










Services on Demand
Journal
Article
 text in
text in  English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista mexicana de ciencias agrícolas
Print version ISSN 2007-0934
Rev. Mex. Cienc. Agríc vol.12 n.6 Texcoco Aug./Sep. 2021 Epub Mar 21, 2022
https://doi.org/10.29312/remexca.v12i6.3012
Articles
PVA-Chitosan-nCu complex improves yield and defense response in tomato
1Master of Science in Horticulture-Antonio Narro Autonomous Agrarian University. Saltillo, Mexico. CP. 25315. (athalia-rivera@hotmail.com).
2Center for Research in Applied Chemistry. Saltillo, Mexico. CP. 25294. (gregorio.cadenas@ciqa.edu.mx).
3Department of Horticulture-Antonio Narro Autonomous Agrarian University. (abenmen@gmail.com; asandovalr16@gmail.com; cafum7@yahoo.com).
4CONACYT-Antonio Narro Autonomous Agrarian University.
Currently the use of nanotechnology is revolutionizing agricultural production. Copper nanoparticles have been shown to influence the growth and development of different plant species, in addition to operating as stress resistance inducers. The objective of the present research was to evaluate the response in growth and yield, as well as the activation of the defense system of tomato plants. The treatments evaluated were a complex of polyvinyl alcohol-chitosan-copper nanoparticles (PVA-Cts-nCu), another complex of PVA-Cts and an absolute control (T0). The treatments were applied via foliar in tomato plants under greenhouse conditions. During the crop cycle, agronomic variables were determined, and the activity of enzymes related to stress tolerance such as β-1,3 glucanase, chitinase and phenylalanine ammonia lyase (PAL), as well as the expression of the PR1 gene. The PVA-Cts-nCu complex increased yield, number of fruits, average fruit weight, aerial fresh weight and root fresh weight, in addition, it promoted the defense system by increasing the PAL enzyme activity, as well as the overexpression of the PR1 gene.
Keywords biostimulant; gene expression; stress; vegetables
Actualmente el uso de la nanotecnología está revolucionando la producción agrícola. Se ha demostrado que las nanopartículas de cobre tienen un efecto en el crecimiento y desarrollo de las diferentes especies vegetales, además de operar como inductores de resistencia al estrés. El objetivo del presente trabajo fue evaluar la respuesta en el crecimiento y rendimiento, así como la activación del sistema de defensa de plantas de tomate. Los tratamientos evaluados fueron un complejo de alcohol polivinílico-quitosán-nanopartículas de cobre (PVA-Cts-nCu), otro complejo de PVA-Cts y un testigo absoluto (T0). Los tratamientos se aplicaron vía foliar en plantas de tomate bajo condiciones de invernadero. Durante el ciclo del cultivo, se determinaron variables agronómicas, y la actividad de enzimas relacionadas a la tolerancia a estrés como β-1,3 glucanasa, quitinasa y fenilalanina amonio liasa (PAL), así como la expresión del gen PR1. El complejo PVA-Cts-nCu incrementó el rendimiento, número de frutos, peso promedio de fruto, peso fresco aéreo y peso fresco de la raíz, además, promovió el sistema de defensa mediante el aumento en la actividad enzimática PAL, así como la sobreexpresión del gen PR1.
Palabras clave bioestimulante; estrés; expresión de genes; hortalizas
Introduction
The agricultural sector currently faces significant challenges of increasing productivity to feed the growing world population and increasing the efficiency in resource use (Rouphael and Colla, 2020). This has led to the expansion of the field of nanotechnology research and its possible applications in the agricultural field (Usman et al., 2020). Nanoparticles (NPs) have a highly reactive surface, therefore, they are biologically active, this because their surface area is extremely high proportional to their volume (Pestovsky and Martínez, 2017). They are synthesized in different sizes, shapes, materials and reactivity, this gives them the ability to enhance the agricultural sector (Fatima et al., 2021).
Copper (Cu) is a micronutrient widely distributed in plant tissues, participates in physiological processes and is essential for plant growth (Rajput et al., 2018). It has been shown to activate key enzymes in the Calvin cycle, improving photosynthetic activity and increasing yield in plants (Pradhan et al., 2015). Cu NPs can function as reducers or oxidants in biochemical reactions within the cell so they can catalyze the production of reactive oxygen species (ROS) and induce oxidative stress (Somasundaran et al., 2010). The effects of Cu NPs vary, it has been reported that, at low concentrations, it increases the growth rate and germination of several plants, on the other hand, at high concentrations growth is delayed (Kasana et al., 2017). In addition, in foliar applications with Cu NPs, they favor the firmness of fruits and content of antioxidants such as vitamin C, lycopene and phenols (López-Vargas et al., 2018; Pérez-Labrada et al., 2019).
