











 text new page (beta)
text new page (beta) Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
Permalink
Introducción
Las Rhamnaceae fueron descritas por Antonio Laurent de Jussieu en 1789 y Brongniart en 1827 subdivide a la familia en cinco tribus donde Colubrina es incorporada en la Rhamnaceae (Fernandez, 1993 p.8). Johnston (1963), publica nuevas combinaciones, nuevos taxones y dos subgéneros denominados Colubrina y Serrataria, en el primero se incluye a Colubrina gregii S. Wats. var. yucatanensis M. C. Johnst. y en el segundo a Colubrina texensis (T. & G.) A. Gray var. pedunculata M. C. Johnst. y Colubrina texensis (Cav.) G. Don var. lanulosa (Blake) M. C. Johnston. Posteriormente, Johnston (1971) publica la revisión de Colubrina, y propone la subdivisión de subgéneros y secciones de esta entidad taxonómica.
Para la circunscripción de Colubrina (Johnston, 1971) refiere los siguientes caracteres: árboles y arbustos, raramente trepadores, espinosos o sin espinas, caducifolios o siempre verdes, tallos generalmente pubescentes; hojas alternas u opuestas, pecioladas, penninervadas o triplinervadas desde la base, frecuentemente con pequeñas manchas glandulares redondeadas, ya sea en el margen o cerca de la base o raramente con pocas glándulas dispersas, margen entero, serrados o crenado; estípulas laterales y basales generalmente caducas; inflorescencias cimosas sésiles y umbeliformes o pedunculadas corimbiformes, con el eje ocasionalmente modificado en una espina; flores pequeñas, verdosas o amarillentas, con copa floral donde están insertos el cáliz, la corola y el androceo, dicha copa después de la polinización se acrescenta y permanece sujeta a la parte inferior a la mitad del fruto; sépalos 5 caducifolios, pétalos 5 más cortos que los sépalos, estambres 5, casi tan largos como los pétalos, nectarios, casi llenando la copa y en las primeras etapas ocultan al ovario que lleva tres óvulos (raramente cuatro), estigmas tres; fruto en forma de cápsula subglobosa, dehiscente en la madurez, semillas por lo general ventralmente angulosas y dorsalmente fuertemente convexas, de color marrón oscuro o negro (Johnston, 1971, p. 7-8); este mismo autor consideró a los géneros Cormonema e Hybosperma como afines a Colubrina¸ por lo cual se consideraron dentro de este último género (Johnston, 1971, p. 7).
Los taxones que se estudiaron en este trabajo corresponde al subgénero Serrataria (Johnston, 1971. p.8) en donde se conjuntan aquellas especies que presentan los márgenes de las láminas foliares aserrados, con más de diez dientes en cada lado y cada uno asociado con una glándula marginal con distribución en América y Asia, incluye los siguientes taxones (Cuadro 1).
Cuadro 1 Especies de Colubrina subgénero Serrataria, distribución mundial y cambios nomenclaturales.Table 1. Species of Colubrina subgenus Serrataria, worldwide distribution, and nomenclatural changes.
| Nombre de la especie y cambios nomenclaturales | Distribución en América |
|---|---|
| C. berteroana Urb. | República Dominicana |
| C. cubensis (Jacq.) Brongn. | Florida y Cuba |
| C. californica I. M. Johnst. | Baja California, California y Arizona |
| C. celtidifolia (Cham. & Schltdl.) Schltdl. | Veracruz, Jalisco, Michoacán y Guerrero |
| Según Johnston (1971)C. greggii S. Wats. var. angustior M. C. Johnston Según Nesom (2013)C. angustior (M. C. Johnston) Nesom | San Luis Potosí, Tamaulipas y Veracruz |
| Según Johnston (1971)C. greggii S. Wats. var. greggii Según Nesom (2013)C. greggii S. Wats. | Texas, Coahuila, Nuevo León, Tamaulipas, Guanajuato, Hidalgo, Puebla, Oaxaca, Querétaro, San Luis Potosí y Veracruz |
| C. greggii S. Wats. var. yucatensis M. C. Johnston Según Nesom (2013)C. yucatanensis (M. C. Johnston) Nesom | Campeche, Quintana Roo, Yucatán y Guatemala |
| C. macrocarpa (Cav.) G. Don var. lanulosa (S. F. Blake) M. C. Johnston | Guerrero |
| C. macrocarpa (Cav.) G. Don var. macrocarpa | Morelos y Puebla |
| C. macrocarpa (Cav.) G. Don var. macrocarpoides (Suessenguth ex Suessenguth & Overkott) M. C. Johnston | Querétaro |
| C. sordida M. C. Johnston | Guerrero |
| C. stricta Engelm. ex Blankinship | Texas, Nuevo León, Coahuila, Nesom (2013) la cita también para Durango |
| Según Johnston (1971)Colubrina texensis (Torrey & Gray) A. Gray var. pedunculata M. C. Johnston Según Nesom (2013)C. stricta Engelm. ex Blankinship | Texas y Coahuila |
| Según Johnston (1971)C. texensis (Torrey & Gray) A. Gray var. texensis Según Nesom (2013)C. texensis (Torrey & Gray) A. Gray | Texas, noreste de Coahuila, Nuevo León y Tamaulipas Texas, Coahuila y Nuevo León |
| Distribución en Asia, África y Australia | |
| C. anomala King | Malasia |
| C. asiatica (L.) Brongn. | África (Mozambique y Kenia), India, Nueva Guinea y Caribe |
| C. beccariana Warb. | Nueva Guinea y Borneo |
| C. pedunculata Baker | India |
| C. travancorica Bedd. | India |
Nesom (2013), en las notas del género Colubrina considera que las variedades de C. greggii y C. texensis deben tratarse como especies y no como variedades como las describe Johnston (1971). Nesom (2013) sustenta estos cambios con base en datos morfológicos y de distribución geográfica. De esta manera Colubrina texensis var. texensis queda como C. texensis (Torrey & Gray) A. Gray con distribución en Texas, noreste de Coahuila, Nuevo León y Tamaulipas, mientras que Colubrina texensis var. pedunculata se valida como Colubrina stricta Engelm. ex Blankinship quedando como sinónimo C. texensis (Torrey & A. Gray) A. Gray var. pedunculata M. C. Johnston.
En este mismo contexto, Nesom (2013), eleva a rango de especies las variedades de Colubrina greggii (sensuJohnston, 1971) y así separa a Colubrina greggii S. Wats., Colubrina angustior (M. C. Johnston) Nesom y Colubrina yucatensis (M. C. Johnston) Nesom por la forma de las hojas y caracteres florales y por la distribución geográfica. La primera especie presente en Texas, Coahuila, Nuevo León, Tamaulipas, Guanajuato, Hidalgo, Puebla, Oaxaca, Querétaro, San Luis Potosí y Veracruz; la segunda en San Luis Potosí, Tamaulipas y Veracruz y la tercera en Campeche, Quintana Roo, Yucatán, y Guatemala.
