











 text new page (beta)
text new page (beta) Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkIntroducción
La esclerosis múltiple (EM) se considera una de las causas más comunes de discapacidad neurológica en adultos jóvenes. Consiste en una enfermedad crónica e inflamatoria del sistema nervioso central (SNC), caracterizada por desmielinización axonal, gliosis y degeneración neuronal. Una hipótesis patógena relaciona la EM con respuestas disinmunes, con la participación de autoanticuerpos, complemento y mediadores innatos de respuesta inmunitaria1. Los fármacos inmunosupresores se consideran terapias modificadoras de la enfermedad, ya que pueden reducir la discapacidad, la frecuencia de recaída, la gravedad y la duración de los síntomas.
Desafortunadamente, las terapias modificadoras de la enfermedad no son curativas y, en ocasiones, no evitan el cambio a cursos progresivos o, una vez alcanzados, la progresión de la discapacidad2.
Las terapias celulares representan una nueva frontera en el tratamiento de enfermedades neurológicas. En este contexto, las células madre mesenquimales adultas (aMSC) han demostrado su potencial terapéutico en varios modelos de enfermedades. Debido a que las aMSC se comportan de forma heterogénea según el tejido de origen y las condiciones de cultivo. La International Society for Cell Therapy en el año 2006, estableció unas pautas para la caracterización de acuerdo a tres criterios3: 1) crecimiento en adherencia a las superficies plásticas utilizadas para su cultivo; 2) expresión de las moléculas de superficie (CD73, CD90 y CD105), y la ausencia de marcadores de monocitos y macrófagos (CD11b y CD14), marcador leucocitario (CD45), marcador de células B (CD19 o CD79a), marcador de progenitores hematopoyéticos y marcadores de células endoteliales (CD34), y HLA-DR; y 3) capacidad de diferenciación in vitro hacia osteoblastos, adipocitos y condroblastos.
El beneficio ejercido por las aMSC se ha asociado con la capacidad de diferenciarse en varios linajes celulares, segregar factores solubles cruciales para la proliferación y la supervivencia celular4, modular la respuesta inmune5 y migrar al sitio de la lesión6. Dado que el daño tisular derivado de la inflamación es un proceso clave desencadenado en respuesta a una lesión y enfermedad, las aMSC podrían convertirse en un tratamiento para cualquier daño tisular u orgánico asociado con una actividad inflamatoria intensa5.
Se han diseñado múltiples modelos de EM en investigación experimental para lograr una mejor comprensión de la patogénesis de la enfermedad y los mecanismos de progresión. La encefalomielitis (encefalitis autoinmune experimental, EAE) es un modelo animal de EM basado en la inmunización activa que reproduce clínicamente la lesión inflamatoria que se produce en la EM. La neuritis óptica, una inflamación del nervio óptico (NO) y la lesión progresiva de la médula espinal se han descrito en ratones de la cepa B6/C57 tras la inmunización con un fragmento de la glicoproteína mielínica del oligodendrocito (MOG) que comprende de 35 a 55 residuos de aminoácidos (MOG35-55). Esta lesión incluye la desmielinización con daño axonal posterior y la pérdida neuronal que también afecta al ojo, en particular a las células ganglionares de la retina (CGR)7. De hecho, se ha demostrado que la gravedad del curso de la EAE se correlaciona con los signos de desmielinización e inflamación en el NO8.
Objetivo
Estudiar la respuesta neuroprotectora y neurorregeneradora de las aMSC sobre las CGR y el NO en un modelo animal de EM.
Material y métodos
Los animales
Se utilizaron 24 ratones hembra B6/C57 de 10 semanas de edad. Los animales se dividieron en tres grupos: grupo control (10 animales), grupo EAE (7 animales) y grupo EAE-MSC (animales con EAE tratados con aMSC, administradas por vía intravenosa) (7 animales). Los posibles efectos de la inyección de aMSC en animales control se descartó durante el proceso de desarrollo y optimización del modelo animal. Estos animales fueron utilizados para la experimentación animal según las directrices éticas de experimentación animal. Los animales con EAE se inyectaron con MOG35-55 (200 µg de péptido/animal en 200 µl intraperitoneal)9, se monitorizaron cada 24-48 h y se les evaluó la discapacidad motora. Al grupo EAE-MSC se le inyectó 1 x 106 aMSC10 junto con una proteína verde fluorescente (GFP) por vía intravenosa en la vena de la cola. Consideramos el inicio de la enfermedad como la aparición de debilidad de la cola distal y/o leve movimiento irregular. Cuando los animales cumplieron con los criterios de punto final (cuando el animal es incapaz de conseguir alimento o agua), se sacrificaron mediante eutanasia. Posteriormente, se perfundieron a través del ventrículo izquierdo del corazón con 50 cc de solución salina estéril (NaCl al 0.9%), seguido de 150 cc de paraformaldehído (PFA) al 4% o glutaraldehído al 2% de PFA-2.5% solución. Más tarde, los globos oculares y los NO se extrajeron y, posteriormente, se fijaron en el mismo fijador durante 24 h. Luego, se lavaron con solución salina tamponada con fosfato (PBS) 0.1 M. Finalmente, las muestras se mantuvieron a 4 °C en PBS 0.1 M con azida de sodio al 0.05% para evitar la contaminación bacteriana y por hongos.
