT, UC y US.
Para las especies distribuidas fuera de México, incluyendo Canadá, Estados Unidos, Centroamérica y Sudamérica, se consultó la base de datos del Global Biodiversity Information Facility (GBIF 2024). Los registros obtenidos a través de esta plataforma corresponden a ejemplares físicos o digitales depositados en herbarios como AAU, ALCB, B, BHSC, BRIT, CDS, CR, DINO, EAC, FSU, G, GREE, JBGP, KANU, LAGU, KHD, LOJA, MOSS, MPUC, NCSC, PI, PSO, UB, UBC, UTC, VDB y W (acrónimos según Thiers 2024). Además, se consultaron las bases de datos Tropicos (tropicos.org 2024) y Global Plants (plants.jstor.org 2024) con el propósito de verificar y complementar la información taxonómica y geográfica de algunos registros.
Para detectar y eliminar registros con coordenadas geográficas sospechosas, se utilizó el programa CoordinateCleaner (Zizka et al. 2019). Se consideraron sospechosos aquellos registros con latitud y longitud idénticas o iguales a cero, así como registros ubicados cerca de centroides políticos, capitales nacionales o instituciones de biodiversidad.
Para los ejemplares sin coordenadas geográficas, la longitud y la latitud fueron estimadas a partir de la descripción de las localidades de recolecta, utilizando el programa Google Earth (2024) y siguiendo las recomendaciones metodológicas de García -Milagros & Funk (2010).
La base de datos fue depurada según los criterios de Castillo et al. (2014), corrigiendo o eliminando registros con datos inconsistentes. La distribución geográfica de las especies se verificó mediante la visualización de los registros en QGIS v. 3.36 (qgis.org 2024) y su comparación con las áreas de distribución reportadas en la literatura.
La nomenclatura y sinonimia de las especies fueron revisadas y homologadas según los criterios taxonómicos establecidos en Plants of the World Online (Kew 2024); así como mediante la validación realizada por el especialista en la familia Asteraceae, José Luis Villaseñor.
La distribución de las especies se determinó a partir de la base de datos geográficos utilizando el programa QGIS (qgis.org 2024). La distribución por provincias biogeográficas se realizó con base en los esquemas de regionalización propuestos por Morrone et al. (2022) y Escalante et al. (2021).
Análisis de riqueza de especies y endemismo. La riqueza de especies y el endemismo fueron estimados utilizando celdas y provincias biogeográficas como Unidades Geográficas Operativas (UGOs). Para los análisis por celdas, se generó un mapa con una cuadrícula de 1° de latitud y longitud, que abarcó un total de 840 celdas. A partir de este mapa, se calcularon la riqueza de especies y el endemismo ponderado utilizando el programa Biodiverse v. 4.3 (Laffan et al. 2010). Por otro lado, los análisis por provincias biogeográficas se realizaron a partir de una matriz de presencia-ausencia de las especies, considerando un total de 57 provincias biogeográficas definidas por Morrone et al. (2022) para la región neotropical y por Escalante et al. (2021) para la región neártica (ver mapa en el Material Suplementario 1 Figura S1).
La riqueza de especies se calculó sumando el número total de especies presentes en cada UGO. El endemismo se estimó mediante la métrica de endemismo ponderado (Weighted Endemism, WE), calculada como el inverso del número de UGOs en las que se distribuye cada especie, siguiendo la fórmula WE = ∑(1/Ci), donde Ci representa el número de UGOs en las que se encuentra la especie i. Por lo tanto, las UGOs con valores altos de WE indican una mayor proporción de especies con distribución restringida (Crisp et al. 2001). Los resultados de estos análisis se proyectaron en mapas de celdas y provincias biogeográficas, utilizando el programa QGIS (qgis.org 2024).
Finalmente, se calculó el coeficiente de correlación de Spearman, tanto por celdas como por provincias, con el propósito de evaluar la relación entre los datos de riqueza de especies y el endemismo. Este análisis permitió determinar si las UGOs con alta riqueza de especies también presentan valores altos de endemismo. El análisis se llevó a cabo en el software R (R Core Team 2024), utilizando la función cor.test() para calcular correlación.
Resultados
Base de datos. La base de datos geográficos contiene un total de 11,658 registros únicos, obtenidos tras eliminar duplicados e inconsistencias en las coordenadas (Material Suplementario 2 Tabla S1). Estos registros corresponden a 109 especies del género Brickellia y fueron utilizados para elaborar los mapas de distribución geográfica, así como para realizar los análisis de riqueza de especies y endemismo.