Chitosan (Cts) has been used to improve the natural defenses of plants, defense responses include lignification, activation of enzymes associated with the response to pathogens in plants, phytoalexin biosynthesis, ROS generation, biosynthesis of jasmonic acid, salicylic acid, abscisic acid and the expression of genes related to defense (El Hadrami et al., 2010). The application of chitosan increases the activity of β-1,3 glucanase and chitinase in different plant species, helping to hydrolyze the cell wall of phytopathogenic fungi (Rodríguez-Pedroso et al., 2009; González Peña et al., 2014).
Polyvinyl alcohol (PVA) has been widely used in the preparation of hydrogels and in the inhibition of bacterial growth, in addition, it has a high potential as a controlled release system (Yang et al., 2016). The PVA-Cts mixture has been shown to have in vitro effects on the antioxidant response by removing free radicals, as well as on the viability of Staphylococcus aureus 8325-4 and Escherichia coli RB (Yang et al., 2018).
There are currently few studies on the application of PVA-Cts-nCu complexes. These complexes applied in the form of hydrogels increase the nutraceutical quality in tomato fruits (Hernández-Hernández et al., 2018a). They also promote the increase of vigor and number of floral clusters in tomato (Hernández-Hernández et al., 2017). Applied in substrate, they increase the yield in jalapeño chili fruits (Pinedo-Guerrero et al., 2017). The objective of the present study was to evaluate the response of the PVA-Cts-nCu complex in the growth and quality of fruit, as well as the activation of the defense system of tomato plants.
Materials and methods
Synthesis of the PVA-Cts and PVA-Cts-nCu complex
For the preparation of the PVA-Cts-nCu complex, 1.5 L of PVA (BP-05, Nacional PIM Mexico SA de CV, molecular weight of 27 000-32 000 g mol-1) at 1% (w/v) were mixed with 1.5 L of Cts (Marine Chemicals, India, molecular weight of 200 000 g mol-1) at 0.5% (w/v). The nCu (1.85 g) was ground with 3 ml of Agrex® F (Agroenzymas® SA de CV) as a dispersant for NPs. The nCu exhibited a spherical and hemispherical morphology and an average size of 30 nm (Sierra-Ávila et al., 2014, 2015).
Establishment of the crop
The test was carried out in a tunnel-type greenhouse. A soilless growing system was used, the substrate was a mixture of peat moss (Premier® Premier Horticulture LTD) and perlite (Multiperl® Lagoon Perlite Group) (1:1 v/v). Cid F1 saladette-type hybrid tomato plants (Harris Moran®) of indeterminate growth were used, they were placed in polyethylene bags of 14 L.
The experiment was carried out applying the treatments PVA-Cts-nCu 500 mg L-1, PVA-Cts 500 mg L-1 and an absolute control with distilled water (T0). The application of the treatments was foliar, with an approximate consumption of 75 ml of solution per plant throughout the production cycle. The seedlings received an application two days before the transplantation, subsequently it was applied at intervals of 20 days starting after the transplant. The Steiner solution (Steiner 1961) was used as a means of nutrition.
Variables of development and productivity in tomato
The evaluation of plant height and stem diameter was carried out 120 days after transplantation (DDT). At 120 DDT, the yield per plant was determined. In addition, the aerial fresh weight was determined, as well as the root fresh weight.
Enzyme extraction
The activity of the enzymes phenylalanine ammonia lyase (PAL), chitinase and β-1,3 glucanase was analyzed in leaves in three different samples, at the time of transplantation (48 h after the first application of the complex), at 40 DDT and 80 DDT. The leaves were collected with liquid nitrogen.
The extraction of biomolecules was performed according to Rodríguez-Pedroso et al. (2006). PAL activity was determined according to Sykłowska-Baranek et al. (2012). The results were expressed as U (production of μmol of trans-cinnamic acid equivalent per milliliter per minute) per total proteins (mg g-1). Chitinase activity was determined according to Rodríguez-Pedroso et al. (2006). The results were expressed as U (production of μg ml-1 of glucose per minute per total proteins (mg g-1). The β-1,3 glucanase activity was determined according to Rodríguez-Pedroso et al. (2006). Enzymatic activity was determined by measurement of the level of production of reducing sugars and was expressed in terms of production of μg ml-1 of glucose per minute per total proteins (mg g-1).