La familia Rhamnaceae ha sido estudiada desde el punto de vista taxonómico-florístico por autores como: Standley (1923), Johnston (1963, 1969 y 1971), Wiggins (1964 y 1980) Martínez (1979), Fernández (1993, 1996, 2010) y Nesom (2013). Entre los trabajos relacionados con la anatomía y arquitectura foliar de la familia Rhamnaceae encontramos los de Meyer y Meola (1978) que citan el tamaño de los estomas y la densidad por mm2 de Colubrina texensis; Schirarend (1991) investiga la anatomía del tallo de la Tribu Zizipheae; Colares y Arambarri (2008) describe la anatomía de Ziziphus mistol Griseb.; Shisode y Patil, (2011) estudiaron la anatomía del tallo de algunas Rhamnaceae; Hernández et al. (2021) describen la arquitectura foliar y la anatomía epidérmica de las especies mexicanas del género Gouania. El interés de este trabajo fue describir la arquitectura foliar y las superficies epidérmicas de las variedades de Colubrina greggii, C. texensis y C. macrocarpa sensuJohnston (1971) que prosperan en México, así como apoyar o denegar algunos de los cambios nomenclaturales propuestos en los últimos años.
Materiales y métodos
El material utilizado se encuentra depositado en el Herbario de la Escuela Nacional de Ciencias Biológicas del Instituto Politécnico Nacional (ENCB) y del Herbario del Instituto de Biología de la Universidad Nacional Autónoma de México (MEXU). La identificación de los taxones fue confirmada por el Dr. Rafael Fernández Nava especialista en Rhamnaceae. Los ejemplares de herbario de los que se tomaron muestras fueron los siguientes:
Colubrina angustior (M. C. Johnston) Nesom; J. Rzedowski 10321 (ENCB); J. Rzewdoski 7032 (ENCB); P. A. Fryxell y W.R. Anderson 3428 (ENCB).
Colubrina greggii S. Wats.; R. Fernández 1650 (ENCB); H. Puig 7255 (ENCB); R. Fernández 2406 (ENCB).
Colubrina yucatensis (M. C. Johnston) Nesom; L. M. Arias 649 (ENCB); E. Estrada E-110, (ENCB); M. García y A. Vargas 3777 (ENCB).
C. macrocarpa (Cav.) G. Don var. macrocarpa; R. Fernández. 2621 (ENCB); R. Fernández 4984 (ENCB); R. Weber 307 (ENCB).
C. lanulosa (M. C. Johnston) Fernández & Arreguín; Blanco, Toledo y Cabrera 765 (ENCB); E. Halbinger s/n (ENCB), J. Chavelas GR 22 (ENCB).
C. stricta Engelm. ex Blankinship; J. García INEGI 3.2V (ENCB).
C. texensis (Torrey & Gray) A. Gray; A. Rodríguez y M. A. Carranza s/n (ENCB), M. Martínez 44266 (ENCB), E. Torrecillas 176 (ENCB).
Se utilizaron las siguientes técnicas para transparentar las láminas foliares, sugeridas por Aguirre Claverán y Arreguín Sánchez (1988) y el método químico de decoloración foliar de Payne, Sheffy y Potter (1969) modificado por Dilcher (1974).
De cada espécimen se tomaron cuatro hojas maduras totalmente extendidas situadas entre el tercero y quinto nudo de las ramas montadas en las muestras de herbario y de preferencia de tres ejemplares de cada especie y variedad, aunque de C. stricta solo fue posible obtener muestras de un solo espécimen debido a que hay pocos ejemplares representados en los herbarios o bien los ejemplares estaban adheridos a la cartulina a través de pegamento blanco y no se pudieron obtener muestras. En las láminas de las fotografías de cada especie incluye la hoja antes de diafanizar.
Una vez que se obtuvo el material foliar, se prosiguió con la técnica de Aguirre-Claverán y Arreguín-Sánchez (1988) para lo cual se colocaron dos hojas completas de cada uno de los taxones los cuales se diafanizadas con NaOH al 5% hasta cubrir el material; se hirvieron de 5 a 10 minutos, dependiendo de la textura de la hoja, luego se pasaron a una caja de Petri con agua por 10 a 15 minutos, posteriormente se colocaron en una solución de hipoclorito de sodio al 30% y se dejaron hervir hasta que los tejidos se transparentaron por completo, se colocaron en agua hasta el día siguiente. Las láminas aclaradas siempre permanecieron en cajas de Petri con agua y se tomaron fotografías de este material.
Posteriormente una de las hojas contenidas en la caja de Petri, se tiñó con dos gotas de azul de metileno comercial “fish care” por un minuto y después se pasó a otra caja con agua. A la segunda hoja se les añadió dos gotas de safranina por dos minutos y se colocaron en cajas de Petri con agua. Se tomaron fotografías de este material (Hernández et al., 2021).
Con la técnica de Payne, Sheffy y Potter (1969) tomada de Dilcher (1974, p. 60) dos hojas del material seco se colocaron sobre una caja Petri adicionando NaOH al 5%, en seguida se calentó por 5 minutos y después se realizó un lavado con agua de la llave, posteriormente se blanqueó con hipoclorito de sodio al 5% por 10-15 minutos, checando el material frecuentemente hasta que se transparentó y se lavó con agua dos veces. Después el material se deshidrató en series de alcoholes al 50% y 75% durante de 10 a 15 minutos en cada solución en alcohol al 95% durante 20 minutos y finalmente en alcohol al 100% para alcanzar la completa deshidratación de las muestras, después se transfirieron a una solución de alcohol absoluto y xileno 1:1, posteriormente se transfiere a xilol al 100%. Para la tinción se pasaron a una caja de Petri con agua y se siguió el procedimiento referido por Hernández et al. (2021). Se tomaron fotografías con una cámara digital Samsung Hmx-e10 con 8 megapixeles.
El propósito de utilizar azul de metileno y safranina acuosa como colorantes permitió visualizar mejor diferentes características anatómicas y contrastar algunas de ellas. El utilizar dos técnicas de transparentación permitió que con la técnica de Aguirre-Claverán y Arreguín-Sánchez (1988) permitió mejores observaciones para hojas membranáceas y subcoriaceas y la de Payne, Sheffy y Potter (1969) para material coriáceo.
La arquitectura foliar se describió bajo un microscopio estereoscópico American Optical utilizando diferentes aumentos para describir las venas de primera, segunda, tercera y cuarta categoría, posición de los tricomas y la posible presencia de estructuras glandulares. Las fotografías se tomaron con una cámara digital Samsung Hmx-e10 con 8 megapixeles.
En cuanto a la superficie epidérmica se hicieron cortes de 1 cm de porciones de las hojas teñidas con azul de metileno y safranina que se colocaron en diferentes portaobjetos y como medio de montaje se utilizó una solución de miel de maíz con agua 1:1 y trazas de fenol, después se cubrieron con los cubreobjetos y se sellaron con barniz transparente, una vez seco el esmalte las preparaciones se etiquetaron. Las descripciones de los caracteres de la superficie epidérmica se realizaron con un microscopio óptico binocular Zeiss y las fotomicrografías se obtuvieron con una cámara digital Samsung Hmx-e10 con 8 megapixeles adaptada al microscopio.
Con base en las preparaciones semipermanentes que se elaboraron se midieron cinco tricomas de cada especie y espécimen en relación al largo de los mismos, las medidas que se presentan es el promedio de ellos, así también se contó el número de tricomas por mm2. Para los estomas se midieron cinco de cada especie y espécimen y se tomó en consideración la longitud y el ancho, las medidas que se citan es el promedio de estas mediciones. La densidad estomática se calculó según la fórmula de (Salisbury, 1927 mencionado en Colares y Arambarri (2008, p. 570) . Número de estomas + número de células epidérmicas x 100.