Área de distribución. Los límites geográficos del género Brickellia se extienden desde los 50° N, al sur de Canadá, hasta los -31° S, al norte de Argentina. Las especies se distribuyen en 20 países y 57 provincias biogeográficas a lo largo del continente americano. México alberga el mayor número de especies (91), seguido por Estados Unidos de América (36), Guatemala (7), Nicaragua (6), Honduras (5), Costa Rica y El Salvador (4), y Canadá y Panamá (2). En Argentina, Belice, Bolivia, Brasil, Colombia, Cuba, Ecuador, Paraguay, Perú, República Dominicana y Venezuela se ha registrado una única especie, Brickellia diffusa, la especie más ampliamente distribuida del género, registrada en 17 países desde el norte de México hasta el norte de Argentina.
La información sobre el número de registros, colecciones, países y provincia biogeográficas por especie se presenta en el Material Suplementario 3 Tabla S2. Los mapas de distribución geográfica de cada especie del género están disponibles en el Material Suplementario 1 Figura S2.
Análisis de riqueza de especies y endemismo. México presenta las celdas con los valores más altos de riqueza de especies, destacando celdas con hasta 24 especies, ubicadas en el norte del país (Figura 1A y 1B, Material Suplementario 3 Tabla S3, Material Suplementario 1 Figura S3). De manera general, a lo largo del territorio mexicano se observan celdas que contienen más de 10 especies. El suroeste de Estados Unidos de América también muestra altos niveles de riqueza, con celdas que contienen entre 11 y 15 especies (Figura 1B). En contraste, las celdas con valores