










Services on Demand
Journal
Article
 text in
text in  English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista mexicana de fitopatología
On-line version ISSN 2007-8080Print version ISSN 0185-3309
Rev. mex. fitopatol vol.38 n.1 Texcoco Jan. 2020 Epub Nov 27, 2020
https://doi.org/10.18781/r.mex.fit.1911-4
Scientific articles
Morphological and molecular characterization of Fusarium spp. associated to the regional occurrence of wilt and dry bud rot in Agave tequilana
1 Programa de Fitosanidad-Fitopatología; Colegio de Postgraduados, Campus Montecillo, Km 36.5 Carretera México-Texcoco, Montecillo, Texcoco, Estado de México, México. CP 56230
2Programa de Postgrado en Fruticultura; Colegio de Postgraduados, Campus Montecillo, Km 36.5 Carretera México-Texcoco, Montecillo, Texcoco, Estado de México, México. CP 56230
3Laboratorio de Análisis de Riesgo Epidemiológico Fitosanitario (CP-LAN REF). Colegio de Postgraduados, Campus Montecillo, Km 36.5 Carretera México-Texcoco, Montecillo, Texcoco, Estado de México, México. CP 56230
Wilt and dry bud rot of the agave (Agave tequilana var. azul) are diseases of high economic impact for this crop. The implication of Fusarium spp. in both diseases at a regional context was determined in this research. Root and soil samples were collected from 40 commercial plantations located in 13 counties at Los Altos, Jalisco an important cultivation region of agave azul in Mexico; Inoculum charge, as a Fusarium Index was obtained base on colony units (Fusarium vs total fungi) and its relationship with pH and organic matter were estimated from each sampling plantation. A total of 109 isolates were morphologically characterized as Fusarium spp. of which 25 were selected for molecular identification with ITS and EF-1a. The selection considered symptomatology, macro and microscopic characters and prevalence of colony types observed in vitro in Komada, Sabouraud, SNA and CLA media. The cultural and morphological characters evaluated were mycelial coloration, size, shape and septation of macro and microconidia, and length and number of phialides. Five species were associated with wilt and/or dry bud rot within three phylogenetic complexes: F. oxysporum of the Fusarium oxysporum species complex (FOSC) with 56% (46.2 % soil y 66.7 % root) regional representation; F. solani, F. falciforme and Fusarium sp. of Fusarium solani complex (FSSC) (40%); and Fusarium sp. of Fusarium fujikuroi complex (FFSC) (4%). MO and pH correlated inversely with Fusarium Index (r 2=0.68-0.70). It is postulated that wilt and dry bud rot of blue agave constitute a syndrome in which several Fusarium species are associated and parasitically specialized. A single isolate per each species complex was found associated specifically to each or both symptoms. Most isolates were associated to wilting with prevalence of F.oxysporum.
Key words: Blue agave; soil; root; prevalence; EF-1a.
La marchitez y pudrición seca del cogollo del agave (Agave tequilana var. azul) son enfermedades de alto impacto económico para este cultivo. En este trabajo se planteó determinar la implicación de Fusarium spp. en ambas enfermedades bajo un enfoque regional. Se colectaron muestras de raíz y suelo en 40 plantaciones comerciales ubicadas en 13 municipios de Los Altos Jalisco, importante región de cultivo de agave azul en México. De cada plantación de colecta se estimó carga de inóculo mediante un índice de Fusarium obtenido de unidades formadoras de colonias (Fusarium vs hongos totales) y se analizó su relación con pH y materia orgánica. Se obtuvieron 109 aislados caracterizados morfológicamente como Fusarium spp. de los cuales se seleccionaron 25 para identificación molecular con ITS y EF-1a. La selección consideró sintomatología, caracteres macro y microscópicos y prevalencia de tipologías de colonia observadas in vitro en medios Komada, Sabouraud, SNA y CLA. Los caracteres culturales y morfológicos evaluados fueron: coloración micelial, tamaño, forma y septación de macro y microconidios, y longitud y número de fiálides. Se asociaron cinco especies con marchitez y/o pudrición seca ubicadas en tres complejos filogenéticos: F. oxysporum del complejo de especies Fusarium oxysporum (FOSC) con 56% (46.2% suelo y 66.7% raíz) de representatividad regional; F. solani, F. falciforme y Fusarium sp. del complejo Fusarium solani (FSSC) (40%); y Fusarium sp. del complejo Fusarium fujikuroi (FFSC) (4%). MO y pH tuvieron correlación inversamente proporcional con Índice de Fusarium (r 2 = 0.68-0.70). Se postula que la marchitez y pudrición seca del cogollo de agave azul constituyen un síndrome en el cual se asocian y especializan parasíticamente diversas especies de Fusarium. Se encontró un aislado de los tres complejos de Fusarium asociados específicamente a cada tipo de síntoma y la combinación de ellos. La mayoría se asociaron a marchitez con predominancia de F. oxysporum.
Palabras clave: Agave azul; suelo; raíz; prevalencia; EF-1a.
Mexico holds the Tequila Designation of Origin (DOT) since 1997, consisting of 181 counties distributed in the states of Nayarit (8), Guanajuato (7), Tamaulipas (11), Michoacán (30) and Jalisco with 125, of which 21 are at Los Altos region (CRT, 2017). In 2018, the primary production of blue agave (Agave tequilana var. azul) as prime product for tequila processing involved 64,582.76 has equivalent to 206.6 million plants, with 57% of the production concentrated at Los Altos (SIAP, 2018).
In the last 20 years blue agave production has been primarily affected by wilting and dry bud rot (Rubio, 2007; Gómez-Ortiz et al., 2011). Vascular wilt begins with curled tip leaved due to obstruction and plugging of vascular bundles that cause dehydration and progressive tissue death, followed by a change in color from turquoise to opaque green. In advanced disease stages, plants show firm leaf curl, and eventually coriaceous, greater than 50% of the foliage area. In most cases, anchorage is lost due to reduction or total destruction of the root system (Jiménez-González et al., 2017). Etiologically, the association of various organisms, among them; F. oxysporum and F. solani (Rubio, 2007; Vega-Ramos et al., 2013; Ramírez-Ramírez et al., 2017) has been reported. In contrast, plants with dry bud rot are characterized by corrugated lesions of black and yellow-pale color that descend from tip to base of the bud. Initially, rot is restricted to the inner apex of the bud. Unlike gray spot by Cercospora (Coria-Contreras et al., 2019) or soft rot, the bud maintains its rigid texture, without collapse, and has no foul odor. The causal agent of dry rot has also been associated with Fusarium sp. (Gómez-Ortiz, 2011), whereas in soft rot it has been associated to Pectobacterium spp. and Erwinia sp. (Jiménez-Hidalgo et al., 2004; Rincón-Enríquez et al., 2014). Of the three types of bud rot, the dry one has a higher occurrence although lesser than that of wilting (www.sivea.org.mx). In some subregions and farms with high wilt intensity, dry rot can occur in the same plant. Although the official phytosanitary campaign of agave was established in 2013 these diseases repeatedly implicate production inventories due to the absence of effective management methods and abandonment of commercial plantations infested during low-price cyclical periods.