Los términos que se utilizaron para describir los tricomas fueron los de Dilcher (1974, p. 110-111) , este autor realizó una clasificación de los mismos en relación a varios criterios como: número de células que los forman; si los tricomas son simples o agrupados; glandulares o eglandulares; la forma por la que se unen a las células epidérmicas con un pie apenas modificado, en forma de pija, a través de dos células o de varias; con base engrosada o no, con células epidérmicas no modificadas o modificadas.
En el caso de este trabajo todos los taxones presentaron tricomas unicelulares, simples, eglandulares y con relación a la base se encontraron dos tipos: uno en Colubrina greggii y C. angustior con base engrosada y células epidérmicas no modificadas (Figura 1A). El dibujo corresponde al trabajo de Dilcher (1974, p.110) y las fotomicrografías a las bases de las especies antes citadas. El otro tipo se muestra en la (Figura 1B) presentes en el resto de las especies, donde la base está engrosada y las células epidérmicas modificadas y a esto le nombra radial (R). El dibujo corresponde a Dilcher (1974, p. 111) y la fotomicrografía corresponde a Colubrina macrocarpa. En las descripciones de este trabajo, el primer tipo se describió como tricomas con base engrosada y células epidérmicas no modificadas (Figura 1A) y al segundo como base radial (Figura 1B).
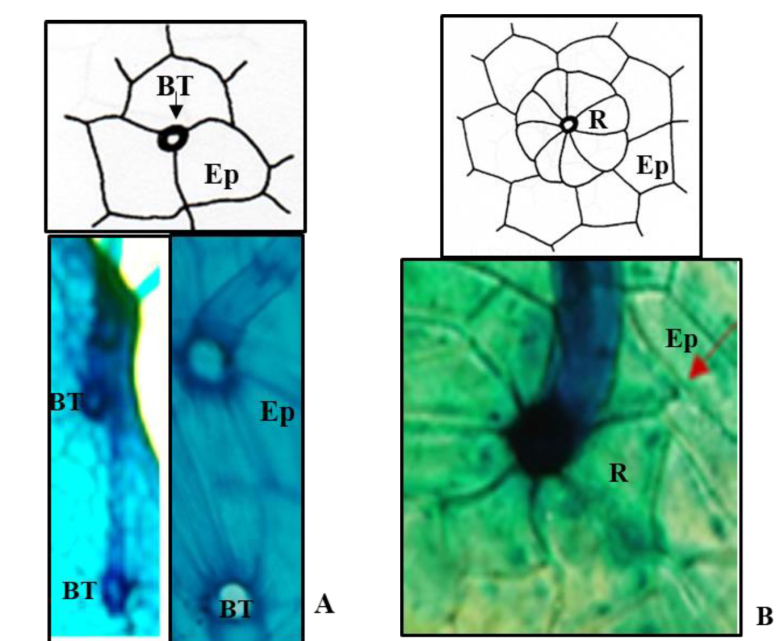
Figura 1 Base de los tricomas. A, tricoma con base engrosada y células epidérmicas no modificadas. B, tricoma con base radial de células epidérmicas modificadas. Referencias: BT inserción de la base de los tricomas. Ep células epidérmicas no modificadas. R Células epidérmicas modificadas en forma radial.
Para las descripciones de la arquitectura foliar se tomaron en consideración los trabajos de (Hickey, 1973 y 1974), Dilcher (1974), Hickey y Wolfe (1975), Gomes et al. (2011) y fundamentalmente se siguieron los términos y criterio de Ash, et al. (1999). Para la epidermis foliar se consideraron los criterios de Gola et al., (1965); Dilcher (1974), Cortés (1982) y Azcárraga et al., (2010). Los caracteres que se tomaron en consideración son los siguientes:
Caracteres exomorfológico y de arquitectura foliar: tipos de hojas (simples o compuestas), simetría, forma, medidas de largo y ancho, ápice, base, textura, margen, presencia o ausencia de glándulas, peciolo, tipo de arquitectura foliar, venas de primera, segunda, tercera y cuarta categoría.
Caracteres de epidermis foliar: tipos de tricomas, tamaño, base de los mismos y densidad por mm2; forma de células epidérmicas adaxial y abaxial y tipos de paredes; tipos de estomas, longitud, ancho y densidad estomática.
Resultados
A continuación, se describe e ilustra la arquitectura y superficie foliar de las especies de las especies de Colubrina estudiadas.
1. -Colubrina greggii S. Wats. (Figura 2)
Láminas foliares simples, simétricas, ovadas a lanceoladas-ovadas o elíptico-ovadas de 4 a 18 cm de largo y 3 a 10 cm de ancho, ápice claramente acuminado, agudo acuminado, acuminado corto a agudo, base redondeada a truncada a cordada, textura membranosa a semicoriácea, margen serrado con 60 a 130 dientes de cada lado (Figura 2A, B, C, D), sin glándulas (Figura 2E). Peciolo de 4 a 20 mm de largo y 1 a 2 mm de diámetro. Venación acródroma basal imperfecta donde se observan tres venas primarias basales que se originan en un punto de la hoja, la central sigue un curso recto, las dos externas ligeramente curveadas (Figura 2C, D), par inferior de las venas primarias laterales se ramifican (Figura 2C, D), y estas forman arcos en el margen de la lámina foliar y de ahí deriva una vena corta que alimenta a los pequeños dientes marginales (Figura 2E), venas secundarias craspedódromas, ligeramente curveadas y en el margen forman arcos (Figura 2C, D) venas terciarias reticuladas superadyacentes (Lám. 1F), las de cuarta categoría reticuladas (Figura 2F).
Epidermis en vista superficial. Células epidérmicas hexagonales con paredes rectas para ambas superficies (Figura 2G, H). Estomas de tipo anomocíticos en la superficie abaxial de 26.1 µm de longitud y 17.3 µm de ancho (Figura 2 ), densidad estomática 39/ mm2. Tricomas eglandulares en las superficies adaxial y abaxial, unicelulares de 32.3 µm, de largo, base engrosada y células epidérmicas no modificada, presentes en ambas superficies con una densidad de 12/mm2 (Figura 2G, H, I).

Figura 2 Colubrina greggii. A.- Lámina foliar de un ejemplar de herbario. B.- Detalle del margen de la lámina foliar, el círculo muestra, la zona donde se ampliaron los pequeños dientes. C y D.- Hojas diafanizada de diferentes formas mostrando las tres venas primarias basales ramificadas. E.- Venas de primera, segunda, tercera y cuarta categoría, venas secundarias formando arcos en el margen de donde deriva una pequeña vena que alimenta a los diminutos dientes (VRD). F.- Venas secundarias, terciarias y cuaternarias reticuladas. G.- Superficie adaxial mostrando un diente, tricomas con base engrosada y células epidérmicas hexagonales no modificadas. H.- Detalle superficie abaxial con tricomas y células epidérmicas hexagonales. I.- Detalles de las bases de los tricomas engrosadas. J.- Estomas anomocíticos. Referencias: BA bases de las láminas foliares donde parten las tres nervaduras de primera categoría. R ramificación del primer par basal de las venas primarias. P vena de primera categoría. S vena de segunda categoría. T vena de tercera categoría. C vena de cuarta categoría. RS ramificación de las venas secundarias que en el margen forma arcos. VRD Vena corta de la ramificación de la vena secundaria que alimenta a un diente marginal. Tr tricoma. BT base del tricoma. E Estoma. El círculo en D corresponde a la porción de la lámina foliar que se muestra a mayor aumento en la fotomicrografía de la letra E donde también se muestra un mayor acercamiento en la alimentación de la vena que alimenta a los dientes.