Crecimiento, caracterización y transfección de células madre mesenquimales adultas con proteína verde fluorescente
Todas las muestras fueron recogidas como un producto de cirugía plástica de pacientes sometidos de forma electiva a procedimientos de lipoaspirados quirúrgicos en la Clínica Cervera Sebastián (Valencia, España). Se entregó un formulario de consentimiento informado a los pacientes para que fuera firmado y almacenado en la base de datos CELULIFE, tras recibir información del estudio. El uso de muestras de tejido adiposo fue aprobado por el Comité de Ética correspondiente (CAEC, Comité Autonómico de Ensayos Clínicos, Valencia, España). Los participantes no recibieron compensación financiera. El estudio siguió en todo momento las leyes y regulaciones vigentes, incluyendo los principios éticos derivados de la Declaración de Helsinki, la política internacional sobre la realización de estudios clínicos recogida en las International Guidelines for Ethical Review of Epidemiological Studies (Council for the International Organizations of Medical Sciencies-CIOMS-Ginebra, 2009) y las Recomendaciones sobre Buena Práctica Clínica.
El aislamiento de las células madre derivadas de tejido adiposo se llevó a cabo utilizando un método mecánico y enzimático. La mezcla de digestión estaba compuesta por Dulbecco’s Phosphate-Buffered Saline (DPBS) pH 7.5 (Life technologies Cat nº10010056) con 1 mg/ml de colagenasa tipo I (Gibco Cat Nº 17100-017) y el mismo volumen de tejido adiposo lipoaspirado. El tejido era digerido durante 30 minutos a 37 °C en condiciones de agitación, y más tarde se filtró para separar tejido no digerido y centrifugado a 500 g durante 10 minutos a temperatura ambiente. El sobrenadante (adipocitos flotantes) se separó del sedimento (fracción de células madre derivadas de tejido adiposo). Las células se resuspendieron en un tampón de lisis de eritrocitos (BD Cat # 555900), se incubaron a temperatura ambiente durante 10 minutos y se centrifugó a 500 g durante 10 minutos a 20 °C. Finalmente, las células fueron resuspendidas y expandidas en medio de cultivo Dulbecco’s Modified Eagle Medium (DMEM) con bajo contenido de glucosa (Gibco 31885023), suplementado con 10% de suero humano y 1% (v/v) de penicilina/estreptomicina (10,000 U/ml de penicilina, 10,000 µg/ml de estreptomicina, Gibco Life Technologies, n.°15140-122). Las células madre derivadas del tejido adiposo se cultivaron a 37 °C en una atmósfera de aire con 5% de CO2 con cambios medios cada 3 días. Cuando las células alcanzaron en el matraz aproximadamente el 80% de confluencia, el subcultivo (pase) se realizó mediante tripsinización, se contó y se volvió a colocar en una caja de cultivo celular T75 tratadas con polilisina para crecimiento celular en mono capa a una densidad de 5,000 células/cm2. Para la tripsinización, el medio de cultivo se aspiró y el matraz se lavó usando PBS. Luego, se desechó la solución y se añadió tripsina 6 ml/matraz y se dejó durante 5 minutos. La tripsina se neutralizó utilizando la misma cantidad de medio completo. La suspensión celular se centrifugó (500 g durante 7 minutos) y se aspiró su sobrenadante. El sedimento se resuspendió en medio completo y las células se contaron utilizando la prueba de exclusión de azul tripán (hemocitómetro). En el cuarto paso, se transfectaron por electroporación con un plásmido que expresaba GPF. El vector de expresión se construyó mediante la inserción en el vector vacío obtenido comercialmente (Invitrogen) pCEP4 el gen de la GPF. De este modo, el plásmido se introdujo en la concentración adecuada, y 24 horas después fue posible determinar la eficacia del proceso mediante detección por fluorescencia. Esta placa se incubó durante 3 días a 37°C con 5% de CO2. Las células se caracterizaron completamente como aMSC por inmunofenotipo con citometría de flujo11.
Procesado del nervio óptico y la retina para el estudio histológico
Se obtuvieron secciones longitudinales de globo ocular (400 µm de espesor) con un vibrátomo (Vibratomo Leica 1000mM Biosystems). Las secciones del NO se seleccionaron a su salida del globo ocular y se sometieron a dos procedimientos diferentes: 1) crioprotección en sacarosa al 30%, congelación rápida con nitrógeno líquido y secciones histológicas de 10 µm de espesor con un criostato (CM1900, Leica, Heidelberg, Alemania), las secciones se tiñeron con hematoxilina-eosina y azul de toluidina; 2) inclusión en resina epoxi y secciones de 1.5 µm-semifinas con un ultramicrotomo (Ultracut, Leica ultracut-MZ6). La glía residente y reactiva se detectó en secciones congeladas de 10 µm mediante inmunomarcaje con anticuerpo policlonal de conejo anti-GFAP (proteína ácida fibrilar glial) (#12692777, Fisher Scientific, Hampton, New Hampshire, EE.UU.) diluido 1:200, se incubaron diluidas en tampón de bloqueo (5% normal goat serum (NGS), 5% caseína en PBS) durante toda la noche a 4 ºC. Se lavaron las muestra en PB 0.1M, y se realizó la incubación del anticuerpo secundario (cabra anti-conejo alexa 555, #A21428, Fisher Scientific) diluido 1:500 en 5% (NGS) en PBS durante 1 hora a temperatura ambiente, conjugado con fluorescencia. A continuación se lavaron las muestras en PB 0.1M y se montaron con medio acuoso (Fluorsave®).