Although classical morphological characterization is important to identify Fusarium spp. (Leslie and Summerell, 2006; Ceja-Torres et al., 2008), increase in diseases and epidemics caused by this genus and its taxonomic complexity justifies the use of molecular techniques for species identification, population structure and phylogeny (Liew et al ., 2016; Herrera-Parra et al., 2017; Parra-Cota et al., 2018; Almaraz-Sánchez et al., 2018). Conventionally, the internal transcribed spacer region (STI) of nuclear ribosomal DNA has been used in Fusarium and fungi in general (O’Donnell et al., 2015). However, the current consensus on Fusarium is that it should be analyzed with genes associated with the elongation factor (EF-1a), the major RNA polymerase I subunit (RPB1) and second largest subunit (RPB2) (O’Donnell et al., 2015). The objective of this research was to identify the Fusarium species obtained from the root and soil of commercial blue agave plantations associated with wilting and dry bud in Los Altos, Jalisco through morphological, epidemiological and molecular techniques in order to know the diversity and regional prevalence that contributes to the establishment of methodological, operational and management strategies that support the Agave Phytosanitary Integral System (SIVEA), the basis of the agave phytosanitary campaign.
Materials And Methods
Study area. The sample collection was carried out in 40 commercial plantations of 3-4 years distributed in 13 counties of Los Altos, Jalisco (Table 1). Plantations were selected using epidemic inductive criteria integrated in the REG-N methodology (Acevedo-Sánchez et al., 2017; Coria-Contreras et al., 2018). Additionally, the predominant planting symptomatology and phytosanitary condition was considered: wilting (M), dry bud rot (P), apparently healthy (AS) and unproductive (IP). The IP plots were chosen because of its high incidence backgrounds, causing their abandonment for commercial agave production.
Sampling method. In plantations with M and P, sampling was carried out around initial foci with 1-4 contiguous plants with 3-4 severity wilt (M) and / or 3-5 dry rot (P) according to classes of diagrammatic scales proposed by Jiménez-González et al. (2017). Five subsamples, two intra-focal and three peripheral, were integrated to form a sample composed of 300 g of soil and roots. The subsample was taken with a metal cylindrical shovel designed to standardize the collection volume at 10 cm from the plant neck and at 35 cm depth. For AS, the same methodology was applied in an internal section of the plantation around apparently healthy plants. In IP plots, the compound sample consisted solely of soil collected in the middle third of the land. Each compound sample was divided for microbiological analysis and physicochemical analysis. The latter was carried out in the soil fertility laboratory of the Colegio de Postgraduados.
Isolates and morphological characterization. Fusarium isolates were obtained from soil samples by dilutions of stock solution (SM), for which 5 g of soil, properly homogenized and grounded were placed in Falcon tubes with 45 mL of sterile distilled water (1:9/p:v). The mixture was homogenized by a stirring equipment for 5 minutes. A 1x100.5 dilution, obtained with 1 mL of SM in 1 mL of sterile distilled water was selected to optimize the colony count. Through a Drigalsky spatula, a 0.1 mL aliquot was distributed in a modified Komada medium, with three Petri dishes as repetitions. The PCNB in Komada was added previous to the sterilization in order to reduce its fungicidal effectiveness and allow a controlled growth of other fungi besides Fusarium. The root tissue (<0.5cm) was disinfested for 1 min with 2% hypochlorite, followed by 1 min in 70% alcohol and three washes with sterile distilled water. Four 1 cm pieces were sown per Petri dish; each piece of tissue was taken as a repetition. Sowings were incubated at 28 °C for 5-7 days. Subsequently, putative Fusarium colonies were transferred to a modified SNA-Agar medium (Leslie and Summerell, 2006). They were incubated for five days and 1-2 monosporic cultures were obtained per isolate.
Table 1. Phytosanitary condition, identification and location of 40 commercial plantations of Agave tequilana 3-4 years old, selected by criteria of epidemic inductivity for regional etiological study of wilting and dry bud rot at Los Altos, Jalisco. Summer 2016.
| Condición Fitosanitaria | Municipio | Latitud | Longitud | Altitud (msnm) | ID Plantación |
|---|---|---|---|---|---|
| Marchitez de planta | Arandas | 20.695398 | -102.326988 | 2082 | A-ARA-M09 |
| 20.850716 | -102.158635 | 1954 | A-ARA-M10 | ||
| Atotonilco A. | 20.580920 | -102.516410 | 1940 | A-ATO-M04 | |
| 20.515050 | -102.612560 | 1624 | A-ATO-M03 | ||
| 20.56022 | -102.53321 | 1911 | A-ATO-M15 | ||
| Jesús María | 20.630650 | -102.213000 | 2132 | A-JM-M11 | |
| 20.613504 | -102.266203 | 2026 | A-JM-M12 | ||
| 20.707930 | -102.122470 | 2235 | A-JM-M14 | ||
| 20.595730 | -102.156940 | 2172 | A-JM-M13 | ||
| Ayotlán | 20.606100 | -102.378590 | 1946 | A-AYO-M06 | |
| 20.601070 | -102.402080 | 1901 | A-AYO-M05 | ||
| San Ignacio C. G. | 20.674480 | -102.505280 | 1949 | A-SICG-M08 | |
| 20.724880 | -102.467340 | 2014 | A-SICG-M07 | ||
| Poncitlán | 20.367180 | -102.836960 | 1533 | A-PON-M01 | |
| Zapotlanejo | 20.575610 | -102.889340 | 1556 | A-ZAP-M02 | |
| Pudrición seca del cogollo | Arandas | 20.683519 | -102.264241 | 2003 | A-ARA-P11 |
| 20.745702 | -102.324358 | 2222 | A-ARA-P05 | ||
| 20.694717 | -102.324106 | 2243 | A-ARA-P10 | ||
| 20.631250 | -102.272260 | 2053 | A-ARA-P12 | ||
| Jesús María | 20.723180 | -102.127880 | 2271 | A-JM-P14 | |
| 20.708100 | -102.122040 | 2238 | A-JM-P13 | ||
| 20.692854 | -102.065208 | 2250 | A-JM-P15 | ||
| Atotonilco A. | 20.588600 | -102.513800 | 1826 | A-ATO-P06 | |
| 20.634495 | -102.435671 | 1957 | A-ATO-P07 | ||
| Ayotlán | 20.581670 | -102.370740 | 1944 | A-AYO-P08 | |
| Poncitlán | 20.36105 | -102.96944 | 1618 | A-PON-P03 | |
| San Ignacio C. G. | 20.723920 | -102.493020 | 2018 | A-SICG-P09 | |
| Tepatitlán M. | 20.649600 | -102.632200 | 1915 | A-TEP-P02 | |
| Yahualica G. G. | 21.113520 | -102.866870 | 1824 | A-YAH-P01 | |
| Zapotlanejo | 20.576420 | -102.889370 | 1869 | A-ZAP-P04 | |
| Improductivos | Cuquío | 21.016096 | -103.006563 | 1810 | A-CUQ-IP01 |
| 20.98254 | -103.02026 | 1860 | A-CUQ-IP02 | ||
| Degollado | 20.42574 | -102.13187 | 1723 | A-DEG-IP07 | |
| 20.46372 | -102.21277 | 1748 | A-DEG-IP08 | ||
| Tepatitlán M. | 20.63825 | -102.90195 | 1751 | A-TEP-IP05 | |
| 20.63964 | -102.638365 | 1904 | A-TEP-IP04 | ||
| San Miguel A. | 20.89801 | -102.40221 | 2049 | A-SMA-IP06 | |
| Acatic | 20.783383 | -102.887821 | 1685 | A-ACA-IP03 | |
| Aparentemente sanos | Jesús María | 20.594201 | -102.156956 | 2176 | A-JM-AS01 |
| Atotonilco A. | 20.662899 | -102.472285 | 1840 | A-ATO-AS02 | |
The characterization of isolates was performed with their growth in Komada, SNA, CLA, PDA and Sabouraud media according to the attribute of interest (Leslie and Summerell, 2006). The analyzed characters were: mycelial coloration; presence or absence of monofialides, polyhalides, klamidospora, conidiogenous cells, and type, shape, septa and size of conidia (Leslie and Summerell, 2006). The observations were made in a Carl Zeiss microscope model Axio Imager A1 with AxioCam ICc1 image analysis system and the AxioVision digital program.