Expression of the PR1 gene
The leaf samples were collected at three different times, starting at 48 h after the first application of the complex, after this first sampling the following were every 40 days. Young leaves completely expanded were collected with liquid nitrogen. RNA extraction was carried out using the TRIzolTM Reagent (Invitrogen) technique.
The cDNA was synthesized with the SensiFASTTM (Bioline) kit. The primers used correspond to an endogenous gene (Actin) and the PR1 gene. ACT (fwd 5’-CCCAGGCACACAGGTGTTAT-3’; rev 5’-CAGGAGCAACTCGAAGCTCA-3’); PR1 (fwd 5’-AAGTAGTCTGGCGCAACTCA-3’; rev 5’-GTCCGATCCAGTTGCCTACA-3’). The quantification of the PR1 gene was carried out in real-time PCR equipment (Applied Biosystems StepOneTM version 2.3) by the ΔΔCt method, measuring the fluorescence intensity of Sybr Green (Hernández-Hernández et al., 2018b).
Data analysis
A completely random design was used in the test in tomato plants. For the agronomic variables, 20 experimental units were analyzed for each treatment. In the quantification of PAL, chitinase and β-1,3 glucanase activity, five repetitions per treatment were analyzed and in the expression of the PR1 gene, four composed repetitions were analyzed (10 plants per repetition). To detect statistical differences between treatments, an analysis of variance (Anova) was performed, and a mean separation test was carried out according to Fisher’s LSD test (p≤ 0.05). All statistical analyses were performed in the InfoStat v2018 statistical software.
Results and discussions
Effect of the PVA-Cts-nCu complex on developmental and productivity variables in tomato
The foliar application of the PVA-Cts-nCu complex had significant effects (p≤ 0.5) on the variables related to vigor in tomato plants (Table 1). The PVA-Cts-nCu treatment showed significant differences compared to the control, increasing the yield (60.68%), the average fruit weight (18.20%), the number of fruits (35.99%), the aerial fresh weight (26.99%) and the root fresh weight (80.87%). While the PVA-Cts treatment increased the average fruit weight (8.39%) and the root fresh weight (52.22%) compared to the control. No significant differences were observed in the variables of stem height and diameter.
Table 1 Effect of the PVA-Cts-nCu and PVA-Cts complex on tomato growth and productivity.
| Treatment | Height (cm) |
DT (mm) |
Yield (g plant-1) |
PPF (g fruit-1) |
NF | PFA (g plant-1) |
PFR (g plant-1) |
|---|---|---|---|---|---|---|---|
| T0 | 132.29 a | 17.02 a | 2573.31 b | 58.62 c | 43.9 b | 2384 b | 98.8 b |
| PVA-Cts-nCu | 134.78 a | 16.79 a | 4134.95 a | 69.29 a | 59.7 a | 3027.65 a | 175.8 a |
| PVA-Cts | 119.71 a | 15.04 a | 2690.92 b | 63.54 b | 42.35 c | 2299.47 b | 150.4 a |
T0= control; DT= stem diameter; NF= number of fruits; PPF= average fruit weight; PFA= aerial fresh weight; PFR= root fresh weight, means with the same letter within each column are statistically equal (Fisher’s LSD, p≤ 0.05).
The results obtained in this study demonstrate that the PVA-Cts-nCu and PVA-Cts complexes applied via foliar do not show toxicity in tomato plants. Rajput et al. (2018) mention that phytotoxicity will depend on the concentration, type and size of the NPs, in addition to the species in which they are applied, the growing conditions and the time of exposure.
Saharan et al. (2015) report an increase in biomass in tomato plants treated with Cu-Cts NPs. Adhikari et al. (2012) observed that the application of CuO NPs increased root growth in soybean and chickpea. In the present study, it was shown that the PVA-Cts-nCu complex improved growth by increasing the aerial and root fresh weight, very important parameters for the development of the crop. This effect might be induced by the synergy between nCu and chitosan. Since this biopolymer and nCu have shown to influence the growth and development of plants (El Hadrami et al., 2010; Hernández et al., 2017; Pinedo-Guerrero et al., 2017).
At the production stage, the PVA-Cts-nCu complex increased yield, number of fruits and average fruit weight. This coincides with what was reported by Pinedo-Guerrero et al. (2017), who point out that when applying the hydrogel of PVA-Cts-nCu direct to the soil in jalapeño chili, it increased the yield (8.27%) and the number of fruits (9.32%) per plant. Also, Hernández-Hernández et al. (2018a) reported that the application of hydrogels of PVA-Cts-nCu direct to the substrate increased the number of fruits in tomato plants (20%).