Análisis cuantitativos de muestras histológicas e inmunohistoquímicas
En las secciones teñidas con hematoxilina-eosina/azul-toluidina, se realizaron 246 fotos, se seleccionaron 3 secciones por muestra y se realizaron 3-4 fotos por sección, dependiendo de la presencia del NO o la retina, o de la presencia de ambos. Finalmente, se tomaron y analizaron 80 fotos del grupo de control, 84 fotos del grupo EAE y 82 fotos del grupo EAE-MSC. Los números de CGR por unidad de longitud se midieron en una distancia promedio de 300 µm a ambos lados de la entrada del NO, un área de referencia con menos variabilidad anatómica para permitir recuentos reproducibles en muestras distintas. Los CGR se contaron manualmente.
Para estudiar la inflamación endógena, se detectaron células gliales residentes y reactivas con anticuerpos anti-GFAP. El total de células positivas a GFAP por micrón se contó en 3 secciones/animal por grupo y se analizaron las diferencias entre ellas.
Análisis cuantitativos bajo microscopía electrónica: G-ratio y densidad axonal
Para los estudios cuantitativos de microscopia electrónica de transmisión (MET), los NO se seccionaron transversalmente. De todas las secciones semifinas, se seleccionaron aquellas con NO y retina, y luego se obtuvieron secciones ultrafinas, de 70-80 nm, se incluyó en resina (Leica EM-UC6) para completar el análisis morfológico con un estudio ultraestructural bajo el microscopio electrónico (microscopia electrónica de transmisión FEI-Tecnai-Spirit-G2, Oregon, EE.UU.).
La G-ratio se obtuvo como la relación entre el diámetro axonal interno (excluyendo la mielina) y el diámetro exterior total (incluida la vaina de mielina). En las secciones ultrafinas del NO, se estudiaron más de 100 axones por grupo y se clasificaron como normales (proporción G:0.74-0.84)12, axones remielinizantes (proporción G:0.85-0.94) y axones con daño severo a la mielina (proporción G:0.95-1). Se obtuvieron 103 micrografías con microscopía electrónica de transmisión (48 fotos del grupo de control, 25 fotos del grupo EAE y 30 fotos del grupo EAE-MSC). La densidad axonal se calculó como el número de axones por micrómetro cuadrado en cuatro secciones por animal. Esta medida se obtuvo como la media promedio de dos observadores independientes cegados (RGG, LCN).
Análisis estadístico
Debido a que nuestro tamaño de muestra excluyó las pruebas paramétricas, realizamos la prueba U de Mann-Whitney para estudiar la significación estadística entre grupos experimentales. Las diferencias entre los grupos analizados se consideraron significativas si el valor de p < 0.05. Para comparaciones de más de dos grupos, utilizamos la prueba de Kruskal-Wallis.
Resultados
Los animales tratados con células madre mesenquimales adultas presentaban un periodo libre de enfermedad mayor que los animales no tratados
Los animales con EAE experimentaron los primeros signos clínicos desde el día 7 hasta el día 19 después de la inmunización. Estos síntomas consistían en déficits motores secundarios a la lesión de la médula espinal y eran progresivos, ya que nuestro modelo reproducía un curso crónico9. El inicio consistió en un déficit focal con pérdida del tono muscular de la cola, seguido de una paresia progresiva ascendente que condujo a criterios de punto final en distintos puntos temporales (los animales fueron llevados a eutanasia por consideraciones éticas). El curso clínico fue variable, con una esperanza de vida promedio de 57 días (rango 30-70). Aunque hubo un retraso en el inicio de la enfermedad en el grupo EAE-MSC en comparación con el grupo EAE (10 vs.13 días), los valores de p no fueron estadísticamente significativos (p = 0.07). En un momento dado (45 días a partir de la inmunización), los animales en el grupo EAE alcanzaron una discapacidad máxima promedio de 3.5 puntos, se evidenció la instauración de la enfermedad, con una afectación motora grave, parálisis de las cuatro extremidades, que dificultó al animal la obtención de alimentos y agua, mientras que los animales del grupo tratado con aMSC presentaron una puntuación máxima de 2.5 puntos, la enfermedad se manifestó con la parálisis de dos extremidades (las dos traseras o una delantera y una trasera). Todos los animales cumplieron los criterios de punto final y fueron sacrificados para su procesamiento.
La inmunización con encefalitis autoinmune experimental condujo a cambios morfológicos y disminuyó el número de células ganglionares de la retina con un efecto favorecedor tras la administración de células madre mesenquimales adultas
Las aMSC se transfectaron con un plásmido que expresaba GFP para poder detectar aMSC injertadas mediante inmunofluorescencia. No pudimos encontrar aMSC como células GFP+ en nuestras muestras de la retina o el NO, sin embargo, observamos cambios en la viabilidad neuronal, la mielinización y la inflamación local, como lo describimos con más detalle.