DNA extraction, PCR and sequencing. For the molecular identification a total of 13 monosporic isolates obtained from soil and 12 from the root were selected by their representativeness out of 109 isolates based on in vitro cultural characteristics, morphology, source of isolation, prevalence and regional disease inductivity. Genomic DNA was extracted by the Sambrook and Russell method. The DNA concentration and quality was verified with NanoDrop 2000 (Thermo Fisher Scientific, USA) and the concentration was homogenized at 40 ng µL-1 for the polymerase chain reaction (PCR). The ITS region of the ribosomal DNA was amplified using the published parameters (White et al., 1990) and the elongation factor gene (EF-1a) with the initiators EF1 * (5’-ATG GGT AAG GAR GAV AAG AC) / EF2 * (5’- GGA DGT ACC AGT RAT CAT G). Each PCR reaction for EF-1a consisted of a mixture of 0.8 X of PCR buffer solution, 1.6 mM of MgCl2 (50mM), 0.2 µM of deoxynucleotide triphosphate (dNTP’s), 0.2 pmol of each initiator, 0.5 units of Platinum® Taq DNA polymerase (Invitrogen) and 2.5 µL of DNA (40 ng µL-1) at a final volume of 25 µL. The PCR conditions in thermal cycler (T-100, BioRad) consisted of an initial cycle of denaturation at 94 °C for 5 min, followed by 30 cycles at 94 °C for 30 s, at 58 °C for 40 s and 72 °C for 55 s and a final extension at 72 °C for 7 min. The products of both amplifications were analyzed in 1.5% agarose gel at 90 Volts for 90 min, stained with ethidium bromide and developed with UV photodocumenting light (UVP, BioImaging Systems, Epi Chemi II Darkroom). The amplified fragments of approximately 550 and 750 bp were sequenced by Macrogen Inc., Korea.
Sequence alignment and phylogenetic analysis. The sequences were edited with the SeqAssem program and compared by BLASTn with the Fusarium-ID, Fusarium-MLST and GenBank databases to determine taxon identity based on comparative sequence similarity, expressed as identity percentage. To determine phylogenetic relationships and confirm identification, the sequences obtained were aligned with GenBank reference sequences, using the MUSCLE and Maximum likelihood (ML) algorithm in MEGA7 with 1000 Bootstrap replications. The model selected for ML was Kimura-2 parameters +G. F. dimerum (NRRL 36140) was integrated as an external group. Representative sequences of Fusarium sp. were recorded in Genbank. with accession number MH172303 for the FOSC complex and MH179126 for the FSSC complex.
Regional analysis of Fusarium Index. Samples from soil were only used to evaluate the number of colonies (n) of Fusarium spp. and of total fungi in Komada cultivate medium at seven days of incubation. Each sample had three repetitions consisting of the same number of Petri dishes 90x15 mm. The Fusarium Index (IF) was generated as, IF = (n - colonias Fusarium spp) / (n - hongos totales) where IF ≈ 1.0 indicates higher charge of Fusarium spp. in the soil sample.
Fusarium index analysis with pH and organic matter. Univariate dependence of the Fusarium Index (IF) with pH and organic matter (MO) was analyzed throughout adaptations to the negative exponential models [ln (y) = a-b (x) ] and Gregory [ln (y) = a-bln (x) ]; where y represents the Fusarium Index, x values of pH and MO, a and b are parameters of adjustment and rate, respectively. A total of 28 commercial plots with pH and MO analyzes were included in this analysis.
Results
Morphological and cultural characterization. A total of 109 Fusarium isolates were obtained from 248 in vitro cultures from 40 soil samples (57 isolates) and 32 from root (52). The Komada medium, due to its specificity to Fusarium, was suitable to isolate the fungus from both soil and root and to evaluate mycelial texture and coloration. CLA and SNA were optimal for inducing sporulation and evaluating spore type and septation. These mediums were complementary by differential response between strains (Table 2). In CLA and SNA, macroconidia predominated in a straight to slightly curved and falcate manner, from three to six cells, grouping into two groups: group A included strains with macroconidia of an average length of 26.01 µm (range: 17.12-57.62 µm), and group B with a mean of 33.19 µm (27.16-48.8 µm). One strain presented macroconidia with a length of 76.45 µm (Figure 1A-D). Ellipsoidal microconidia without septa and arranged in false heads in monophialids characterized group A, while in group B they presented a septum. In general, the microconidia had an average length of 10 µm and a range of 3.7-12.4 µm (Figure 1E-F). The size of phialides was decisive to differentiate between strains of group A and B. In the first case short phialides were observed with average length of 23.43 µm (Figure 1G-H) against long phialides of 147.91 µm on average of group B (Figure 1I -J). Chlamydospores were scarce in most strains; however, some isolates of both groups formed them individually or in a chain (Figure 1K). The morphological characteristics found and their variability matched the taxonomic description of the genus Fusarium (Leslie and Summerell, 2006). The 109 isolates were placed at SENASICA-DGSV-CNRF collection. Isolates registered in genbank NCBI were: A115a=MH172303; A129a=MH179126.
Table 2. Culture media selected for isolation, purification, cultural and morphological characterization of Fusarium spp. from 248 cultural sowings, from 40 commercial plantations and 72 samples of soil and blue agave root. Summer, 2016. Los Altos, Jalisco.
| Medio de cultivo x | Objetivo | Características observadas y | |||
|---|---|---|---|---|---|
| Coloración | Esporulación | Esporas | Septación | ||
| Komada | Aislamiento | + | - | - | - |
| SNA | Inducción de esporulación y purificación | - | + | - | - |
| CLA | Caracterización morfológica de esporas | - | + | + | + |
| PDA | Caracterización cultural de colonias | + | - | - | - |
| Sabouraud | Purificación y obtención de cultivos monospóricos | + | - | - | - |
xSNA= Spezieller Nährstoffarmer Agar; CLA= agar hojas de clavel; PDA= agar papa dextrosa.
yColoración= evaluación del color de 109 colonias con 7-10 días de crecimiento; Esporulación= cuantificación cualitativa de la producción de esporas (alta, media y baja); Esporas= presencia o ausencia y tamaño de macro y microconidios; tipo de clamidospora; Septación= cuantificación del número de septos en macro y microconidios.