de riqueza relativamente bajos (entre 1 y 10 especies) se encuentran principalmente en Centro y Sudamérica, así como en el norte de Estados Unidos de América y Canadá (Figura 1A, Material Suplementario 1 Figura S3, Material Suplementario 3 Tabla S3).
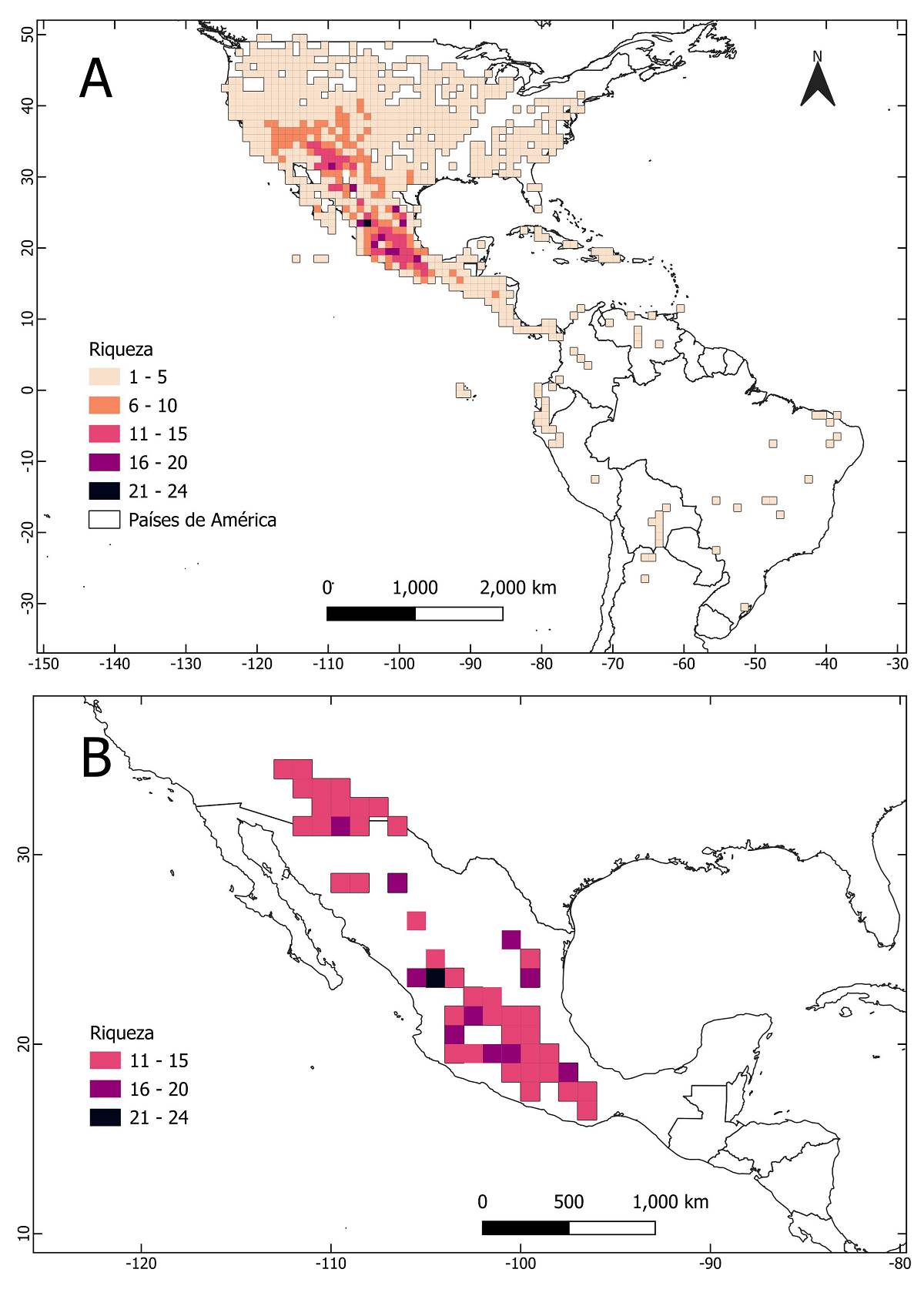
Figura 1 Mapa de la riqueza de especies del género Brickellia en celdas de 1 × 1° de latitud y longitud. A. Riqueza de especies del género en el continente americano. B. Detalle de la riqueza de especies por celdas con los valores más altos en México.
La riqueza de especies del género Brickellia se concentra principalmente en nueve provincias biogeográficas, que en conjunto albergan 102 de las 109 especies registradas (Tabla 1). En contraste, las provincias de Estados Unidos de América, Canadá, Centro y Sudamérica muestran una riqueza relativamente baja, con entre 1 y 15 especies por provincia (Figura 2A y 2B, Material Suplementario 3 Tabla S4). La provincia Chihuahuense presenta la mayor riqueza de especies con 71, seguida por la Sierra Madre Occidental con 45, la Sierra Madre del Sur con 37, las Tierras Bajas del Pacífico con 36 y la Faja Volcánica Transmexicana con 33.
Tabla 1 Valores de riqueza de especies y endemismo ponderado en las principales provincias biogeográficas donde se distribuye el género Brickellia.
| Provincia | Riqueza de especies | Endemismo ponderado (WE) |
|---|---|---|
| Chihuahuense | 71 | 22.30 |
| Sierra Madre Occidental | 45 | 13.24 |
| Sierra Madre del Sur | 37 | 8.23 |
| Tierras Bajas del Pacífico | 36 | 7.04 |
| Faja Volcánica Transmexicana | 33 | 6.28 |