En la EAE y en los grupos tratados, la morfología retiniana a bajo aumento no difirió de la de los ratones control (Figs. 1 A-C). Sin embargo, el análisis con MET mostró que los CGR en el grupo control eran células grandes con núcleos redondeados y un núcleo central con cromatina dispersa, abundante retículo endoplásmico rugoso y microtúbulos (Fig. 1 D). En contraste, en los grupos de EAE y tratados, el citosol de las CGR fue más oscuro, con núcleos más pequeños y con grandes invaginaciones, muy evidentes, que alcanzaron hasta el centro del núcleo (Fig. 1 E) y que perdieron su esfericidad adquiriendo una forma arriñonada (Fig. 1 F). Presentaron un citoesqueleto desorganizado que sugirió que las CGR estaban dañadas (Figs.1 E y F). Las células gliales adyacentes caracterizadas por citosol ligero, filamentos intermedios y núcleos invaginados se observaron con mayor frecuencia en los grupos de EAE y tratados en comparación con el control. Las células microgliales con citosol oscuro, forma invaginada, cisternas largas y lisosomas fueron más abundantes en las regiones estudiadas de ratones inmunizados (Figs. 1 J y K). Ambos tipos de células también se observaron con inmunohistoquímica contra GFAP, una proteína de los filamentos intermedios presentes en los astrocitos (Figs. 1 L y M).
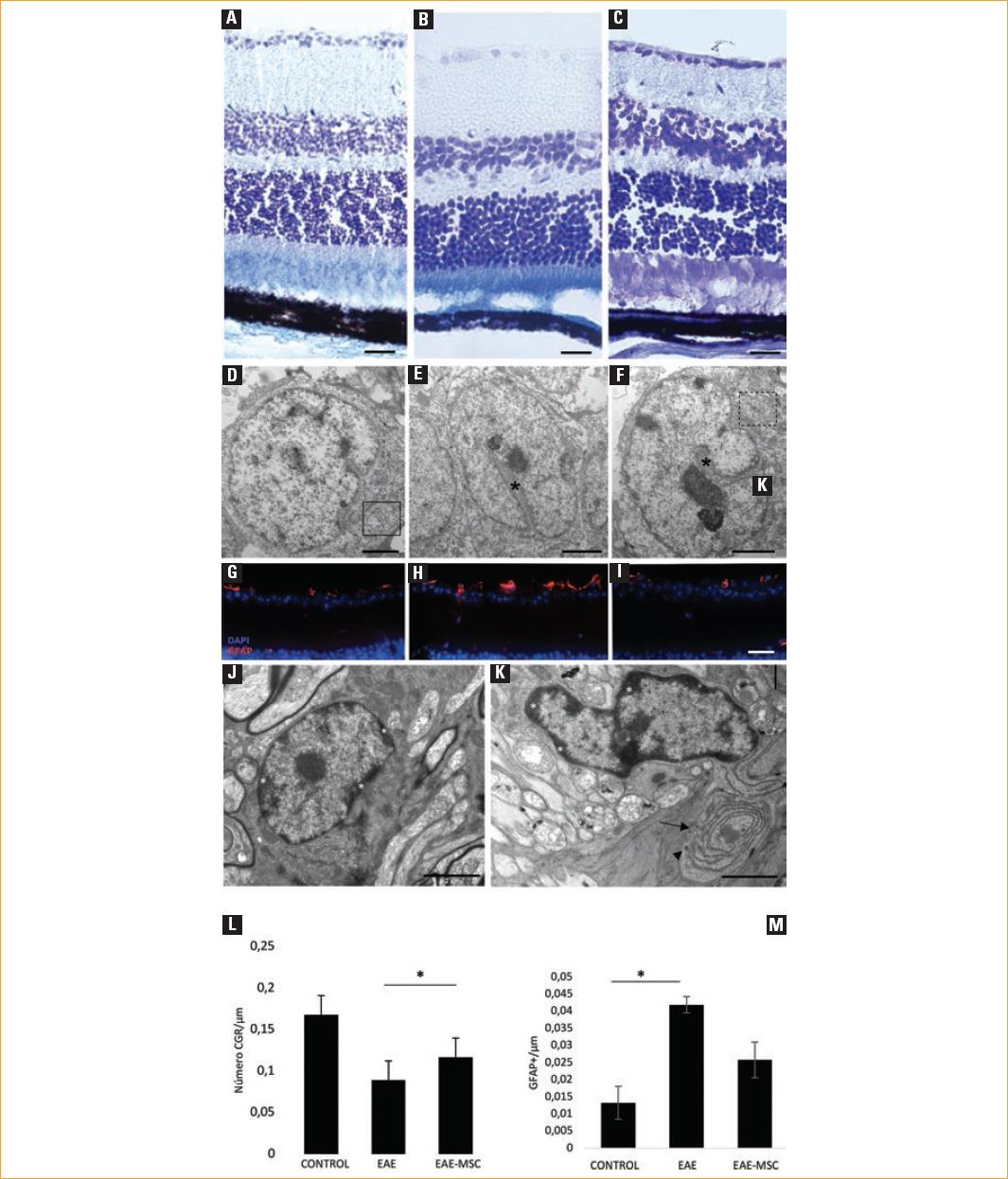
Figura 1 Estudio comparativo histológico y ultraestructural de la retina. A-C: secciones de criostato de la retina teñidas con azul de toluidina en el grupo control (A), grupo de encefalitis autoinmune experimental (EAE) (B) y grupo de EAE en tratamiento con células madre mesenquimales adultas (EAE-MSC) (C) (escala 10 µm). D-F: micrografías, secciones ultrafinas, estudio ultraestructural de las células ganglionares de retina (CGR) en el grupo control (D), donde muestran un núcleo grande y redondeado, cromatina dispersa, abundante retículo endoplásmico y microtúbulos característicos (recuadro), grupo EAE (E) y grupo EAE-MSC (F). La morfología de las CGR está alterada en el grupo tratado con EAE, y el grupo MSC muestra núcleos invaginados (asterisco) y citoesqueleto más desorganizado (recuadro discontinuo) (escala 2 µm). G-I: la expresión de proteína ácida fibrilar glial (GFAP) (rojo) en la capa de CGR en el grupo control (G), grupo EAE (H) y grupo EAE-MSC (I) (escala 20 µm). J,K: células de microglía, se caracterizan por un núcleo irregular, con condensación de cromatina en la periferia (asteriscos), con grandes cisternas alargadas (flecha) y lisosomas secundarios (punta de flecha) (escala 2 µm). L: evaluación cuantitativa de CGR por µm en los tres grupos estudiados. M: evaluación cuantitativa del número de células gliales (GFAP+) en la retina por µm.
Las CGR se alteraron morfológicamente en ratones inmunizados, pero para evaluar si los cambios morfológicos estaban relacionados con la pérdida de células, cuantificamos el número de CGR por unidad de longitud en secciones de retina longitudinales en todos los grupos. En línea con nuestros datos anteriores, el número de CGR en el grupo de control mostró un promedio de 0.1678 CGR/µm. El grupo EAE y el grupo tratado con aMSC mostraron una disminución significativa en el número de CGR con valores de 0.0891 CGR/µm (p = 0.000035) y 0.116 (p = 0.0002) CGR/µm, respectivamente, en comparación con los ratones de control. Cuando se compararon los números de CGR entre EAE y el grupo tratado con aMSC, las diferencias también fueron significativas (p = 0.01) (Fig. 1 L).
La inmunohistoquímica contra GFAP mostró un aumento en las células GFAP+ en el grupo EAE (0.0419 células GFAP+/µm) y en el grupo tratado (0.0322 células GFAP+/µm) en comparación con el grupo control (0.0132 células GFAP+/µm) (Fig. 1 M). Estos resultados mostraron un aumento significativo en las células GFAP+ en animales inmunizados en comparación con los ratones de control (p = 0.038) y una respuesta parcial al tratamiento con aMSC que, sin embargo, no alcanzó la significación estadística (p = 0.068).
La inflamación endógena y la desmielinización en el nervio óptico de ratones inmunizados se rescató parcialmente con células madre mesenquimales adultas intravenosas
La observación de la morfología de las secciones longitudinales del NO con tinción histológica a bajo aumento mostró que en el grupo de EAE el nervio adoptó contornos irregulares con regiones estrechas demarcadas, que se correspondían con áreas de mayor densidad celular (flechas Fig. 2 C), en comparación con la ausencia de estrechamientos del NO en los ratones del grupo control (Fig. 2 A).