In Komada, the isolates showed differences in mycelial texture with a smooth and cottony predominance. Ten colors were distinguished by colorimetry, highlighting white, purple, pinkish, ‘salmon’ and ‘melon’ (Figure 1L-P). In 57 root isolates, from eight counties, six colors with greater frequency of white mycelium were determined in 19/57 isolates (33.3%), mainly in wilted plantations (Figure 2A), followed by pinkish isolates. In 52 soil isolates from 11 counties, the entire range of colors with purple dominance was observed in 14/52 isolates (26.92%) in the four phytosanitary conditions (Figure 2C; Table 1 and 3), but mainly associated with dry bud rot plantations (Figure 2B). White was the second color observed (Figure 1L-P; Figure 2C). In Arandas, the greatest diversity of mycelial color was obtained for soil and root isolates, followed by Jesús María with abundance of ‘salmon’, purple and white. In Tepatitlán, the purple-pink combination for soil isolates in unproductive plantations and with dry bud rot stood out. In Jesús María, soil isolates from apparently healthy plantations were distinguished by white and purple mycelia (Figure 2C).
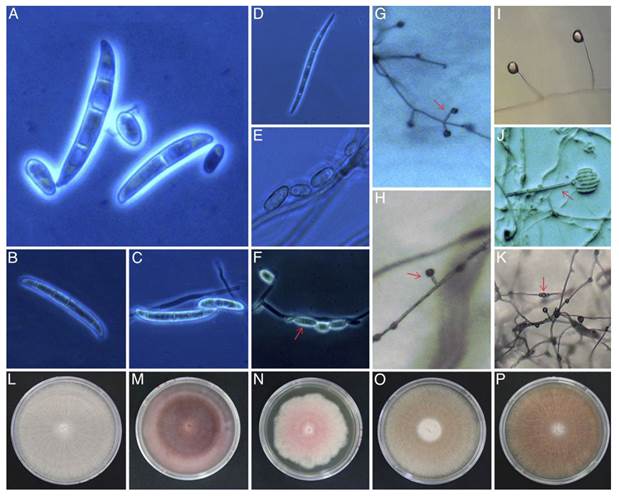
Figure 1. Example of morphological and cultural characteristics used in the characterization of 109 Fusarium isolates obtained from 248 sowings from 40 commercial plantations and 72 samples of soil and blue agave root. Macroconidia (A-D): A-B, slightly curved straight (48.8 and 44.62 µm), 3 to 6 septa, rounded apical cell and basal cell with barely notches. C, Long with a slight curvature of the ventral dorsum (76.45 µm), of 6 septa, sharp apical cell and basal cell barely notched. D, Curved with three septa (32.68 µm), short apical cell, basal cell with distinctive notches. Microconidia (E-F): E, Oval without septa. F, Oval with a septa. Monofiálides (G-J): G-H, Short (23.43 µm). I-J, Long (147.91 µm). Clamidospores (K): In chain. Mycelial coloration in Komada (L-P): L, White. M, Purple. N, Pink. O, ´Melon´. P, ´Salmon´.
Molecular identification and phylogeny. From 25 strains selected for molecular analysis, optimal fragments of 550 bp ITS and 760 bp EF-1a were obtained (Figure 3). The use of STIs allowed the identification of strains at a genus level, but not at a species level because the sequence alignment between members of a species complex or between closely related species was not reliable. The partial sequence analysis of the EF-1a gene along with morphological characterization determined five Fusarium species (two of which unidentified) associated with wilting and dry bud rot, isolated from root and soil, which corresponded to three complexes of species of Fusarium: F. oxysporum of the Fusarium oxysporum species complex (FOSC), with 9 haplotypes, represented morphological group A; F. solani, F. falciforme and Fusarium sp. of the Fusarium solani (FSSC) species complex, with 2, 2 and 1 haplotype, respectively, of morphological group B; and Fusarium sp. of the Fusarium fujikuroi complex (FFSC) with a haplotype. The percentage of coverage and identity for species identification was 98-100%, with an E-value = 0. Representative sequences of the two main complexes, FOSC and FSSC, were registered in Genbank with accession number MH172303 and MH179126, respectively (Table 3).

Figure 2. Diseased plants of selected foci for sampling. A) Plant with symptoms of wilt, severity class 4. Left side, leaf with curl and severe dehydration. B) Plant with symptoms of dry bud rot, severity class 5. Complete bud with advanced drying (arrow). Right side, bud with descending rot and rigid dry texture. C) Diversity of mycelial color of 109 isolates of Fusarium spp. characterized in Komada, obtained from 72 soil and root samples of 40 commercial plantations and 13 counties at Los Altos, Jalisco. Summer 2016.
The phylogenetic analysis based on the EF-1a gene confirmed the taxonomic identification of strains and clearly showed the level of variability and association by species complexes with statistical support of 73-100%. The FSSC strains with the exception of the A185s were grouped with reference sequences. In FOSC only 3 out of 14 isolates were grouped with reference sequences, whereas the rest were grouped into different clades and associated with low reliability values (Figure 3), which suggests greater genetic variation and adaptation of F. oxysporum to regional diversity of the agave production system. The only FFSC isolate, both in GenBank and phylogenetically, was grouped next to F. andiyazi but without corresponding to this species (Figure 3).
Prevalence and symptomatic association. The F. oxysporum (FOSC) complex was the most prevalent with 6/13 (46.15%) soil strains and 8/12 (66.66%) root stains, mainly associated with wilting. The Fusarium solani complex (FSSC) was the second in regional importance with F. falciforme 5/13 (38.46%) strains isolated only from soil, F. solani in 1/13 (7.9%) and 3/12 (25%) strains of soil and root, respectively, and with a strain denoted as Fusarium sp., isolated from the root 1/12 (8.33%). The strains of this complex were associated in the same proportion to plantations with wilting and dry bud rot. Two strains were associated with unproductive sites. Fusarium fujikuroi complex (FFSC) Fusarium sp., which was isolated from the root was associated with wilting symptoms (Figure 4). At the level of haplotypes by species, i.e. intraspecific sequence variants, F. oxysporum had six haplotypes associated solely with wilting, two with dry rot and one to both types of symptoms; F. solani had one associated exclusively to wilt and another that shared both. F. falciforme, although it had a specific haplotype associated with wilting, another haplotype shared both types of symptoms and was found in unproductive soils. The two unidentified Fusarium species at a species level were associated with specific symptoms of wilting or dry rot. F. oxysporum had five repeated haplotypes. Four were associated to different counties and symptoms supporting the highest prevalence of this species.