Pradhan et al. (2015) evaluated the application of Cu NPs in green soybean, where NPs influenced the activity of key enzymes in the Calvin cycle (fructose-1, 6-bisphosphate phosphatase, ribulose-5-phosphate kinase and NADP-glyceraldehyde-3-phosphate dehydrogenase). It is likely that when the activity of enzymes related to the transport of electrons in photosynthesis increases, the rate of production of photosynthates and the rate at which they are transported will be modified, thus increasing the production of fruits and their weight.
Effect of the PVA-Cts-nCu complex on PAL activity
Foliar application of PVA-Cts-nCu had significant effects (p≤ 0.5) on PAL activity in leaves (Figure 1). The PVA-Cts-nCu treatment exhibits an increase in PAL activity at transplantation (301.70%) and at 80 DDT (55.04%) compared to the control. At 40 DDT, the PVA-Cts-nCu treatment increased PAL activity by 369.23% compared to PVA-Cts.
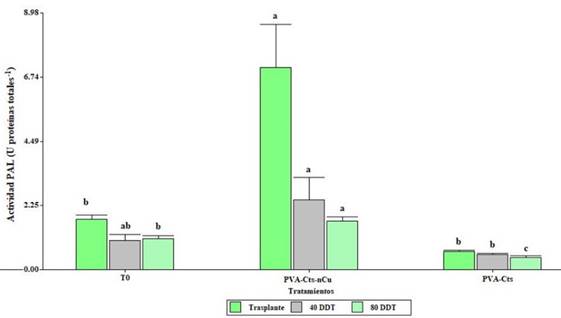
Figure 1 Enzymatic activity of phenylalanine ammonium lyase in tomato with application of PVA-Cts-nCu and PVA-Cts. Means with the same letter are statistically equal LSD Fisher, p≤ 0.05.
Chitosan and NPs are known as potential inducers to improve the defense response in plants to stress, resulting in an accumulation of metabolites related to defense (Zhang and Liu, 2015; Usman et al., 2020), as well as the generation of ROS and the increase in the activity of proteins related to defense (González-Peña et al., 2014).
The PAL enzyme is key in the synthesis of phenols and flavonoids, phenols are important precursors of compounds involved in the control of plant growth, in addition to being powerful antioxidants (Santos-Sánchez et al., 2019).
The increase in PAL can be derived from the direct interaction of nCu and Cts generating a synergistic effect. Cumplido-Nájera et al. (2019) applied Cu NPs foliarly increasing PAL activity in leaves by 1.78 times.
Falcón Rodríguez et al. (2012) reported that by spraying chitosan (0.5, 1 and 2.5 g L-1) in tobacco plants, all concentrations increased the activity of PAL in the leaf. Rodriguez et al. (2006) found that previously treating rice seeds with chitosan hydrolysates (500 mg L-1) stimulated the PAL activity in the leaf. In an essay by Hernández-Hernández et al. (2018a) with tomato plants under saline stress, it was found that the application of PVA-Cts-nCu without saline stress and PVA-Cts under saline stress increased the PAL activity in the leaf. González Peña et al. (2014) report that when spraying chitosan (100 mg L-1) in tomato seedlings, an increase in PAL activity was recorded. Chandra et al. (2015) foliarly applied Cts NPs in Camellia sinensis L., finding a significant increase in the accumulation of peroxidase, polyphenol oxidase and PAL in the leaf.
Effect of the PVA-Cts-nCu complex on the β-1, 3 glucanase and chitinase activity
The foliar application of both treatments had no significant effects (p≤ 0.5) with respect to the control, however, the PVA-Cts-nCu showed an increase in the β-1, 3 glucanase activity with respect to PVA-Cts (162.85%) at 40 DDT. At 80 DDT, T0 increased the activity above PVA-Cts-nCu and PVA-Cts (Figure 2).
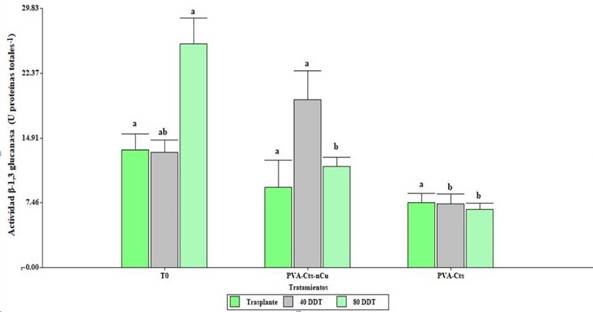
Figure 2 Enzymatic activity of β-1, 3 glucanase in tomato plants with the application of PVA-Cts-nCu and PVA-Cts. Means with the same letter are statistically equal Fisher’s LSD, p≤ 0.05.