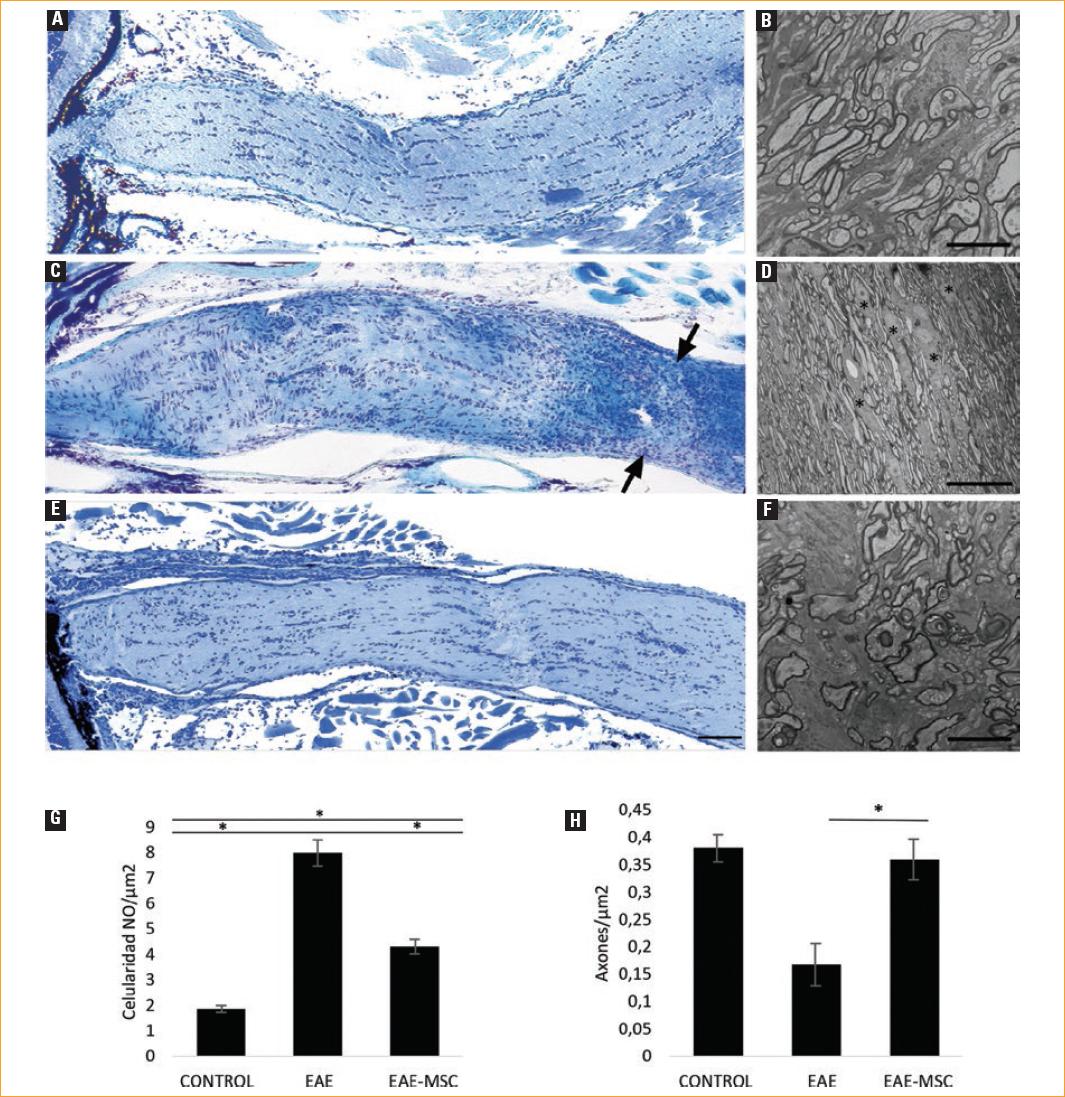
Figura 2 Estudio comparativo histológico y ultraestructural del nervio óptico (NO). A,C,E: secciones de criostato del NO teñidas con azul de toluidina en el grupo control (A), grupo de encefalitis autoinmune experimental (EAE) (C) y grupo de EAE en tratamiento con células madre mesenquimales adultas (EAE-MSC) (E); flechas negras: contornos irregulares focales, con incremento de la densidad celular, escala 200 µm. B,D,F: estudio ultraestructural de los axones del NO en el grupo control (B), grupo EAE (D) y grupo EAE-MSC (F); asterisco: astrocitos que rellenan los espacios existentes por la falta de mielina con sus cuerpos celulares y expansiones. Estos astrocitos reactivos son células grandes, ocasionalmente multinucleadas, con abundantes filamentos intermedios distribuidos en el citoplasma. Los núcleos son grandes, ligeramente irregulares, con cromatina laxa, escala 5 µm (B), escala 20 µm (D) y escala 5 µm (F). G: densidad celular en el NO por µm2. Diferencias significativas entre todos los grupos (p < 0.05). H: densidad axonal (números de axones por µm2). Diferencias estadísticamente significativas a favor del grupo EAE-MSC (p < 0.05).
Estos datos se confirmaron en las imágenes a MET, los axones en el NO de los ratones inmunizados mostraron signos de degeneración, como un citosol oscuro con una red de microtúbulos interrumpida en el axonema y cuerpos de esferoides de mielina, en lugar de vainas mielínicas concéntricas regulares, como se observó en los ratones de control (Fig. 2 D), además se observaron abundantes astrocitos reactivos en el espacio intercelular entre los axones mielinizados dañados (asterisco en 2 D).
El tratamiento con aMSC disminuyó los signos degenerativos en la mielina y los axones, mostró menos infiltrados de células gliales en el NO dañado y dio lugar a un estado intermedio entre los ratones control y tratados. Para determinar estos cambios cuantitativamente, obtuvimos secciones transversales de NO y, al teñir con azul de toluidina, se contaron las células por unidad de área (µm2) (Fig. 2 G). En el grupo control, el promedio de células por unidad de área fue de 1.86 células/µm2, mientras que en el grupo de EAE, 7.99 células/µm2. El tratamiento con aMSC dio como resultado 3.69 células/µm2. Los estudios estadísticos mostraron una diferencia significativa entre los tres grupos (p < 0.0001). Posteriormente, las diferencias específicas entre la celularidad media de los sujetos en ambos grupos (EAE y EAE-MSC) se compararon mediante la prueba U de Mann Whitney, que también resultó estadísticamente significativa (p = 0.0143). Para calcular la densidad axonal (Fig. 2 H), contamos el número de axones por unidad de área. Mientras que en el grupo de control obtuvimos una densidad axonal de 0.38049 axones/µm2, en el grupo EAE, el valor obtenido fue de 0.16745 axones/µm2, y esta disminución fue significativa (p = 0.0003). La comparación de la densidad axonal entre EAE y el grupo tratado con administración aMSC (0.3598 axones/µm2) fue estadísticamente significativa a favor del grupo tratado (p=0.0251).