2. - Colubrina angustior (M. C. Johnston) Nesom (Figura 3)
Colubrina greggii S. Wats. var. angustior M. C. Johnston
Láminas foliares simples, simétricas (Figura 3B), ocasionalmente asimétricas en la base (Figura 3A); oblongo lanceoladas de 3 a 10 cm de largo y 2 a 4 cm de ancho, ápice agudo a levemente acuminado, base redondeada, textura membranosa, margen dentado con 50 a 70 pequeños dientes de cada lado (Figura 3D), sin glándulas (Figura 3D). Peciolo de 1 a 2 cm de largo y 1 a 2 mm de diámetro. Vena de primera categoría pinnada de curso recto (Figura 3A, B, C) las secundarias craspedódromas libres con 6 a 7 venas de cada lado, primer par basal ramificado, las terciarias reticuladas superadyacentes, en el margen estas venas alimentan los dientes que pueden o no formar arcos (Figura 3C, D), las de cuarta categoría reticuladas (Figura 3D).
Epidermis en vista superficial. Células de la epidermis adaxial hexagonales con paredes rectas (Figura 3E), las abaxiales con paredes onduladas (Figura 3G). Estomas en la superficie adaxial ausentes, en la abaxial anomocíticos de 26.1 µm de longitud y 17.3 µm de ancho, densidad estomática en promedio de cinco ejemplares 37/mm2 (Figura 3G, H). Tricomas eglandulares, con la base engrosada y células epidérmicas no modificada, se presentan en ambas superficies de 34.32 µm de largo con una densidad de 18/mm2 (Figura 3F, H).

Figura 3 Colubrina angustior. A y B.- Láminas foliares de diferentes formas tomadas de ejemplares de herbario. C.- Detalle de las venas de primera, segunda y tercera categoría. D.- Detalles de las venas marginales de segunda categoría, las de tercera formando o no arcos que alimentan a los pequeños dientes y venas de cuarta categoría reticulada. E.- Células de la epidermis adaxial hexagonales. F.- Detalle de la base de los tricomas engrosada y las células epidérmicas no modificadas. G.- Células de la epidermis abaxial con paredes onduladas. H.- Epidermis abaxial con tricomas y estomas. Referencias: P vena de primera categoría. S venas de segunda categoría. T venas de tercera categoría. C venas de cuarta categoría. Tr tricoma. BT base del tricoma. Di diente del margen. CAd Células de la superficie adaxial. CAb Células de la superficie abaxial. E Estoma.
3. - Colubrina yucatensis (M. C. Johnston) Nesom (Figura 4).
Colubrina greggii S. Wats. var. yucatanensis M. C. Johnston
Hojas simples, simétricas, oblongo-lanceoladas de 6.5 a 12 cm de largo y 3 a 7 cm de ancho, ápice agudo, acuminado a corto acuminado, base redondeada a truncada, textura membranácea (Figura 4A, B, C) margen serrulado con más de 80 dientes de cada lado (Figura 4B), sin glándulas (Figura 4D). Peciolo 4 a 20 mm de largo y 1 a 1.5 mm de diámetro. Vena primaria pinnada de curso recto (Figura 4A, C), las secundarias craspedródroma con 6 a 7 venas del lado izquierdo y derecho de la lámina foliar, primer par basal ramificado (Figura 4C), las de tercera categoría reticuladas superadyacentes, en el margen, estas venas alimentan a los pequeños dientes, a veces forman arcos (Figura 4D), las de cuarta categoría reticuladas (Figura 4D).
Epidermis en vista superficial: Células de la epidermis adaxial poligonales con paredes rectas y algunas más gruesas (Figura 4E), en la abaxial las células poseen paredes onduladas (Figura 4G). Estomas ausentes en la superficie adaxial, en la abaxial de tipo anomocíticos de 21.1 µm de longitud y 27 µm de ancho, densidad estomática en promedio de cinco ejemplares de 45/mm2 (Lám. 4G). Tricomas eglandulares unicelulares, con base radial de 5 células pentagonales presentes en ambas superficies de 27.2 µm de largo, con densidad de 27/mm2 (Figura 4F).
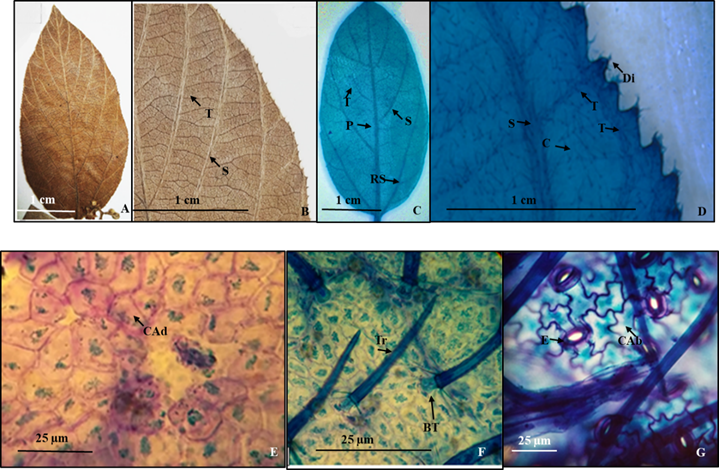
Figura 4 Colubrina yucatensis. A.- Lámina foliar de un ejemplar de herbario. B.- Detalle de la venación de la lámina foliar del ejemplar de herbario y del margen con pequeños dientes. C.- Lámina foliar diafanizada mostrando venas de primera, segunda y tercera categoría. D.- Detalle del margen con venas de tercera categoría alimentando un pequeño diente y venas de cuarta categoría reticuladas. E.- Células epidérmicas adaxiales poligonales con paredes rectas y algunas con paredes más gruesas. F.- Detalle de los tricomas con base radial (nótese la base engrosada y las células epidérmicas modificadas). G.- Células epidérmicas con paredes onduladas, estomas anomocíticos y tricomas. Referencias: P vena de primera categoría. S vena de segunda categoría. T vena de tercera categoría. C vena de cuarta categoría. Tr tricoma. BT base del tricoma. Di diente. CAd Células de la superficie adaxial. CAb Células de la superficie abaxial. E Estoma.
4.- Colubrina macrocarpa (Cav.) G. Don var. macrocarpa (Figura 5).
Hojas simples, simétricas, ovadas a ovado-oblongas de 2 a 9 cm de largo y 1 a 7 cm de ancho, ápice redondeado a veces agudo, base redondeada a truncada a levemente cordada, textura subcoriácea, margen serrado, con 50 a 70 dientes de cada lado (Figura 5A, B) sin glándulas (Figura 5C). Peciolo de 2 a 20 mm de largo y 1 a 2 mm de diámetro. Venación acródroma basal imperfecta, en su curso las tres venas parten de un mismo punto que convergen hacia el ápice (Figura 5B), venas de segunda categoría craspedódromas más o menos rectas, no ramificadas a veces 1-ramificadas que alimentan a los pequeños dientes (Figura 5D), las venas terciarias reticuladas superadyacentes, en el margen de la lámina también alimentan a los pequeños dientes (Figura 5D) con venas libres no ramificadas curveadas (Figura 5D, E).