Foliar application of PVA-Cts-nCu had significant effects (p≤ 0.5) on chitinase activity (Figure 3). At 40 DDT, the PVA-Cts-nCu treatment increased the activity by 121% compared to PVA-Cts. At 80 DDT, T0 increased the activity above PVA-Cts-nCu and PVA-Cts. The PVA-Cts-nCu treatment increased the activity by 54.67% compared to PVA-Cts.
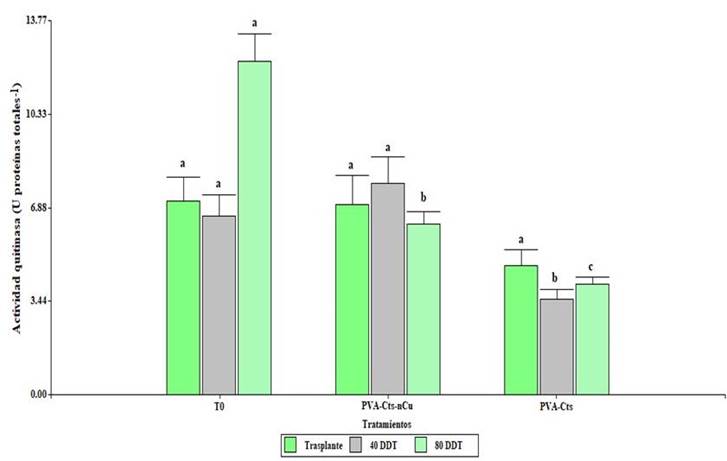
Figure 3 Enzymatic activity of chitinase in tomato plants with the application of PVA-Cts-nCu and PVA-Cts. Means with the same letter are statistically equal Fisher’s LSD, p≤ 0.05.
Proteins related to pathogenicity (PR) such as β-1,3 glucanase and chitinase can be expressed constitutively and induced in response to an infection. Regarding the effectiveness of chitosan in the induction of the defense in plant, in leaves of Camellia sinensis L., it increased the activity of β-1,3-glucanase 29.26%, as well as the content of peroxidase and polyphenol oxidase (Chandra et al., 2015). When applying NPs Cts and Cts in tomato plants, these increased the relative expression of genes related to the synthesis of chitinase and β-1,3 glucanase compared to the control (Chun and Chandrasekaran, 2019).
Effect of PVA-Cts-nCu on the expression of the PR1 gene
The results of the expression of the PR1 gene are shown in Figure 4. The PVA-Cts-nCu treatment overexpressed the PR1 gene (268.3, 0.72 and 8 times with respect to T0) at transplantation, 40 and 80 DDT. Like PVA-Cts-nCu, the PVA-Cts treatment overexpressed the PR1 gene (3.9, 1.5 and 123.2 times with respect to T0) at transplantation, 40 and 80 DDT. These results suggest that the PVA-Cts-nCu complex could play an important role in the activation of genes that encode the production of PR proteins, related to the salicylic acid signaling pathway and systemic acquired resistance (RSA) linked to stress tolerance (AbuQamar et al., 2009).
There is evidence that chitosan can activate the defense response in plants, increasing the activity of PAL enzyme that is directly related to the synthesis of salicylic acid, thus inducing the synthesis of PR proteins, involved in the mechanisms of RSA (Rodríguez-Pedroso et al., 2006; Sánchez et al., 2010; González Peña et al., 2014).
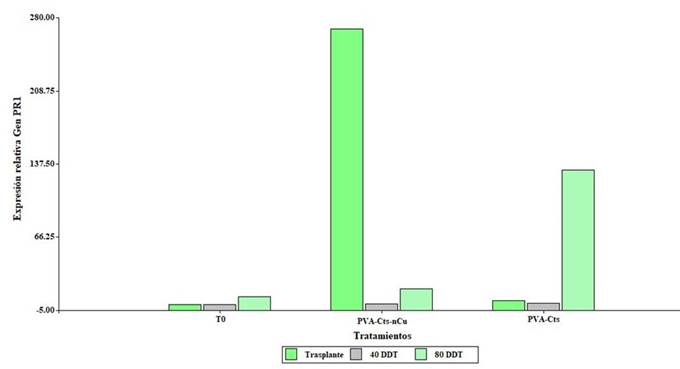
Figure 4 Relative expression of the PR1 gene in plants tomato leaves treated with PVA-Cts-nCu and PVA-Cts.