| Sierra Madre Oriental | 32 | 7.59 |
| Navahoniana | 26 | 4.97 |
| Sonorense | 22 | 6.03 |
| Cuenca del Balsas | 22 | 4.95 |
| Veracruzana | 18 | 3.06 |
| Californiana | 14 | 2.62 |
| Kansas | 14 | 2.02 |
| Mohaviana | 12 | 2.25 |
| Tamaulipas | 11 | 1.32 |
| Artemisiana | 10 | 1.59 |
| Baja California | 10 | 4.25 |
| Colorada | 10 | 1.21 |
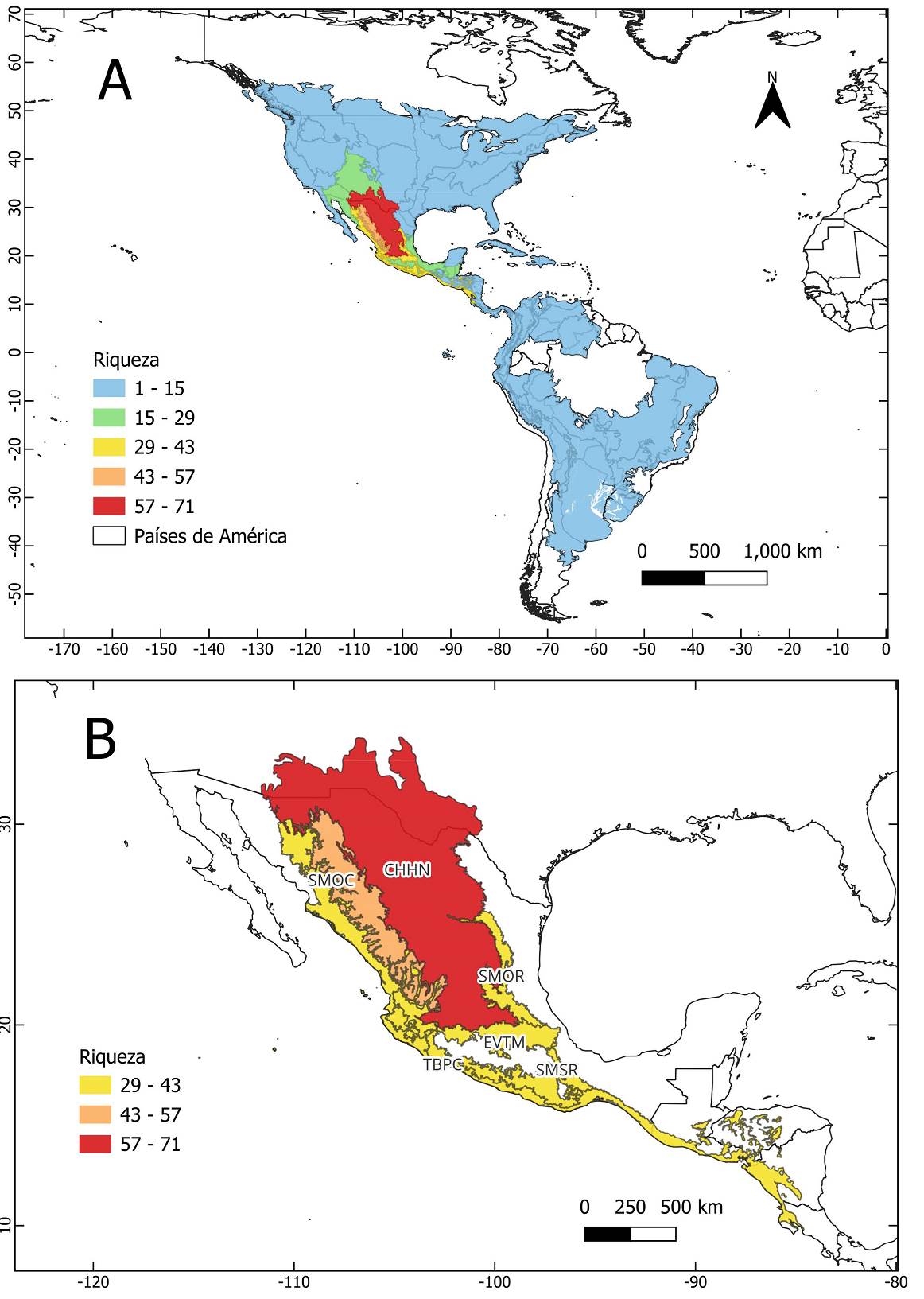
Figura 2 Mapa de la riqueza de especies del género Brickellia por provincias biogeográficas. A. Riqueza de especies por provincias en el continente americano. B. Detalle de la riqueza de especies por provincias con los valores más altos en México. Provincia Chihuahuense (CHHN), Sierra Madre Occidental (SMOC), Sierra Madre Oriental (SMOR), Sierra Madre del Sur (SMSR), Faja Volcánica Transmexicana (EVTM), Tierras Bajas del Pacifico (TBPC).
Al igual que la riqueza de especies, los valores más altos de endemismo ponderado se encuentran en México (Figura 3A y 3B, Material Suplementario 1 Figura S3, Material Suplementario 3 Tabla S3). En contraste, las celdas ubicadas en Centro y Sudamérica, así como en Estados Unidos y Canadá, presentan valores bajos de endemismo ponderado (Figura 3A). La provincia Chihuahuense presenta el valor más alto de endemismo ponderado, seguida por la Sierra Madre Occidental, la Sierra Madre del Sur y la Sierra Madre Oriental (Tabla 1, Figura 4A y 4B). Por el contrario, las provincias de Canadá, Estados Unidos de América, Centro y Sudamérica, así como algunas provincias mexicanas presentan valores relativamente bajos de endemismo ponderado (Figura 4A, Material Suplementario 3 Tabla S4).