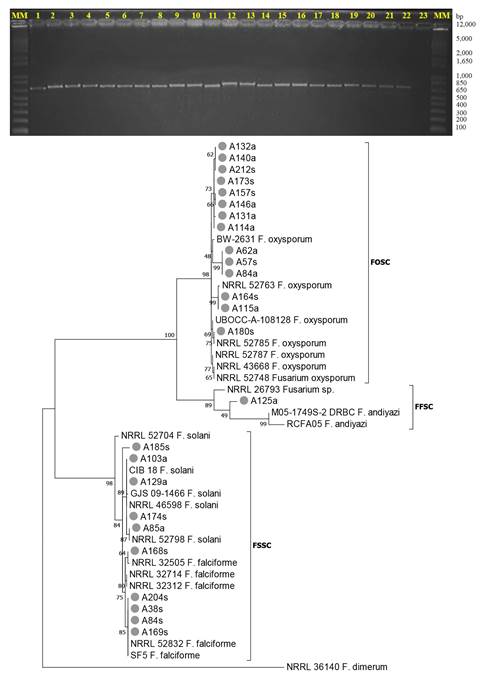
Figure 3. Top image: Example of amplification of PCR products with EF-1a gene (760 bp). Lanes 1-21: Fusarium spp. DNA 21: F. oxysporum (control), 23: White with H2O. MM: 1kb plus Invitrogen molecular weight marker. Bottom: Phylogeny of 25 isolates of Fusarium spp. of soil and root of A. tequilana (marked with ●) based on the EF-1a gene, using Maximum likelihood (ML) and Kimura-2 parameters +G with 1000 Bootstrap repetitions. Text and numeral keys are GenBank reference sequences.
Table 3. Source of isolation and molecular identification, based on the partial sequence of the EF-1a gene, of 25 strains of Fusarium spp. selected from a collection of 109 isolates obtained from soil and root in four phytosanitary conditions and previously characterized morphologically and culturally.
| Fuente de Aislamiento | Especie/Complejox | Condición Fitosanitaria | ID aislado | Coberturay % | Identidady % | Secuencia Referencia |
|---|---|---|---|---|---|---|
| Suelo | F. oxysporum [FOSC] | Marchitez | A173s | 100 | 99 | KP964900 |
| A157s | 100 | 99 | JF740855 | |||
| A164s | 100 | 99 | JF740837 | |||
| A57s | 100 | 98 | JF740855 | |||
| Pudrición | A180s | 100 | 100 | JF740853 | ||
| A212s | 100 | 99 | JF740855 | |||
| F. solani [FSSC] | Pudrición | A174s | 100 | 99 | JF740846 | |
| Fusarium sp. [FSSC] | Pudrición | A185s | 100 | 100 | - | |
| F. falciforme [FSSC] | Improductivo | A38s | 100 | 99 | GU170631 | |
| A84s | 100 | 99 | GU170631 | |||
| Marchitez | A169s | 100 | 99 | GU170631 | ||
| A168s | 100 | 99 | GU170631 | |||
| Pudrición | A204s | 100 | 99 | GU170631 | ||
| Raíz | F. oxysporum [FOSC] | A. sano | A132a | 100 | 99 | JF740855 |
| Marchitez | A114a | 100 | 99 | JF740855 | ||
| A84a | 100 | 98 | JF740855 | |||
| A115aZ | 100 | 99 | JF740837 | |||
| A131a | 100 | 98 | JF740855 | |||
| A140a | 100 | 99 | JF740855 | |||
| A62a | 100 | 98 | JF740855 | |||
| Pudrición | A146a | 100 | 98 | JF740855 | ||
| F. solani [FSSC] | Marchitez | A103a | 100 | 98 | JF740846 | |
| A85a | 100 | 99 | EF452941 | |||
| Pudrición | A129aZ | 100 | 99 | EF452945 | ||
| Fusarium sp. [FFSC] | Marchitez | A125a | 100 | 100 | - | |
xIdentificación taxonómica de aislados de suelo y raíz a nivel de género, especie y complejo mediante caracterización morfológica y molecular con EF.
yCobertura % = Valor de cobertura en porcentaje determinado por nBLAST de NCBI usado en la identificación de cada aislado; Identidad % = Valor de identidad en porcentaje determinado por nBLAST de NCBI.
zAislados seleccionados para registró en el banco de genes de NCBI: A115a=MH172303; A129a=MH179126.
Regional analysis of the Fusarium Index. FI was higher in Atotonilco and Poncitlán with values of 0.95 and 0.91 respectively. These values were associated with plantations with dry rot. In the case of wilting rot the counties with the highest FI were San Ignacio C. G. and Ayotlán with values of 0.48 and 0.45, respectively. The lowest FIs (<0.25) were obtained in Ayotlán, Arandas and Zapotlanejo in the case of dry rot, and in J. María, Poncitlán and Atotonilco for wilt (Figure 5A). In plantations with apparently asymptomatic agaves, FI was 0.49 and 0.42 for Atotonilco and J. María respectively, whereas in unproductive plots of Cuquío, Degollado and Tepatitlán, FI was within the range of 0.37-0.58. These results show the wide distribution of Fusarium and the epidemic risk it represents. Among the total fungi counted in soil samples some genus with antagonistic and suppressive capacity such as Trichoderma spp., Penicillium spp., Bionectria spp. and Aspergillus spp. were found.
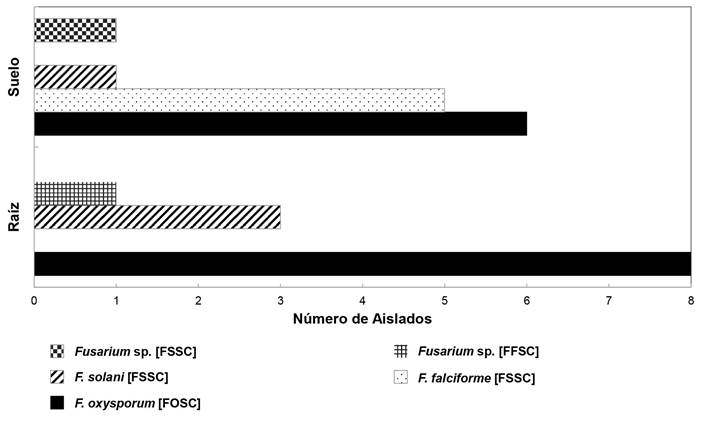
Figure 4. Frequency of species / complex in 25 isolates of Fusarium spp. selected from a collection of 109 isolates obtained from soil and root in four phytosanitary conditions and previously characterized morphologically and culturally. Summer, 2016. Los Altos, Jalisco.
Figure 5. A) Colony Forming Units (UFC) and Fusarium Index obtained from 28 commercial blue agave plantations in four phytosanitary conditions in 11 counties at Los Altos, Jalisco. Error bars indicate standard deviation. Absence of bars indicate a single sample per county (Table 1). Health condition: AS = Apparently healthy, IP = Unproductive, M = Wilt and P = Dry bud rot. B-C) Goodness-of-fit of Fusarium index maximum level to Exponential Negative and Gregory models with respect to soil pH (B) and Organic matter (C). Summer 2016. Los Altos, Jalisco.
FI had an inversely proportional relationship with soil pH and MO (p ≤ 0.02). The FI decremental rate with respect to the increase in pH was adequately estimated with the negative exponential model (b= 0.83, r 2 = 0.71, p <.0001) and Gregory (b = 4.91, r 2 = 0.72, p=0.0098). With these models, it was estimated that pH≤5.5 is inductive to Fusarium (FI≥0.33). The highest Fusarium Index (0.92) was at a pH of 4.6 and the lowest (0.1) in neutral-alkaline soils (6.9-7.7) (Figure 5B). With respect to MO, the estimated rate was slightly better with the negative exponential model (b=0.64, r 2=0.68, p<.0001) than with Gregory (b=1.98, r 2=0.63, p=0.0238). Similarly, it was estimated that MO≤2.5 is inductive to Fusarium (FI≥0.30). Thus, lower FI values ≤0.2 were associated with MO 3.3-5.4 (Figure 5C). Comparing contrasting counties: in Atotonilco, a plantation with a value of FI=0.95 was associated with dry rot and pH=6.18 and MO=3.16; whereas another plantation associated with wilting had that of FI=0.21 with pH=6.95 and MO=5.36. In J. María, a plantation associated with dry rot with a value of FI=0.51 was related to pH=5.57 and MO=2.34, and another one associated to wilt had one of IF=0.11 pH=6.57 and MO=2.48. This suggests that, although FI may be determined by other edaphic-biological factors, it is possible to establish pH:MO thresholds suppressive to Fusarium (Figure 6).