Los resultados del G-ratio en los ratones EAE-MSC muestran una desmielinización que se recuperó tras el tratamiento con aMSC (Fig. 3). En el grupo de control, el 79% de los axones estaban normalmente mielinizados, el 21% de los axones ingresaron en el grupo de los axones remielinizados y ninguno de los axones estudiados fue desmielinizado. En contraste, en el grupo de EAE, encontramos un 12% de los axones normales, un 73% de remielinización de los axones y un 15% de los axones con daño agudo de la mielina. Los ratones EAE-MSC exhibieron el 46% de los axones envueltos con mielina normal, el 47% en proceso de remielinización y el 7% de los axones desmielinizados. Cabe señalar que, si consideramos el porcentaje de axones que habían sufrido algún daño (agrupando los procesos de remielinización y desmielinización) con valores de G por encima de 0.84, el total de axones alterados fue del 88% en el grupo de EAE comparado con el 54% en el grupo EAE-MSC.
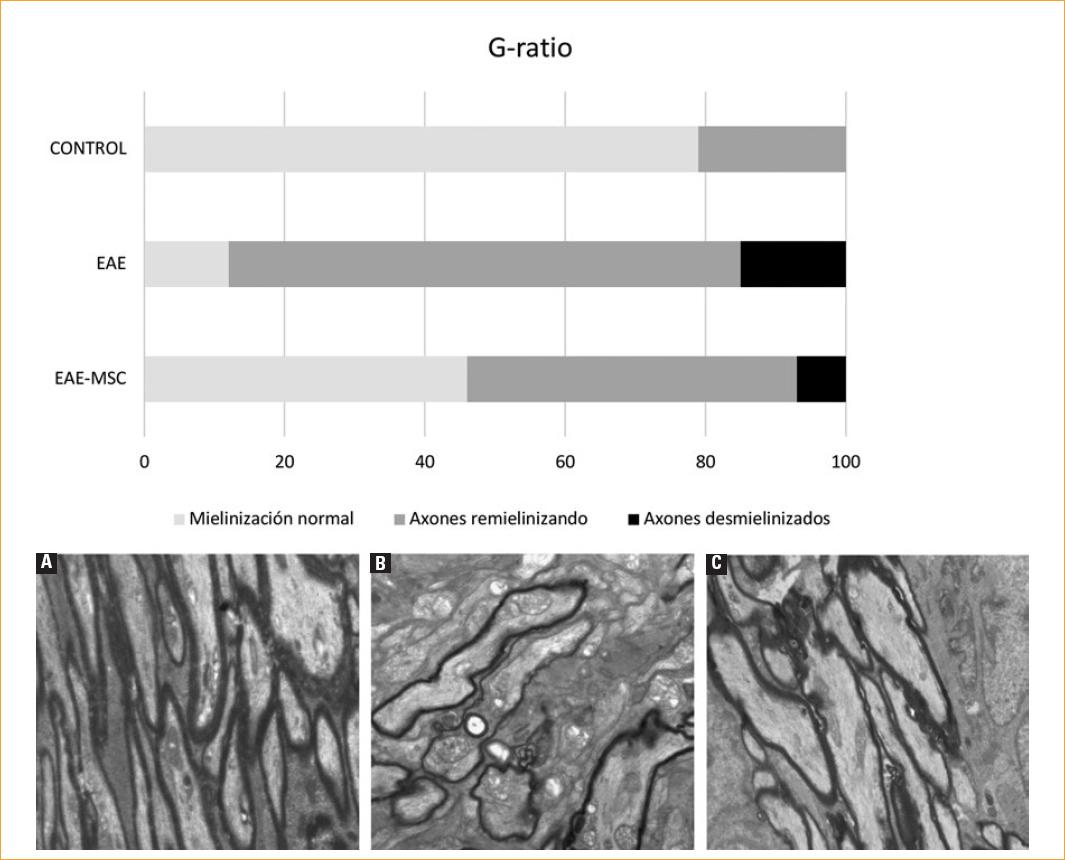
Figura 3 Estudio del grado de mielinización. G-ratio. Los ratones EAE-MSC muestran una desmielinización que se recuperó tras el tratamiento con aMSC. A-C: estudio ultraestructural axonal. A: control: axones con mielinizacion homogénea: mielinización normal. B: EAE: axones desorganizados con gran desmielinizacion. C: EAE-MSC: axones desorganizados en proceso de remielinización, escala 5 µm.
Discusión
En este trabajo hemos estudiado el posible efecto de las aMSC, hemos evaluado que la infusión intravenosa de aMSC podría mejorar el daño inflamatorio, desmielinizante y neurodegenerativo tanto en la retina como en el NO, mientras que, desafortunadamente, el efecto clínico no fue suficiente para detener la progresión de la enfermedad.