Chun and Chandrasekaran (2019) applied Cts NPs foliarly in tomato plants inoculated with Fusarium andiyazi, these increased the relative expression of the PR1 gene in the leaf compared to the control. Hernández-Hernández et al. (2018b) applied hydrogels of PVA-Cts-nCu direct to the substrate in tomato under conditions of saline stress and found that the expression of the PR1 gene was repressed under saline condition and without saline stress, however, the JA gene related to the jasmonate pathway was overexpressed and it is mediated by induced systemic resistance (RSI).
These results show that the PVA-Cts-nCu and PVA-Cts complexes may be potential inducers to mediate the expression of PR1 and JA genes in tomato plants activating the signaling cascade of synthesis of phenolic compounds and enzymes that will prepare the plant for the effects of stress, either by the salicylic acid pathway (RSA) or the jasmonic acid pathway (RSI).
Conclusions
This study showed that polyvinyl alcohol-chitosan-nanocopper and polyvinyl alcohol-chitosan increased the variables related to vigor in plants, yield, in addition to activating the defense mechanism of plants by increasing enzymes such as PAL, β-1, 3 glucanase and chitinase, in addition to increasing the expression of the PR1 gene linked to stress tolerance in plants.
Acknowledgements
This work was carried out thanks to the National Council of Science and Technology (CONACYT) grant 887137. Master’s degree in Horticulture-Universidad Autónoma Agraria Antonio Narro. Saltillo, Mexico. G. C. P. for the contribution of reagents and materials. Applied Chemistry Research Center. Saltillo, Mexico.
REFERENCES
AbuQamar, S.; Luo, H.; Laluk, K.; Mickelbart, M. V. and Mengiste, T. 2009. Crosstalk between biotic and abiotic stress responses in tomato is mediated by the AIM1 transcription factor. The Plant J .58(2):347-360. [ Links ]
Adhikari, T.; Kundu, S.; Biswas, A. K.; Tarafdar, J. C. and Rao, A. S. 2012. Effect of copper oxide nano particle on seed germination of selected crops. J. Agric. Sci. Technol. 2(6A):815-823. [ Links ]
Chandra, S.; Chakraborty, N.; Dasgupta, A.; Sarkar, J.; Panda, K. and Acharya, K. 2015. Chitosan nanoparticles: a positive modulator of innate immune responses in plants. Scientific reports. 5(1):1-14. [ Links ]
Chun, S. C. and Chandrasekaran, M. 2019. Chitosan and chitosan nanoparticles induced expression of pathogenesis-related proteins genes enhances biotic stress tolerance in tomato. Inter. J. Biol. Macromol. 125(51):948-954. [ Links ]
Cumplido-Nájera, C. F.; González-Morales, S.; Ortega-Ortíz, H.; Cadenas-Pliego, G.; Benavides-Mendoza, A. and Juárez-Maldonado, A. 2019. The application of copper nanoparticles and potassium silicate stimulate the tolerance to Clavibacter michiganensis in tomato plants. Sci. Hortic. 245(1):82-89. [ Links ]
El Hadrami, A.; Adam, L. R.; El Hadrami, I. and Daayf, F. 2010. Chitosan in plant protection. Marine drugs. 8(4):968-987. [ Links ]
Falcón-Rodríguez, A.; Costales-Menéndez, D.; Martínez-Téllez, M. Á. y Gordon, T. A. 2012. Respuesta enzimática y de crecimiento en una variedad comercial de tabaco (Nicotiana tabacum, L.) tratada por aspersión foliar de un polímero de quitosana. Cultivos Tropicales. 33(1):65-70. [ Links ]
Fatima, F.; Hashim, A. and Anees, S. 2021. Efficacy of nanoparticles as nanofertilizer production: a review. Environ. Sci. Pollut. Res. 28(2):1292-1303. [ Links ]
Fish, W. W.; Perkins-Veazie, P. and Collins, J. K. 2002. A quantitative assay for lycopene that utilizes reduced volumes of organic solvents. J. Food Composition and Analysis. 15(3):309-317. [ Links ]