Discussion
The wilt and dry dud rot constitute phytosanitary problems of high productivity risk in blue agave (Agave tequilana) in the DOT region. The phytosanitary management of these diseases is largely limited by etiological and epidemiological restrictions (Rubio, 2007; Ramírez-Ramírez et al., 2017). In previous studies, the etiological approach has been regionally restricted or morphological and classical molecular methods based on ITSs have been emphasized with partial results (Ávila-Miranda et al., 2010; Vega-Ramos et al., 2013; Ramírez-Ramírez et al., 2017). This is the first comprehensive regional work that involves epidemiological inductivity approaches in the selection of 40 commercial plantations in stages of productive stress of 3-4 years. The research included 11 counties at Los Altos, the main agave producing region of Jalisco with historical background of high epidemic intensity on these diseases (Acevedo-Sanchez et al., 2017). Additionally, this work differentiated soil and root samples obtained at the focus and peripheral level from four predominant phytosanitary conditions: wilting, dry rot, seemingly healthy plantations and unproductive plots. The objective was to maximize and minimize regional and intra-plot variability, respectively, in the morphological, cultural and molecular characterization of Fusarium isolates associated to the agave. The conception of this approach considers previous evidence of the implication of F. oxysporum and F. solani on wilting (Ávila-Miranda et al., 2010; Vega-Ramos et al., 2013; Ramírez-Ramírez et al., 2017) and Fusarium sp. on dry bud rot (Gómez-Ortiz et al., 2011) and was conceived assuming implication of a Fusarium species complex with contrasting interspecific parasitic fitness both, at regional and symptom level. This work demonstrated the association of five species with symptoms of wilting and / or dry rot located in three phylogenetic complexes: F. oxysporum of the Fusarium oxysporum species complex (FOSC) with 56% regional representativeness (46.2% soil y 66.7% root); F. solani, F. falciforme and Fusarium sp. of the Fusarium solani (FSSC) complex (40%); and Fusarium sp. of the Fusarium fujikuroi complex (FFSC) (4%). For the first time, the association of F. falciforme is reported, which was the only species associated to plantations with wilting, dry bud rot and unproductive condition, and two species still unidentified, one of them belonging to a complex without previously reported species in agave (FFSC). Root and soil isolates allowed analyze the results consistence due to the fungus association directly with host tissue. F. falciforme and Fusarium sp. (FSSC) were the only ones isolated only from soil. However, the former has already been found in aerial agave tissue in other Jalisco regions (CP-LANREF. Unpublished data).
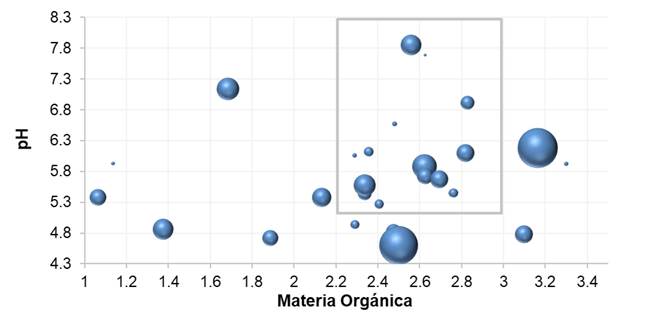
Figure 6. Relationship of pH and MO in function to Fusarium Index. The box shows a potential pH: MO threshold associated with low levels of IF. The size of the circle is proportional to the value of IF. Summer 2016. Los Altos, Jalisco.
Sequence analysis and phylogeny confirmed the intra and interspecific variability of Fusarium and the association of haplotypes, referred to here as intra-specific sequence variants based on the EF-1a gene, with specific symptoms. Clearly, wilting was associated with more haplotypes specific to this symptom belonging to F. solani, F. falciforme and F. oxysporum. The later was more diverse and included the only haplotype associated exclusively to dry bud rot and the only one found in apparently healthy plantations. The existence of only one haplotype by complex associated only with dry bud rot or with the ability to cause both types of symptoms suggests an interspecific process of divergent evolution with adaptation to aerial symptoms, wilt being the parasitic condition associated to the ancestral specie(s). The occurrence of two unidentified species may involve advanced speciation or less likely, a convergent evolution process. The possibility of these population genetics processes is highly possible considering the continued and confined agave cultivation for more than 200 years and the agave semi-perennial condition with vegetative propagation. These adaptive mechanisms have been suggested in crops such as mango (Mangifera indica) given the increasing variability and occurrence of new Fusarium species (Liew et al., 2016). In this context, it is concluded that the wilting and dry rot of the agave bud are symptomatic expressions of a syndrome caused by different Fusarium species with differential parasitic adaptability at the intra and interspecies level. One single isolate (haplotype) of F. oxysporum, F. solani and F. falciforme can apparently cause wilt and dry bud rot but are not the most prevalent, which explain the low incidence of both symptoms at the same plant in the field. In total, these species have the highest numbers of isolates (haplotypes) specialized only to cause wilting, thus clarify the intense occurrence and epidemic rates reported for this symptom (Rubio, 2007; Ramírez-Ramírez et al., 2017).
These findings require further specialized epidemiological and pathogenicity studies, expanding other DOT agave regions, air tissue sampling, including more sequences, and the use of other genes such as those associated with RPB1 and RPB2 (O’Donnell et al., 2015). However, these results contribute to the improvement of regional control and monitoring strategies by clarifying the etiology of the three types of bud rot (Gómez-Ortiz et al., 2011; Jiménez-Hidalgo et al., 2004; Rincón-Enríquez et al., 2014; Coria-Contreras et al., 2019). To this respect, in this work and that conducted with grey spot at state level (Coria-Contreras et al., 2019), the bacterial rot was never detected nor isolated from the field indicating a too low prevalence or occurrence only under specific agronomic and/or climatic condition (Jiménez-Hidalgo et al., 2004; Rincón-Enríquez et al., 2014).
Regional etiological studies, in addition to an epidemiological approach (Ceja-Torres et al., 2008), require methodological strategies to optimize time, economic resources and to provide a rational framework. This work shows the importance of combining morphological and classical cultural strategies with the current molecular approach (Leslie and Summerell, 2006; O’Donnell et al., 2015; Gogoi et al., 2017). The characterization of a total of 109 regional Fusarium isolates in five culture media allowed categorizing by groups and finally selecting the collection of representative strains for molecular identification (Burgess et al., 1991; Leslie and Summerell, 2006). Previously determined as important for screening Fusarium isolates in PDA (Gogoi et al., 2017), mycelial colorimetry was essential as an initial phase due to the large amount of initial cultures in Komada (248). On the other hand, due to the complexity of the etiological problem of this work, the ITS region was not functional for the identification of Fusarium.Vetrovsky and coworkers (2016) indicate that this region is not suitable due to its high variability, its multi-copy nature and its variable length. Additionally, ITS is a region highly conserved to resolve interspecific Fusarium boundaries. Therefore, it is estimated that 50% of Fusarium sequences or more are incorrectly identified at NCBI (O’Donnell et al., 2015). These aspects are fundamental from the etiological and population genetic perspective due to the current relevance of Fusarium. In Mexico, recent studies show the importance of this genus in crops such as Jatropha curcas, Capsicum pubescens and Citrus sinensis (Herrera-Parra et al., 2017; Almaraz-Sánchez et al., 2019; Parra-Cota et al., 2019).