Epidermis en vista superficial: Células de la epidermis adaxial alargadas con paredes rectas (Figura 5F), en la epidermis abaxial las células son pentagonales con paredes rectas (Figura 5G). Estomas en la abaxial mayoritariamente del tipo anisocíticos, ocasionalmente del tipo pericítico de 22.7 a 23.9 µm de longitud y ancho respectivamente, densidad estomática 21/ mm2 (Figura 5G). Tricomas unicelulares, eglandulares, base radial con 7 células epidérmicas pentagonales, dispuestos en ambas superficies de 25.32 µm de largo con una densidad de 38/mm2 (Lám. 5F).
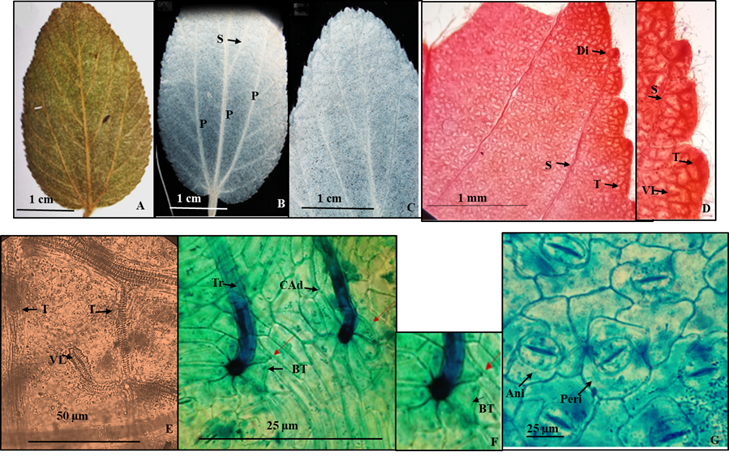
Figura 5 Colubrina macrocarpa var. macrocarpa. A.- Lámina foliar de material herborizado. B.- Material diafanizado, detalle de la base. C.- Material diafanizado, detalle del ápice y margen. D.- Detalle del margen donde se aprecia la alimentación de venas de segunda y tercera categoria que alimentan a los pequeños dientes. E.- Detalle de una vena de tercera categoría y en el interior una vena libre no ramificada curveada. F.- Detalle de la base radial de los tricomas y células epidérmicas adaxiales alargadas. G. Células epidérmicas abaxiales anticlinales con estomas anisocítos y pericíticos. Referencias: P venas de primera categoría. S vena de segunda categoría. T vena de tercera categoría. VL vena libre. Tr tricoma. BT base del tricoma. CAd Células de la superficie adaxial. E Ani estomas anisocíticos. Peri estomas pericíticos.
5.- Colubrina lanulosa (M. C. Johnston) Fernández & Arreguín, comb. et stat. nov. (Figura 6).
Colubrina macrocarpa (Cav.) G. Don var. lanulosa (S. F. Blake) M. C. Johnston.
Hojas simples, simétricas, oblonga a oblonga-lanceolada de 3 a 9 cm de largo y 2 a 7 cm de ancho, ápice agudo, base redondeada, textura subcoriácea, margen serrado con más de 50 dientes de cada lado o margen crenado (Figura 6A, B), sin glándulas (Figura 6C). Peciolo de 1.2 a 2 mm de largo y 1 a 1.5 mm de diámetro. Vena de primera categoria pinnada de curso recto (Figura 6A, B), las secundarias craspedródromas, cinco de cada lado, curveadas, el primer par inferior ramificado (Figura 6B) las venas secundarias en el margen 1-ramificadas y alimentan a los pequeños dientes y crenaciones (Figura 6D), las de tercera categoría reticuladas superadyacentes, las marginales alimentan los dientes (Figura 6E), las venas de cuarta reticuladas (Figura 6F).
Epidermis en vista superficial: Células en ambas superficies de la lámina foliar anticlinales con paredes rectas (Figura 6G). Estomas abaxiales mayoritariamente de tipo anisocíticos y algunos anomotetracíticos de 27.1 µm de longitud y 24 µm de ancho, densidad estomática 29/mm2 (Figura 6G, H). Tricomas eglandulares, unicelulares muy densos en ambas superficies de la lámina foliar insertos en las venas de primera, segunda, tercera y cuarta categoría, base radial con 6 células pentagonales dispuestos en ambas superficies de 25.32 µm con una densidad de 102/ mm2 (Figura 6G).
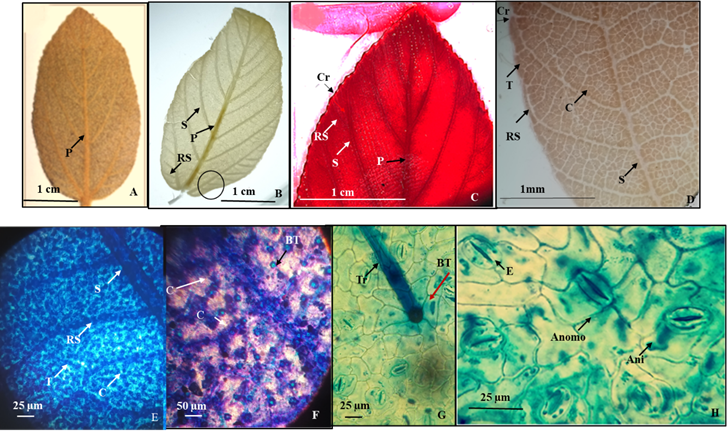
Figura 6 Colubrina lanulosa (M. C. Johnston) Fernández & Arreguín, comb. et stat. nov. A.- Lámina foliar de un ejemplar de herbario. B.- Lámina foliar diafanizada mostrando detalle de las venas secundarias inferiores ramificadas. C.- venas secundarias del margen que alimentan a los pequeños dientes. D.- Material diafanizado mostrando detalle de las venas de segunda y tercera categoría que en el margen alimentan a las crenaciones. E. Venas de segunda, tercera y cuarta categoría reticuladas. F.- Detalle de venas de cuarta categoría, los puntos brillantes son la base de los tricomas. G. Detalle de un tricoma con base radial. H. Células epidérmicas abaxiales mostrando estomas anisocíticos y anomotetracíticos. Referencias: P vena de primera categoría. S vena de segunda categoría. T vena de tercera categoría. C vena de cuarta categoría. RS Ramificación de vena secundaria. Cr crenación. Tr tricoma. BT base del tricoma. Anomo estoma anomotetracíticos. Ani estoma anisocítico. El círculo en B corresponde a la porción de la lámina foliar que se muestra a mayor aumento en las fotomicrografías de la letra D y E.
6.- Colubrina stricta Engelm. ex Blankinship (Figura 7).
Colubrina texensis (Torrey & Gray) A. Gray var. pedunculata M.C. Johnston
Hojas simples, simétricas, ovadas a ovado-oblongas de 3 a 7.5 cm de largo y 2 a 4 cm de ancho, ápice redondeado a agudo, base redondeada a truncada, textura subcoriácea, margen diminutamente serrado con 15 a 30 dientes a cada lado, muchas veces casi imperceptibles (Figura 7A, B), sin glándulas (Figura 7C). Peciolo de 1 a 2 cm de largo por 1 a 1.5 mm de diámetro. Vena de primera categoría pinnada con curso recto (Figura 7B), las secundarias caspedódromas, con cinco a seis a cada lado de la lámina foliar, no ramificadas (Figura 7B), las de tercera categoría reticuladas supradyacentes y en el margen alimentan a los diminutos dientes (Figura 7C, D), venas de cuarta categoría reticuladas (Figura 7D, E) que incluyen una vena no ramificada curveada (Figura 7E).