González-Peña, D.; Costales, D. y Falcón, A. B. 2014. Influencia de un polímero de quitosana en el crecimiento y la actividad de enzimas defensivas en tomate (Solanum lycopersicum L.). Cultivos Tropicales . 35(1):35-42. [ Links ]
Hernández, H H.; Benavides-Mendoza, A.; Ortega-Ortiz, H.; Hernández-Fuentes, A. D. and Juárez-Maldonado, A. 2017. Cu Nanoparticles in chitosan-PVA hydrogels as promoters of growth, productivity and fruit quality in tomato. Emirates J. Food Agric. 29(8):573-580. [ Links ]
Hernández-Hernández, H.; González-Morales, S.; Benavides-Mendoza, A.; Ortega-Ortiz, H.; Cadenas-Pliego, G. and Juárez-Maldonado, A. 2018a. Effects of chitosan-PVA and Cu nanoparticles on the growth and antioxidant capacity of tomato under saline stress. Molecules. 23(1):178-192. [ Links ]
Hernández-Hernández, H.; Juárez-Maldonado, A.; Benavides-Mendoza, A.; Ortega-Ortiz, H.; Cadenas-Pliego, G.; Sánchez-Aspeytia, D. and González-Morales, S. 2018b. Chitosan-PVA and copper nanoparticles improve growth and overexpress the SOD and JA genes in tomato plants under salt stress. Agronomy. 8(9):175-185. [ Links ]
Kasana, R. C.; Panwar, N. R.; Kaul, R. K. and Kumar, P. 2017. Biosynthesis and effects of copper nanoparticles on plants. Environ. Chem. Letters. 15(2):233-240. [ Links ]
López-Vargas, E. R.; Ortega-Ortíz, H.; Cadenas-Pliego, G.; de Alba Romenus, K.; Cabrera de la Fuente, M.; Benavides-Mendoza, A. and Juárez-Maldonado, A. 2018. Foliar application of copper nanoparticles increases the fruit quality and the content of bioactive compounds in tomatoes. Appl. Sci. 8(7):1020-1034. [ Links ]
Padayatt, S. J.; Daruwala, R.; Wang, Y.; Eck, P. K.; Song, J.; Koh, W. S. and Levine, M. 2001. Vitamin C: from molecular actions to optimum intake. Handbook of antioxidants. CRC Press. Washington, DC. USA. 117-145. [ Links ]
Pérez-Labrada, F.; López-Vargas, E. R.; Ortega-Ortiz, H.; Cadenas-Pliego, G.; Benavides-Mendoza, A. and Juárez-Maldonado, A. 2019. Responses of tomato plants under saline stress to foliar application of copper nanoparticles. Plants. 8(6):151-167. [ Links ]
Pestovsky, Y. S. and Martínez-Antonio, A. 2017. The use of nanoparticles and nanoformulations in agriculture. J. Nanoscie. Nanotechnol. 17(12):8699-8730. [ Links ]
Pinedo-Guerrero, Z. H.; Hernández-Fuentes, A. D.; Ortega-Ortiz, H.; Benavides-Mendoza, A. and Cadenas-Pliego, G. 2017. Cu nanoparticles in hydrogels of chitosan-PVA affects the characteristics of post-harvest and bioactive compounds of jalapeño pepper. Molecules. 22(6):926. [ Links ]
Pradhan, S.; Patra, P.; Mitra, S.; Dey, K. K.; Basu, S.; Chandra, S.; and Goswami, A. 2015. Copper nanoparticle (CuNP) nanochain arrays with a reduced toxicity response: a biophysical and biochemical outlook on vigna radiata. J. Agric. Food Chem. 63(10):2606-2617. [ Links ]
Rajput, V. D.; Minkina, T.; Suskova, S.; Mandzhieva, S.; Tsitsuashvili, V.; Chapligin, V. and Fedorenko, A. 2018. Effects of copper nanoparticles (CuO NPs) on crop plants: a mini review. Bionanoscience. 8(1):36-42. [ Links ]
Ramos, S. J.; Faquin, V.; Guilherme, L. R. G.; Castro, E. M.; Ávila, F. W.; Carvalho, G. S. and Oliveira, C. 2010. Selenium biofortification and antioxidant activity in lettuce plants fed with selenate and selenite. Plant Soil Environ. 56(12):584-588. [ Links ]
Rodríguez, A. T.; Ramírez, M. A.; Falcón, A.; Utria, E. y Bautista, S. 2006. Estimulacion de algunas enzimas en plantas de arroz (Oryza sativa, L.) tratadas con un hidrolizado de quitosana. Cultivos Tropicales . 27(2):87-91. [ Links ]