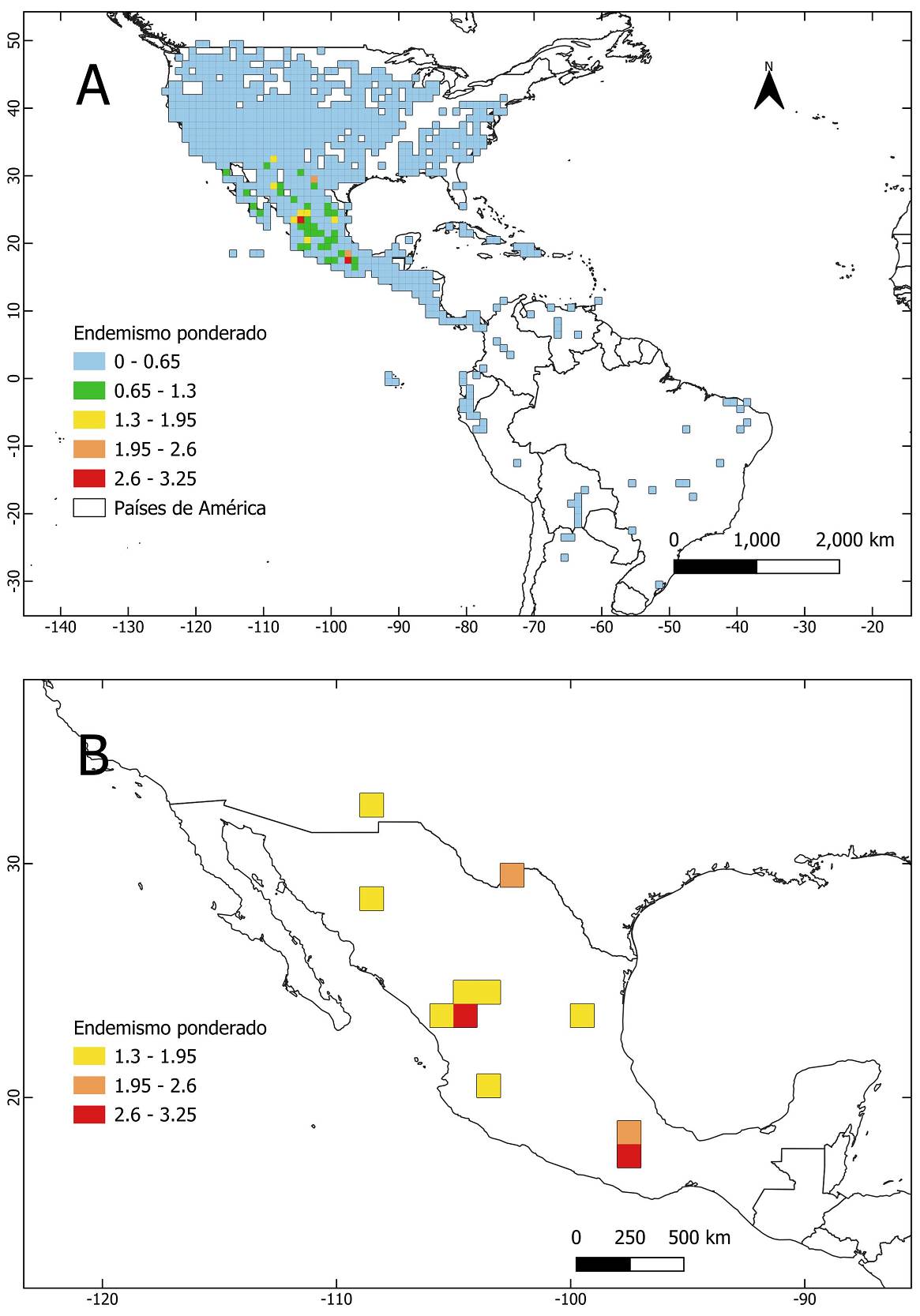
Figura 3 Mapa del endemismo ponderado del género Brickellia por celdas de 1 × 1° de latitud y longitud. A. Endemismo ponderado por celdas del género en el continente americano. B. Detalle del endemismo ponderado por celdas con los valores más altos en México.