Epidermis en vista superficial: Células epidérmicas adaxiales y abaxiales hexagonales con paredes rectas (Figura 7F, H). Estomas restringidos a la superficie abaxial de tipo anomotetraciticos de 26.1 a 17.3 µm de longitud y ancho respectivamente, densidad estomática 14/mm2 (Figura 7H). Tricomas eglandulares, unicelulares con base radial de 5 a 7 células pentagonales dispuestos en ambas superficies de 19.54 µm de largo con densidad de 58/mm2 (Figura 7F, G).
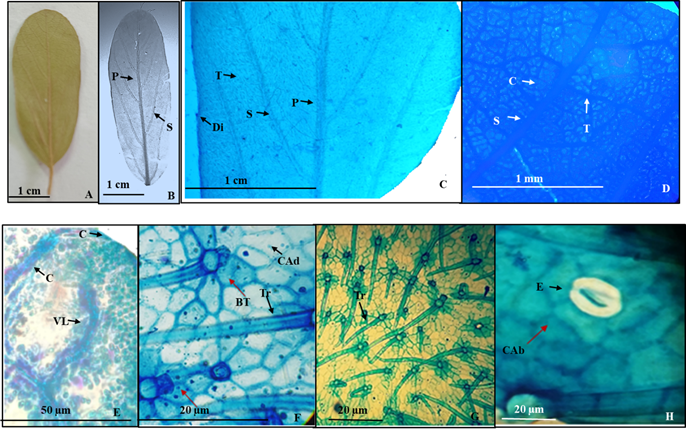
Figura 7 Colubrina stricta. A.- lámina foliar de ejemplar de herbario. B.- Lámina foliar diafanizada con venas de primera y segunda categoría. C.- Detalle de un pequeño diente marginal alimentado por una vena de tercera categoría, venas de primera, segunda y tercera categoría. D.- Detalle de las venas de segunda, tercera y cuarta categoría. E.- Vena de cuarta categoría con una vena incluida. F.- Células epidérmicas adaxiales hexagonales con tricomas de base radial. G.- Células epidérmicas abaxiales con células epidérmicas hexagonales y tricomas con base radial. H. Detalle estoma anomotetracítico. Referencias: P vena de primera categoría. S vena de segunda categoría. T vena de tercera categoría. C vena de cuarta categoría. Tr tricoma. BT base del tricoma. Di diente del margen. VL vena libre. CAd Células de la superficie adaxial. CAb Células de la superficie abaxial. E. Estoma anomotetracítico.
7. - Colubrina texensis (Torrey & Gray) A. Gray (Figura 8).
Hojas simples, simétricas, estrechamente obovadas, obovado-oblongas a anchamente ovadas, de 1 a 3.7 cm de largo y 0.5 a 2 cm de ancho, ápice redondeado, base redondeada a ancha decurrente, textura subcoriacea, margen serrulado con 10 a 30 dientes a cada lado de la lámina foliar (Figura 8A, B, C) sin glándulas (Figura 8D). Peciolo de 1 a 4 mm de largo y 0.25 a 0.50 mm de diámetro. Vena de primera categoría pinnada con curso recto (Figura 8A, B, C), las secundarias craspedródromas, con tres o cuatro venas a cada lado de la lámina foliar, el par inferior ramificado (Figura 8C), en el margen algunas venas no ramificadas y otras 1-ramificadas que alimentan a los pequeños dientes (Figura 8C, D), venas de tercera categoría reticuladas superadyacentes (Figura 8E), las de cuarta reticuladas con venas 2 o más ramificadas (Figura 8F).

Figura 8 Colubrina texensis. A y B.- Láminas foliares de ejemplares de herbario. C.- Lámina de la hoja diafanizada mostrando venas de primera, segunda y tercera categoría. D.- Detalle de un diente del margen alimentado por la ramificación de una vena de segunda categoría. E.- Detalle de venas de tercera y cuarta categoría. F.- Detalle de venas de tercera y cuarta categoría con venas ramificadas. G. Detalle de tricomas con la base radial. H.- Detalles de la base radial de los tricomas. I.- Células epidérmicas adaxiales con paredes onduladas. J.- Células epidérmicas abaxiales con paredes onduladas y estomas anomocíticos. Referencias: P vena de primera categoría. S vena de segunda categoría. T vena de tercera categoría. Tr tricoma. VR venas ramificadas. BT base del tricoma. BR base radial de los tricomas. CAd células de la superficie adaxial. CAb células de la superficie abaxial. E estomas anomocíticos.
Epidermis en vista superficial: Células epidérmicas adaxiales y abaxiales con paredes onduladas (Figura 8H, I). Estomas en la superficie adaxial ausentes, en la abaxial de tipo anomocítico de 21.1 a 18.4 µm de longitud y ancho respectivamente, densidad estomática 55/mm2 (Figura 8I). Tricomas eglandulares, unicelulares con base radial de 3 a 5 células pentagonales dispuestos en ambas superficies de 18.24 µm de largo con una densidad de 42/mm2 (Figura 8G).
En el Cuadro 2 se muestra la comparación entre la arquitectura y superficie foliar de las especies estudiadas del género Colubrina.
Cuadro 2 Comparación de la arquitectura y anatomía foliar de las especies estudiadas del género Colubrina.Table 2. Comparison of the leaf architecture and anatomy of the studied species of the genus Colubrina.
| Especie | Primera categoría | Segunda categoría | Tercera categoría | Cuarta categoría | Alimentación diente | Celulas adaxiales | Celulas abaxiales | Tipos de estomas | Base de los tricomas |
|---|---|---|---|---|---|---|---|---|---|
| C. greggii | Acrodroma basal imperfecta, par inferior lateral ramificadas | Craspedó-dromas, en el margen forman arcos | Reticuladas super-adyacentes | Reticuladas | De los arcos marginales de las venas de segunda cate-goría sale una vena que alimenta a los dientes | Hexago-nales paredes rectas, | Hexago-nales, paredes rectas | Anomo-cíticos | Base engrosada y células epidérmicas no modificadas |
| C. angustior | Pinnada | Craspedó-dromas, primer par basal ramificado | Reticuladas super-adyacentes | Reticuladas | Venas tercera categoria alimentan los dientes del margen, pueden o no formar arcos | Hexago-nales, paredes rectas | Paredes onduladas | Anomo-cíticos | Base engrosada y células epidérmicas no modificadas |
| C. yucatensis | Pinnada | Craspedó-dromas, primer par basal ramificado | Reticuladas super-adyacentes | Reticuladas | Venas tercera categoría alimentan los dientes del margen a veces forman arcos | Poligona-les, paredes rectas | Paredes onduladas | Anomo-cítico | Radial |
| C. macrocarpa | Acrodroma basal imperfecta | Craspedó-dromas no ramificadas o 1-ramificadas | Reticuladas, super-adyacentes, incluyen venas libres | Venas de segunda y tercera categoría alimentan los dientes del margen | Alargadas paredes rectas | Pentago-nales, paredes rectas | Aniso-cíticosy peri-cítico | Radial | |
| C. lanulosa | Pinnada | Craspedó-dromas, par inferior lateral ramificadas | Reticuladas, en el margen alimentan los dientes | Reticuladas | Venas de segunda y tercera categoría alimentan los dientes del margen | Anticlina-les con paredes rectas | Anticlina-les con paredes rectas | Anomo-citicos y anomo-tetra-citicos | Radial |
| C. stricta | Pinnada | Craspedó-dromas, primer par basal no ramificado | Reticuladas, super-adyacentes | Reticuladas con 1 vena no ramificada | Venas de tercera categoría alimentan los dientes del margen | Hexago-nales paredes rectas | Hexago-nales paredes rectas | Anomo-tetra-citicos | Radial |
| C. texensis | Pinnada | Craspedó-dromas, primer par basal ramificado | Reticula-das super-adyacentes | Reticula-das con venas 2 o más ramificadas | Venas de segunda categoría y muy pocas de tercera alimentan los dientes del margen | Paredes onduladas | Paredes onduladas | Anomo-cítico | Radial |
Al analizar el resultado de la arquitectura y anatomía foliar de las especies y variedades estudiadas se encontraron algunos patrones que las relacionan, pero a la vez, se observaron diferencias que permitieron separarlas como se muestra en la siguiente clave dicotómica.