Rodríguez-Pedroso, A. T.; Ramírez-Arrebato, M. Á.; Cárdenas-Travieso, R. M.; Falcón-Rodríguez, A. y Bautista-Baños, S. 2006. Efecto de la quitosana en la inducción de la actividad de enzimas relacionadas con la defensa y protección de plántulas de arroz (Oryza sativa L.) contra Pyricularia grisea Sacc. Rev. Mex. Fitopatol. 24(1):1-7. [ Links ]
Rodríguez-Pedroso, A. T.; Ramírez-Arrebato, M. A.; Rivero-González, D.; Bosquez-Molina, E.; Barrera-Necha, L. L. y Bautista-Baños, S. 2009. Propiedades químico-estructurales y actividad biológica de la quitosana en microorganismos fitopatógenos. Rev. Chapingo. Ser. Hortic. 15(3):307-317. [ Links ]
Rouphael, Y. and Colla, G. 2020. Biostimulants in agriculture. Frontiers in Plant Sci. 11(1):40-47. [ Links ]
Saharan, V.; Sharma, G.; Yadav, M.; Choudhary, M. K.; Sharma, S. S.; Pal, A. and Biswas, P. 2015. Synthesis and in vitro antifungal efficacy of Cu-chitosan nanoparticles against pathogenic fungi of tomato. Inter. J. Biol. Macromol . 75(1):346-353. [ Links ]
Sánchez, G. R.; Mercado, E. C.; Peña, E. B.; Cruz, H. R. y Pineda, E. G. 2010. El ácido salicílico y su participación en la resistencia a patógenos en plantas. Biológicas. 12(2):90-95. [ Links ]
Santos-Sánchez, N. F.; Salas-Coronado, R.; Villanueva-Cañongo, C. and Hernández-Carlos, B. 2019. Antioxidant compounds and their antioxidant mechanism. London, UK. IntechOpen. 1-28 p. [ Links ]
Sierra-Ávila, R. M. P.; Cadenas-Pliego, G.; Avila-Orta, C.; Betancourt, R.; Jiménez-Regalado, E.; Jiménez-Barrera, R. and Martínez, G. 2014. Synthesis of copper nanoparticles coated with nitrogen ligands. J. Nanomater. 2014(1):74-82. [ Links ]
Sierra-Ávila, R.; Pérez-Alvarez, M.; Cadenas-Pliego, G.; Comparán Padilla, V.; Ávila-Orta, C.; Pérez Camacho, O.; Jiménez-Regalado, E.; Hernández-Hernández, E. and Jiménez-Barrera, R. M. 2015. Synthesis of copper nanoparticles using mixture of allylamine and polyallylamine. J. Nanomaterials. 361797(1):1-8. [ Links ]
Somasundaran, P.; Fang, X.; Ponnurangam, S. and Li, B. 2010. Nanoparticles: Characteristics, mechanisms and modulation of biotoxicity. KONA powder and particle journal. 28(1):38-49. [ Links ]
Steiner, A. A. 1961. A universal method for preparing nutrient solutions of a certain desired composition. Plant Soil. 15(2):134-154. [ Links ]
Sykłowska-Baranek, K.; Pietrosiuk, A.; Naliwajski, M. R.; Kawiak, A.; Jeziorek, M.; Wyderska, S. and Chinou, I. 2012. Effect of l-phenylalanine on PAL activity and production of naphthoquinone pigments in suspension cultures of Arnebia euchroma (Royle) Johnst. In vitro Cellular & Developmental Biology-Plant. 48(5):555-564. [ Links ]
Usman, M.; Farooq, M.; Wakeel, A.; Nawaz, A.; Cheema, S. A.; ur Rehman, H. and Sanaullah, M. 2020. Nanotechnology in agriculture: Current status, challenges and future opportunities. Sci. Total Environ. 721(1):137778. [ Links ]
Yang, W.; Fortunati, E.; Bertoglio, F.; Owczarek, J. S.; Bruni, G.; Kozanecki, M. and Puglia, D. 2018. Polyvinyl alcohol/chitosan hydrogels with enhanced antioxidant and antibacterial properties induced by lignin nanoparticles. Carbohydrate Polymers. 181(1):275-284. [ Links ]
Yang, W.; Owczarek, J. S.; Fortunati, E.; Kozanecki, M.; Mazzaglia, A.; Balestra, G. M. adn Puglia, D. 2016. Antioxidant and antibacterial lignin nanoparticles in polyvinyl alcohol/chitosan films for active packaging. Industrial Crops and Products. 94(1):800-811. [ Links ]
Yu, Z. and Dahlgren, R. A. 2000. Evaluation of methods for measuring polyphenols in conifer foliage. J. Chem. Ecol. 26(9):2119-2140. [ Links ]
Zhang, X. and Liu, C. J. 2015. Multifaceted regulations of gateway enzyme phenylalanine ammonia-lyase in the biosynthesis of phenylpropanoids. Mol. Plant. 8(1):17-27. [ Links ]
Received: June 2021; Accepted: August 2021
 Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