Por otro lado, estudios publicados sobre diferentes modelos de daño axonal (isquemia transitoria de la retina13 o axotomía NO14 describieron que la interrupción de los axones de las CGR causó degeneración axonal retrógrada que provocó la muerte de las CGR15. En nuestro trabajo observamos una pérdida significativa de CGR en ratones EAE en comparación con el grupo control, lo que podría ser el resultado del daño axonal documentado en el NO y la degeneración Walleriana. Otra posibilidad sería que, como la inflamación del NO y el daño estructural parecían ser más relevantes, la muerte de los CGR fuera provocada por el efecto tóxico local de la microglía, que aumentó en los grupos de EAE y EAE-MSC, como se informó en otros modelos de inflamación ocular16. El tratamiento con aMSC mejoró la inflamación en el NO, disminuyó la pérdida de CGR, pero no se observó ningún efecto en el rescate de las CGR ya perdidas, esto podría deberse a un mejor acceso de las aMSC al NO a través de la barrera hematoencefálica17 o a un efecto retardado de la disminución de la inflamación en el NO debido a aMSC en la degeneración de Walleriana en la retina.
Las aMSC representan una población de células madre multipotenciales que cuando se administran por vía intravenosa tienen la capacidad de migrar e injertarse en lugares donde se producen daños en los tejidos e inflamación. Describimos que las aMSC administradas por vía intravenosa tenían la capacidad de disminuir los infiltrados inflamatorios y reducir la pérdida de mielina y el daño axonal. Este efecto no se relacionó con el injerto de aMSC, porque no pudimos encontrar aMSC fluorescentes verdes en nuestras muestras. Esto podría deberse a varias causas, incluido el secuestro en otros órganos (hígado, pulmones), el rechazo inmunológico y/o la muerte de células por apoptosis una vez en el SNC. Según los estudios realizados a este respecto, las aMSC administradas por la vena de la cola en ratones inicialmente se acumulan en los pulmones y pasan gradualmente al hígado, bazo, riñón y médula ósea en las primeras 48 horas18,19. Rochefort, et al. encontró acumulación en pulmón del 50-60% de las aMSC 1 hora después de la inyección, con una disminución subsiguiente hasta aproximadamente el 30% 3 horas después de la inyección20. En humanos, el mismo grupo inyectó aMSC por vía intravenosa y también mostró una alta captación pulmonar en 30 minutos21. Numerosos estudios han demostrado la baja inmunogenicidad de aMSC que, al no expresar MHC-II, evita el reconocimiento por parte de las células dendríticas y los linfocitos T. Sin embargo, aunque se han considerado células que no producen rechazo cuando las células se exponen a un medio inflamatorio caracterizado por la presencia de niveles elevados de interferón-?22 comienzan a expresar el HLA-DR (MHC-II), y por lo tanto se convierten en células inmunogénicas, y son susceptibles de rechazo. Sin embargo, nuestros estudios y otros demuestran que, además de la baja eficiencia del injerto del SNC, hay un efecto neuroprotector después de la administración de aMSC. Este efecto, denominado «by-stander», se ha descrito en estudios en infarto de miocardio después de la infusión de aMSC, donde se observó una mejoría clínica y funcional, pero no fue posible identificar células injertadas o células diferenciadas de los miocardiocitos23,24. Las aMSC se caracterizan por una alta capacidad de comunicación paracrina basada en la liberación de pequeñas vesículas o exosomas25. A los exosomas se les ha atribuido el efecto de las aMSC a distancia. Podría ser que la disminución de la respuesta inflamatoria observada en nuestro estudio pudiera deberse a la liberación a distancia de exosomas con acción neurotrófica e inmunomoduladora adicional. Sin embargo, los mecanismos exactos todavía no se entienden completamente.
Dentro de las limitaciones de este estudio se encuentra el tamaño muestral reducido, y el seguimiento de la enfermedad con escalas de discapacidad motora por falta de medios por nuestra parte; el seguimiento con técnicas como la tomografía de coherencia óptica26 aplicada en animales, para visualización de estructuras retinianas y del NO, in vivo hubiera permitido hacer un mejor control de la enfermedad. Sería interesante en un futuro ampliar este estudio mejorando las limitaciones encontradas.
La aportación más interesante de este trabajo es su enfoque oftalmológico al estudiar la muerte de las CGR, así como las técnicas sofisticadas empleadas para ello como es la microscopía electrónica.
Finalmente, quisimos enfatizar que, tras la administración de aMSC, encontramos una menor pérdida de CGR, disminución de la pérdida de mielina y menor grado de inflamación en el NO; sin embargo, sería aconsejable ampliar la metodología para poder asegurar un efecto inmunosupresor y neuroprotector en este modelo. Dado que el daño tisular derivado de la inflamación es un proceso clave que se desencadena en respuesta a una lesión y enfermedad, las aMSC podrían convertirse en tratamiento de cualquier daño tisular u orgánico asociado con una actividad inflamatoria intensa.
Conclusiones
En este trabajo encontramos, tras la administración de aMSC, una menor pérdida de CGR, disminución de la pérdida de mielina y menor grado de inflamación en el NO; sin embargo, sería aconsejable ampliar la metodología para poder asegurar un efecto inmunosupresor y neuroprotector en este modelo.