Clave dicotómica para separar las especies y variedades del género Colubrina que crecen en México, según su arquitectura y anatomía foliar
1.- Venas de primera categoría acródroma basal imperfecta, las secundarias craspedódromas
2.- Tricomas de la lámina foliar con base engrosada y células epidérmicas no modificadas,
venas de tercera y cuarta categoría reticuladas, estomas anomocíticos
…………………C. greggii
2.- Tricomas de la lámina foliar con base radial, venas de tercera categoría, incluye venas
libres no ramificadas, estomas de tipos pericíticos y anomocíticos
…..………..…C. macrocarpa var. macrocarpa
1.- Vena de primera categoría pinnadas, las secundarias craspedódroma
3.- Tricomas de la lámina foliar con base engrosada y células epidérmicas no modificadas
………….………….C. angustior
3.- Tricomas de la lámina foliar con base radial
4.- Par basal de las venas secundarias ramificadas
5.- Venas de cuarta categoría reticuladas con venas dos o más veces ramificadas, células
epidérmicas adaxiales y abaxiales con paredes onduladas ……………………C. texensis
5.- Venas de tercera y cuarta categoría reticuladas sin venas libres incluidas
6.- Lámina foliar densamente pilosa con células epidérmicas adaxiales y abaxiales con
paredes rectas, estomas anomocíticos y anisocíticos ……………………… C. lanulosa
6.- Lámina foliar pilosa, células de la epidermis adaxiales con paredes rectas y las abaxiales
con paredes onduladas, estomas anomocíticos .……………………...…C. yucatanensis
4.- Par basal de venas secundarias no ramificadas, venas de cuarta categoría con una vena
no ramificada ……………………………………………………………….… C. stricta
Discusión
La base de los tricomas es un carácter importante que permitió separar las especies, la mayoría de ellas presentan tricomas con base radial y solo Colubrina greggii y C. angustior muestran tricomas con base engrosada y células epidérmicas no modificada semejantes a los que presentan el género Gouania, Rhamnaceae (Hérnández et al., 2021).
Johnston (1971, p. 8) en las claves para separar los taxones de los subgéneros de Colubrina, en especial el subgénero Serrataria incluye a los siete taxones estudiados en este trabajo, menciona que las especies presentan el margen de las hojas serrado con más de diez dientes y cada uno de ellos está asociados con glándulas marginales, sin embargo, en los materiales diafanizados de este trabajo no se apreciaron glándulas en los dientes y las pequeñas protuberancias corresponden a las venas secundarias o terciarias que alimentan a cada uno de ellos.
En este mismo trabajo de Johnston (1971, p. 37) reconoció tres grupos de especies con sus respectivas variedades; Colubrina greggii con tres, C. texensis con dos y C. macrocarpa con tres. Sin embargo, Nesom (2013 p. 11) consideró las tres variedades de C. greggii como tres especies y las separa por la forma de las hojas, el tamaño de la planta y por el número de flores que presentan las inflorescencias así como la distribución geográfica. A partir de los caracteres de arquitectura foliar y anatomía de la epidermis fue posible separar estos taxones por los siguientes caracteres:
En los resultados de este trabajo C. greggii mostró venación acródroma basal imperfecta, tricomas de la lámina foliar con la base engrosada y células epidérmicas no modificadas, además presenta células de la epidermis adaxiales y abaxiales hexagonales con paredes rectas. C. angustior con venación craspedódroma, tricomas con base engrosada y células epidérmicas no modificadas, células adaxiales y abaxiales hexagonales con paredes rectas y C. yucatanensis con el mismo tipo de venación, tricomas de la lámina foliar con base radial, células epidérmicas adaxiales poligonales con paredes rectas y las abaxiales con paredes onduladas. Lo anterior justifica el reconocimento de las especies propuestas por Nesom (2013); en donde el complejo de C. greggii presenta caracteres en común como los estomas anomocíticos y las venas de tercera y cuarta categoría reticuladas.
Johnston (1971, p. 43) reconoció las variedades de Colubrina texensis var. texensis y C. texensis var. pedunculata, aunque anteriormente Johnston (1969) consideró a este último taxón de origen híbrido entre C. texensis y C. greggii, pero decidió dejarla como la variedad de C. texensis var. pedunculata. Posteriormente Engelmann decide describirla como C. stricta Engelmann ex M.C. Johnston (1969, p. 257), por lo que Colubrina texensis var. pedunculata pasa a ser sinónimo de C. stricta.
Posteriormente Nesom (2013, p. 1) las consideró como dos especies, una de ellas como C. texensis antes considerada como la variedad tipo y la segunda la valida como Colubrina stricta argumentando que C. texensis y C. stricta son parcialmente simpátricas, pero morfológicamente distintas.
Con respecto al punto anterior, Nesom (2013) separa a C. texensis y C. stricta por las características del tamaño de las flores, largo del pedicelo, tipos de inflorescencia, ramas derechas o en zigzag y la distribución geográfica. Desde el punto de vista de la arquitectura foliar y las características epidérmica se separan por los siguientes caracteres: C. texensis con el par basal de venas secundarias ramificadas, venas terciarias reticuladas y las cuaternarias con venas dos o más veces ramificadas, células epidérmicas adaxiales y abaxiales con paredes onduladas, estomas anomocíticos; mientras que C. stricta no presenta el par basal de venas secundarias ramificadas, venas de cuarta categoría con una vena no ramificada, células epidérmicas adaxiales y abaxiales hexagonales con paredes rectas, estomas anomotetracitico.
Lo anterior justificaría la separación de las especies propuestas por Nesom (2013). El complejo de C. texensis tiene en común las venas de primera categoría pinnadas y las venas de segunda categoría craspedódromas, las venas de tercera categoría reticuladas y las de cuarta incluyen venas no ramificadas o dos o más veces ramificadas, tricomas con base radial.