Although the Fusarium Index IF was highly variable between counties and plantation phytosanitary condition, IF is proposed as an indicator to relate fungus richness to Fusarium spp. to estimate soil suppressive condition under the assumption that sustainable management of Fusarium, a fungus with high saprophytic-parasitic capacity, should be based on prevention and promoting soil health. IF variability can be explained by its multifactorial determinant as evidenced by its inverse relationship with pH and MO (r 2 = 0.72 - 0.68). The inductive effect of Fusarium populations due to acid pH has been reported in Fusarium oxysporum f. sp. cubense (Huang et al., 2019) and Fusarium oxysporum f. sp. fragariae. In this case, pH lower than 5.2 was more inductive (Fang et al., 2012). Similarly, a MO rich soil (i.e. greater than 3%) is widely accepted as a condition of suppressive soils that favors competition or inhibition of Fusarium spp. due to beneficial microorganisms (Orr and Nelson, 2018; Huang et al., 2019). In this investigation, Trichoderma spp. and Bacillus spp. frequently were found associated to agave plantations with low IF (CP-LANREF. Unpublished data) which demonstrates the need for regional comprehensive studies. The pH:MO thresholds suggestion in this work has potential to estimate productive risks due to Fusarium spp. in agave.
The diversity, complexity and prevalence of Fusarium species in commercial plantations at Los Altos and the historical productive impacts caused by this fungus stimulate to investigate the wilt and dry bud rot syndrome with a comprehensive approach relating the pathosystem to physical-chemical soil variables, antagonists richness, climatic factors and crop management. This approach will allow effective preventive management strategies and sustaining soil health. Likewise, it will help to estimate and mitigate risks associated with Fusarium spp. due to the intense mobilization of vegetative planting material required for crop expansion upon demand on tequila exports, currently in fifth income place among Mexican agricultural commodities. Blue agave constitutes a valuable crop chain that justifies research and innovation investment to assure competitiveness and the socio-cultural and environmental preservation values.
Conclusions
This research provides for the first time exhaustive etiological information associated with wilting and dry bud rot of blue agave (A. tequilana) in 40 commercial plantations of 11 counties at Los Altos Jalisco. Morphological, cultural and molecular characterization of Fusarium spp. isolates associated with these symptoms allows unifying them as expressions of a syndrome caused by five species of Fusarium belonging to F. oxysporum (FOSC), F. solani (FSSC) and F. fujikuroi (FFSC) complexes. It is proposed to name this disease as wilt and dry bud rot of agave. Wilt and dry bud rot can be caused by a single isolate (haplotype) of F. oxysporum, F. solani and F. falciforme but, contrary to those that cause only wilting, they were not the most prevalent. F. oxysporum was the most variable and prevalent in soil (46.2%) and root (66.7%) suggesting greater parasitic fitness. F. falciforme and two unidentified species belonging to FSSC and FFSC complexes are reported for the first time associated with agave. Although, these results require greater precision through epidemiological and pathogenicity studies, the extensive nature of this work confers clear etiological trends. The Fusarium Index IF has potential as indicator of fungal richness respect to Fusarium spp. to estimate soil suppressive condition. A significant inverse relationship of IF with pH and MO was found, allowing to suggest an integrated threshold to estimate productive risks due to Fusarium. Finally, considering the official campaign against agave pests, these results justify use of independent wilt and dry bud rot diagrammatic scales for monitoring purposes. However, the inclusion of algorithms in the SIVEA digital system is recommended to automatize analysis of the syndrome regional incidence for risk analysis purposes.
Acknowledgement
The authors thank to CNRF-DGSV for the financial support through the SENASICA-CRT agreement for the execution of the SIVEA project. To COLPOS and CONACYT for the first author’s master’s degree scholarship. To Casa Herradura, Casa Sauza and CESAVEJAL for logistical support and/or infrastructure. To CP-LANREF team for its unconditional support in the development of this research. To Miranda Mora Gutiérrez for the English translation.
REFERENCES
Acevedo-Sánchez, G., Santana-Peñaloza, B., López-Bautista, V., Martínez-Bustamante, V., Coria-Contreras, J., Mendoza-Ramos, C., Jiménez-González, L., y Mora-Aguilera, G. 2017. Criterios epidemiológicos para implementar una red de monitoreo de enfermedades causadas por Fusarium spp. en regiones agaveras de Jalisco. Revista Mexicana de Fitopatología 35: 176-177. http://rmf.smf.org.mx/suple mento/Suplemento352017.html [ Links ]
Almaraz-Sánchez, A., Ayala-Escobar, V., Tlatilpa-Santamaría, IF. y Nieto-Angel, D. 2018. Fusarium sambucinum Fuckel agente causal de la pudrición de frutos de chile manzano (Capsicum pubescens) en México. Revista Mexicana de Fitopatología 37(1): 159-169. http://dx.doi.org/10.18781/R.MEX.FIT.1810-2 [ Links ]
Ávila-Miranda, ME., López-Zazueta, JG., Arias-Castro, C., Rodríguez-Mendiola, MA., Guzmán-de Peña, DA., Vera-Núñez, JA. and Peña-Cabriales, J.J. 2010. Vascular wilt caused by Fusarium oxysporum in agave (Agave tequilana Weber var. azul). Journal of the Professional Association for Cactus Development 12: 166-180. https://pdfs.semanticscholar.org/b624/2327d0c870be2105ebda216325f241da3a7c.pdf [ Links ]
Burgess, LW., Summerell, BA. and Nelson, PE. 1991. An evaluation of several media for use in identification of some Fusarium spp. Australasian Plant Pathology 20: 86-88. https://doi.org/10.1071/APP9910086 [ Links ]
Ceja-Torres, LF., Mora-Aguilera, G., Téliz, D., Mora-Aguilera, A., Sánchez-García, P., Muñoz-Ruíz, C., Tlapal-Bolaños, B. y De La Torre-Almaraz, R. 2008. Ocurrencia de hongos y etiología de la secadera de la fresa con diferentes sistemas de manejo agronómico. Agrociencia 42(4): 451-461. http://www.scielo.org.mx/scielo.php?script=sci_arttext&pid=S1405-31952008000400008&lng=es&nrm=iso [ Links ]