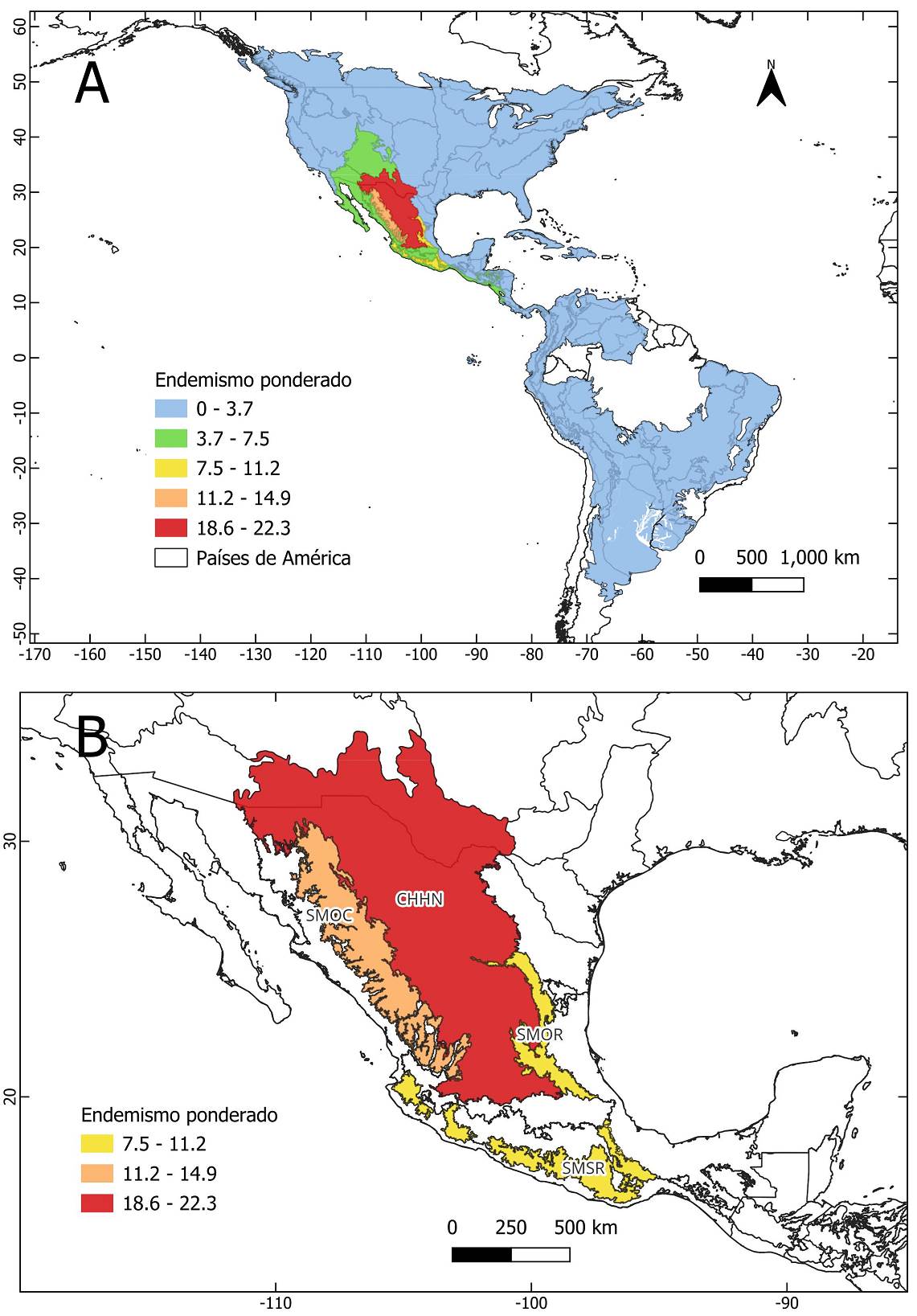
Figura 4 Mapas de endemismo ponderado del género Brickellia por provincias biogeográficas. A. Endemismo ponderado por provincias en el continente americano. B. Detalle del endemismo ponderado por provincias con los valores más altos en México. Provincia Chihuahuense (CHHN), Sierra Madre Occidental (SMOC), Sierra Madre Oriental (SMOR), Sierra Madre del Sur (SMSR).