Con lo que respecta a las variedades de Colubrina macrocarpa, Johnston (1971, p. 41) reconoce tres, la variedad tipo y las otras son C. macrocarpa var. lanulosa y C. macrocarpa var. macrocarpiodes, esta última conocida solo de la localidad tipo en Querétaro entre San Juan del Río y Hacienda Ciervon en el municipio de Cadereyta, recolectada por Rose et al. en 1905 y no se ha vuelto a encontrar, por lo que se excluye en este trabajo por la falta de material foliar. Las variedades de Colubrina macrocarpa var. macrocarpa y C. macrocarpa var. lanulosa, Johnston (1971, p. 42) las separa por las siguientes características:
C. macrocarpa var. macrocarpa son arbustos de 1 a 1.5 m, lámina foliar 1.1 a 1.4 veces más largas que anchas, superficie adaxial con pubescencia uniforme, diminuta, estrigosa, sólo cerca de las venas con un tomento algo apretado de tricomas sedosos rojizos; pecíolos de 1 a 1.5 mm de espesor; inflorescencia tirsoide de10 a 30 flores, de alrededor de 1 cm de largo, en cambio C. macrocarpa var. lanulosa son arbustos de hasta 3 m de altura, láminas foliares 1.3 a 1.6 veces más largas que anchas, superficie adaxial densamente rojizo-estrigoso, superficie abaxial con un tomento denso, suelto, rojizo-sedoso; pecíolos de 1.5 a 2 mm de espesor; inflorescencia tirsoide de 20 a 50 flores, de alrededor de 2 cm de largo.
Las características de arquitectura y anatomía foliar que se encontraron es este trabajo son las siguientes:
C. macrocarpa var. macrocarpa con venación acródroma basal imperfecta, venas de tercera categoría con una vena libre incluida, estomas anisocíticos con algunos pericíticos y C. macrocarpa var. lanulosa presenta venación craspedródroma, venas de tercera y cuarta categoría reticuladas, estomas anisocíticos y algunos anomotetracíticos. La arquitectura y anatomía foliar permite separar estas dos variedades, por lo que se propone una combinación y estatus de una nueva especie de Colubrina lanulosa (M. C. Johnston) Fernández & Arreguín y como sinónimo a C. macrocarpa var. lanulosa.
Colubrina lanulosa (M. C. Johnston) Fernández & Arreguín, comb. et stat. nov.
Colubrina macrocarpa (Cav.) G. Don var. lanulosa (S. F. Blake) M. C. Johnston, Brittonia 23: 41. 1971. Tipo: MEXICO. Guerrero. Volcanic soil, Tlacotepec, ca. 17°40’ N lat., 100°00’ W long., 1600 m, 10 jun 1899, Langlassé 1053 (HOLOTYPO GH!).
Arbustos de hasta 3 m de altura, hojas simples, simétricas, oblonga a oblonga-lanceolada de 3 a 9 cm de largo y 2 a 7 cm de ancho, ápice agudo, base redondeada, textura subcoriácea, margen serrado con más de 50 dientes de cada lado o margen crenado, sin glándulas. Peciolo de 1.2 a 2 mm de largo y 1 a 1.5 mm de diámetro, superficie adaxial densamente rojizo-estrigoso, superficie abaxial con un tomento denso, suelto, rojizo-sedoso; pecíolos de 1.5-2 mm de espesor; inflorescencia tirsoide de 20-50 flores, de 2 cm de largo.
Venas de primera categoría pinnada de recorrido recto, las secundarias craspedródromas, cinco de cada lado, curveadas, el primer par inferior ramificado, las venas secundarias en el margen son 1-ramificada y alimentan a los pequeños dientes o crenaciones, las terciarias reticuladas superadyacentes y las marginales alimentan los dientes, las venas cuaternarias reticuladas. Epidermis en vista superficial con células en ambas superficies de la lámina foliar anticlinales. Estomas abaxiales mayoritariamente de tipo anisocítocos y algunos anomotetracíticos de 27.1 µm de longitud y 24 µm de ancho, densidad estomática 29/mm2. Tricomas eglandulares, unicelulares muy densos en ambas superficies de la lámina foliar insertos en las venas de primera, segunda, tercera y cuarta categoría, con base radial de 6 células pentagonales dispuestos en ambas superficies de 25.32 µm con 102/ mm2.
Distribución: Se le conoce solo del estado de Guerrero.
Colubrina lanulosa (M. C. Johnston) Fernández & Arreguín, comb. et stat. nov.
Shrubs up to 3 m high, leaves simple, symmetrical, oblong to oblong-lanceolate, 3 to 9 cm long and 2 to 7 cm wide, acute apex, rounded base, subcoriaceous texture, serrated margin with more than 50 teeth, each side or margin crenate, without glands. Petiole 1.2-2 mm long and 1-1.5 mm in diameter, adaxial surface densely reddish-strigous, abaxial surface with a dense, loose, reddish-silky tomentum; petioles 1.5-2 mm thick; thyrsoid inflorescence of 20-50 flowers, 2 cm long.
First category pinnate veins with a straight course, the secondary ones craspeddromous, five on each side, curved, the first lower pair branched, the secondary veins on the margin are 1-branched and feed the small teeth or crenations, the tertiary reticulated superadjacent and the marginal ones feed the teeth, the quaternary reticulated veins. Epidermis in superficial view with anticlinal cells on both surfaces of the leaf blade. Abaxial stomata, mostly anisocytotic and some anomotetracytic, 27.1 µm in length and 24 µm in broad, stomatal density 29/mm2. Eglandular, single-celled, very dense trichomes on both surfaces of the leaf blade inserted in the veins of the first, second, third and fourth category, with a radial base of 6 pentagonal cells arranged on both surfaces of 25.32 µm with 102/ mm2.
Distribution: It is known only from the state of Guerrero.
Colubrina lanulosa (M. C. Johnston) Fernández & Arreguín, comb. et stat. nov.
Frutices usque ad 3 m alta, foliis simplicibus, symmetricis, oblongis vel oblongo-lanceolatis, 3 ad 9 cm longis et 2 ad 7 cm latis, apice acutis, basi rotundatis, textura subcoriaceis, margine serratis plusquam 50 dentibus. margine crenata, sine glandulis. Petiolum 1.2 ad 2 mm longum et 1 ad 1.5 mm diametro, superficie adaxiali dense rufo-strigore, superficie abaxiali denso, laxo, tomento rubro-sericeo; petioli 1,5 ad 2 mm crassi; inflorescentia thyrsoidea 20 ad 50 florum, 2 cm longa.
Prioris generis venae pinnatae recto cursu, secundariae craspeddromae, utrinque quinque, curvatae, primo inferiores par ramosae, venae secundariae in margine 1-ramosae sunt et dentes vel crenationes parvos, tertiariis reticulatis superadiacentibus pascunt marginales dentes pascunt, venae quaternariae reticulatae. Epidermis in visu superficiali cum cellulis anticlinalibus utrinque foliorum laminae. Stomata abaxialia, plerumque anisocytotica et quaedam anomotetracytica, 27.1 µm longitudine et 24 µm lata, densitas stomatalis 29/mm2. Eglandulare, unico-cellulatum, trichomatum densissimum in utraque parte laminae folii in venis primae, secundae, tertiae et quartae insertae, cum basi radiali cellularum 6 pentagonarum in utraque superficie 25.32 µm cum 102/ mm2 dispositae.
Distributio: Ex statu Guerrero notum est.
Conclusiones
Fue posible separar los siete taxones estudiados utilizando caracteres de arquitectura y superficie foliar como tipo de venación, base de los tricomas, características de las células epidérmicas adaxiales y abaxiales y los tipos de estomas.
Se apoya la separación de especies propuestas por Nesom (2013) debido a que los datos que aporta la arquitectura foliar y la superficie epidémica permitieron separar estos taxones.
Se propone una combinación y estatus de una nueva especie a Colubrina lanulosa (M. C. Johnston) Fernández & Arreguín.