Coria-Contreras, JJ., Mora-Aguilera, G., Yañez-Morales, MJ., Acevedo-Sánchez, G., Santana-Peñaloza, B., Mendoza-Ramos, C., Jiménez-González, L,, Martínez-Bustamante, VI., García-Martínez, DC. and Rubio-Cortés, R. 2019. Applied regional epidemiology to inductive characterization and forecasting of blue agave gray spot (Cercospora agavicola) in Jalisco, Mexico. Mexican Journal of Phytopathology 37(1): 71-94. http://dx.doi.org/10.18781/R.MEX.FIT.1809-4 [ Links ]
CRT, Consejo Regulador del Tequila. 2017. Geografía de la DOT. https://www.crt.org.mx/index.php/es/ (consulta, enero 2017). [ Links ]
Fang, X., You, MP. and Barbetti, MJ. 2012. Reduced severity and impact of Fusarium wilt on strawberry by manipulation of soil pH, soil organic amendments and crop rotation. European Journal of Plant Pathology 134: 619-629. https://doi.org/10.1007/s10658-012-0042-1 [ Links ]
Gogoi, M., Sarmah, DK. and Ali, S. 2017. Cultural and morphological variations of Fusarium solani (Mart.) Sacc. causing root rot of patchouli in Assam, India. International Journal of Current Microbiology and Applied Sciences 6(11): 1889-1901. https://doi.org/10.20546/ijcmas.2017.611.225 [ Links ]
Gómez-Ortiz, P., Sánchez-Arizpe, A., Virgen-Calleros, G., Carvajal-Cazola, CR. y Padrón-Corral, E. 2011. Incidencia y severidad de la marchitez del Agave tequilana Weber var. azul en la zona sur del estado de Nayarit, México. Agraria 8: 21-25. http://www.uaaan.mx/agraria/attachments/article/45/Agraria_2011(8)-2-3.pdf [ Links ]
Herrera-Parra, E., Cristóbal-Alejo, J., Martínez-Bolaños, M., Hernández-Arenas, M. y López-Guillén, G. 2017. Primer registro de Fusarium solani y F. equiseti en plantaciones de Jatropha curcas en México. Revista Mexicana de Fitopatología 35: 150-161. http://dx.doi.org/10.18781/r.mex.fit.1608-1 [ Links ]
Huang, J., Pang, Y., Zhang, F., Huang, Q., Zhang, M., Tang, S., Fu, H. and Li, P. 2019. Suppression of Fusarium wilt of banana by combining acid soil ameliorant with biofertilizer made from Bacillus velezensis H-6. European Journal of Plant Pathology 154: 585-596. https://doi.org/10.1007/s10658-019-01683-5 [ Links ]
Jiménez-González, LR., Mendoza-Ramos, C., Santana-Peñaloza, B., Coria-Contreras, JJ., Delgado-Mora, F., Acevedo-Sánchez, G., Guzmán-Hernández, E. y Mora-Aguilera, G. 2017. Esca la logarítmica diagramática de severidad para medición de pudrición seca del cogollo, marchitez y mancha gris del agave azul. Revista Mexicana de Fitopatología 35(S): 177. http://rmf.smf.org.mx/suplemento/Suplemento352017.html [ Links ]
Jiménez-Hidalgo, I., Virgen-Calleros, G., Martínez-de la Vega, O., Vandemark, G. and Olalde-Portugal, V. 2004. Identification and characterization of bacteria causing soft-rot inAgave tequilana. European Journal of Plant Pathology 110: 317-331. https://doi.org/10.1023/B:EJPP.0000019791.81935.6d [ Links ]
Leslie, JF. and Summerell, BA. 2006. The Fusarium laboratory manual. Ames, lowa: Blackwell Publishing. 388 pp. [ Links ]
Liew, ECY., Laurence, MH., Pearce, CA., Shivas, RG., Johnson, GI., Tan, YP., Edwards, J., Perry, S., Cooke, AW. and Summerell, BA. 2016. Review of Fusarium species in association with mango malformation in Australia. Australasian Plant Pathology 45:547-559. https://doi.org/10.1007/s13313-016-0454-z [ Links ]
O’Donnell, K., Ward, T., Robert, V., Crous, P., Geiser, D. and Kang, S. 2015. DNA sequence-based identification of Fusarium: Current status and future directions. Phytoparasitica 43: 583-595. https://doi.org/10.1007/s12600-015-0484-z [ Links ]
Orr, R. and Nelson, PN. 2018. Impacts of soil abiotic attributes on Fusarium wilt, focusing on bananas. Applied Soil Ecology 132: 20-33. https://doi.org/10.1016/j.apsoil.2018.06.019 [ Links ]
Parra-Cota, FI., García-Pereyra, J., Aviña-Martínez, GN. y de los Santos-Villalobos, S. 2018. Primer reporte de marchitamiento por Fusarium en Citrus sinensis var. Valencia en el Valle del Yaqui, México. Revista Mexicana de Fitopatología 37(1): 193-201. http://dx.doi.org/10.18781/R.MEX.FIT.1810-3 [ Links ]
Ramírez-Ramírez, MJ., Mancilla-Margalli, NA., Meza-Álvarez, L., Turincio-Tadeo, R., Guzmán-de Peña, D. and Ávila-Miranda, ME. 2017. Epidemiology of Fusarium agave wilt in Agave tequilana Weber var. azul. Plant Protection Science 53: 144-152. https://doi.org/10.17221/142/2016-PPS [ Links ]
Rincón-Enríquez, G., Quiñones-Aguilar, E., Qui-Zapata, J., Vega-Ramos, K. y Uvalle-Bueno, J. 2014. Selección de bacteriófagos para el control biológico del agente causal de la pudrición del Agave tequilana. Revista Mexicana de Fitopatología 32(S): 112. http://rmf.smf.org.mx/suplemento/Suplemento322014.html [ Links ]
Rubio, CR. 2007. Enfermedades del cultivo de agave. Pp:169-195. In: Domínguez, PJF (ed.). Conocimiento y prácticas agronómicas para la producción deAgave tequilanaWeber en la zona con denominación de origen del tequila. Libro técnico No. 4. INIFAP-CIRPAC. Tepatitlán de Morelos, Jalisco, México. 196p. http://biblioteca.inifap.gob.mx:8080/jspui/handle/123456789/630 [ Links ]
SIAP. 2018. Servicio de Información Agrícola y Pes quera. SAGARPA. https://www.gob.mx/siap (Consultado enero 2018). [ Links ]
Vega-Ramos, KL., Uvalle-Bueno, JX. and Gómez-Leyva, JF. 2013. Molecular variability among isolates of Fusarium oxysporum associated with root rot disease of Agave tequilana. Biochemical Genetics 51: 243-255. https://doi.org/10.1007/s10528-012-9559-4 [ Links ]
Vetrovsky, T., Kolarik, M., Zifcakova, L., Zelenka, T. and Baldrian, P. 2016. The rpb2 gene represents a viable alternative molecular marker for the analysis of environmental fungal communities. Molecular Ecology Resources 16: 388-401. https://doi.org/10.1111/1755-0998.12456 [ Links ]
White, TJ., Bruns, T., Lee, S. and Taylor, J. 1990. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In: Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J. (Eds.), PCR Protocols: A Guide to Methods and Applications. Academic Press, San Diego. 315-322. https://nature.berkeley.edu/brunslab/papers/white1990.pdf [ Links ]
Received: November 14, 2019; Accepted: December 20, 2019
 Este es un artículo publicado en acceso abierto bajo una licencia
Creative Commons
Este es un artículo publicado en acceso abierto bajo una licencia
Creative Commons