La correlación de Spearman muestra un valor de rho = 0.8758 para el análisis por celdas y rho = 0.9955 para el análisis por provincias. Estos resultados indican una correlación positiva y significativa entre la riqueza de especies y el endemismo, lo que sugiere que las UGOs con valores altos de riqueza también presentan altos niveles de endemismo ponderado. El valor de p es menor a 2.2e -16 para ambos análisis, lo que confirma que las relaciones observadas son estadísticamente significativas y no son producto del azar.
Discusión
México ha sido ampliamente reconocido como un centro de diversificación para numerosos grupos de plantas, que concentran en su territorio la mayor parte de su riqueza específica. Entre estos se encuentran géneros como Acourtia (Asteraceae), Agave (Asparagaceae), Bursera (Burseraceae), Coryphantha (Cactaceae), Dalea (Fabaceae), Echeveria (Crassulaceae), Mammillaria (Cactaceae), Muhlenbergia (Poaceae) y Verbesina (Asteraceae) (Rzedowski 2005). En este contexto, aunque Brickellia se distribuye desde el sur de Canadá hasta el norte de Argentina, más del 80 % de sus especies se encuentran en México, lo que sugiere que el país constituye un centro clave de diversificación para el género. Por lo tanto, los niveles más altos, tanto de riqueza de especies como de endemismo, se concentran de manera notable en territorio mexicano.
La diversificación de Brickellia ha sido asociada a eventos orogénicos y cambios climáticos ocurridos desde el Mioceno hasta la actualidad, factores que han favorecido el aislamiento geográfico y la especiación de sus poblaciones (Schilling et al. 2015a). Las especies del género se distribuyen en diversos ambientes, aunque su mayor diversidad se concentra en los bosques templados de pino-encino de México. Durante el Pleistoceno, las variaciones climáticas modificaron significativamente la distribución de estos bosques, que lograron persistir en refugios montañosos elevados durante los periodos glaciares (Graham 1993, Mastretta-Yanes et al. 2015). Entonces, estas condiciones favorecieron el aislamiento geográfico y la divergencia genética de las poblaciones de Brickellia, contribuyendo a su alta diversidad de especies y su notable endemismo en México (Schilling et al. 2015a).
La riqueza de especies y endemismo del género Brickellia se concentran principalmente en provincias biogeográficas de México y el sur de los Estados Unidos de América. Las provincias más destacadas en términos de diversidad fueron: Chihuahuense, Sierra Madre Occidental, Sierra Madre del Sur, Tierras Bajas del Pacífico, Faja Volcánica Transmexicana, Sierra Madre Oriental y Navahoniana En particular, la provincia Chihuahuense sobresale por presentar la mayor riqueza de especies y el valor más alto de endemismo. Esta provincia, la más extensa de México, alberga una flora compuesta por aproximadamente 3,500 especies de plantas vasculares, de las cuales más de 1,000 son endémicas (Henrickson & Johnston 2007, Balleza & Villaseñor 2011, Villarreal-Quintanilla et al. 2017). Las familias Cactaceae y Asteraceae destacan por contener el mayor número de especies endémicas en la provincia (Villarreal-Quintanilla et al. 2017). Las especies de las secciones Brickellia, Kuhnia y Microphyllae del género Brickellia se distribuyen principalmente en la provincia Chihuahuense. En particular, las especies de la sección Microphylale habitan predominantemente en ambientes áridos y presentan una serie de características asociadas a dichas condiciones, como hábito arbustivo, hojas sésiles y glandulares, cabezuelas con numerosas flores y un vilano formado por cerdas cortas, lo que refleja su adaptación a estos ambientes (Schilling et al. 2015a,b).
Otras regiones de México, como la Sierra Madre Occidental, la Sierra Madre Oriental, la Faja Volcánica Transmexicana y la Sierra Madre del Sur, también presentan áreas con niveles altos de riqueza y endemismo para el género Brickellia. De acuerdo con Morrone (2019), estos altos niveles de biodiversidad están estrechamente relacionados con la ubicación de estas provincias dentro de la Zona de Transición Mexicana, una región biogeográfica caracterizada por albergar una biota con afinidades tanto neárticas como neotropicales. Diversos estudios han documentado que la mayor riqueza de especies de plantas vasculares en México se concentra principalmente en la Faja Volcánica Transmexicana y la Sierra Madre del Sur (Cruz-Cárdenas et al. 2013, Villaseñor 2016, Rodríguez et al. 2018, Aragón-Parada et al. 2021). En particular, las especies de la sección Barroetea del género Brickellia se distribuyen ampliamente en estas dos regiones; la mayoría son plantas anuales que habitan en bosques templados y bosques tropicales caducifolios (Morales-Garduño et al. 2023). De manera similar, otros grupos de angiospermas también alcanzan su mayor diversidad de especies en la Zona de Transición Mexicana, como Cosmos, Dahlia y Melampodium (Asteraceae) (Vargas-Amado et al. 2013, Carrasco-Ortiz et al. 2019, Stuessy et al. 2025), Tigridieae (Iridaceae) (Munguía-Lino et al. 2015), Lycianthes (Solanaceae) (Anguiano-Constante et al. 2018) y Salvia subg. Calosphace (Lamiaceae) (González-Gallegos et al. 2020).
La provincia Navahoniana, ubicada en el sur de Estados Unidos de América, también presenta altos niveles de riqueza de especies y endemismo para el género Brickellia. Esta región se caracteriza por la presencia de bosques templados y praderas áridas, hábitats que comparten similitudes ecológicas con los presentes en la provincia Chihuahuense (Blair 1950). De acuerdo con Saghatelyan & Escalante (2023)), el sur de Estados Unidos de América es un centro importante de endemismo para plantas, destacando la provincia Navahoniana por sus niveles particularmente elevados de endemismo. Además de su relevancia florística, esta provincia ha sido identificada como una región de interés para el estudio de diversos grupos faunísticos, incluyendo insectos (Baumgardner et al. 1997), mamíferos (Stangl et al. 2012) y parásitos (McAllister et al. 2017).
Las provincias de Sudamérica muestran los valores más bajos tanto en términos de riqueza de especies como de endemismo para el género Brickellia. En esta región, únicamente se ha registrado la presencia de B. diffusa, la especie más ampliamente distribuida del género. Esta especie produce una gran variedad de metabolitos secundarios incluyendo flavonoides (Robinson & King 1980, Norris 1987, Ahmed et al. 1988) y terpenos (Bohlmann et al. 1982), los cuales desempeñan funciones importantes como la atracción de polinizadores y dispersores de semillas, protección contra la radiación UV, señalización en los procesos reproductivos y defensa frente a herbívoros (Calabria et al. 2009). En este sentido, la producción de estos metabolitos podría explicar en parte la amplia distribución geográfica y el éxito ecológico de B. diffusa.
Los valores de correlación de Spearman revelaron una relación positiva entre la riqueza de especies y el endemismo, lo que sugiere que las celdas o provincias con altos niveles de riqueza tienden también a presentar elevados valores de endemismo. Esta correlación ha sido documentada previamente en distintos grupos taxonómicos y regiones geográficas, como en aves de África (Jetz et al. 2004), briofitas de Australia (Stevenson et al. 2012) y la flora Afroalpina tropical (Gehrke & Linder 2014). Sin embargo, investigaciones realizadas a escala global sobre los patrones de riqueza y endemismo en vertebrados han reportado una baja correlación entre ambos patrones, lo que complica la identificación de áreas prioritarias para su conservación (Orme et al. 2005, Lamoreux et al. 2006, Slatyer et al. 2007). La congruencia geográfica entre la riqueza de especies y el endemismo puede ser una herramienta útil para identificar áreas o regiones clave de diversificación de Brickellia, con implicaciones relevantes para su conservación. Sin embargo, estas métricas no consideran la historia evolutiva de los taxones. Por ello, es fundamental llevar a cabo estudios que integren las relaciones filogenéticas del género, con el fin de comprender los procesos históricos y ecológicos que han promovido su diversificación.
En conclusión, la mayor riqueza de especies y endemismo del género Brickellia se concentra en México, principalmente en las provincias Chihuahuense, Sierra Madre Occidental, Sierra Madre del Sur, Tierras Bajas del Pacífico y Faja Volcánica Transmexicana. Este estudio representa un punto de partida para futuras investigaciones que integren la riqueza y el endemismo con las relaciones filogenéticas entre las especies del género, con el fin de profundizar en el entendimiento de su historia evolutiva y orientar estrategias de conservación.
Material suplementario
El material suplementario de este trabajo puede ser consultado aquí: https://doi.org/10.17129/botsci.3705

