











 text new page (beta)
text new page (beta) Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkIntroducción
Desde su descripción en 1888, Chirodines Bates ha llamado la atención de diversos autores debido a su semejanza morfológica con Chlaenobia Blanchard, 1850 y Phytalus Erichson, 1856 (Bates, 1888, Morón, 1991). Morón (1991) menciona que las especies de Chirodines podrían representar una derivación antigua del tronco ancestral de las especies incluidas en Chlaenobia «grupo vexata» pues, a su parecer, comparte con ellas el mayor número de caracteres morfológicos; además, señala que en el linaje de Melolonthini, la estructura sencilla de las uñas tarsales puede considerarse como un estado primitivo, del cual probablemente se derivaron, por un lado, las uñas hendidas y bífidas, y por otro, las uñas dentadas, serradas y pectinadas. El subgénero Chirodines agrupa 4 especies de Phyllophaga Harris, 1827 (Morón y Solís, 1994), cuyos machos adultos se distinguen por presentar el cuerpo esbelto; vestidura dorsal ausente; frente sin carina occipital transversal; antenas con 10 artejos; patas alargadas, tarsos robustos, cobertura setosa ventral escasa; uñas masculinas heteromorfas, las pro- y mesotarsales simples, sin proyecciones en su margen inferior o con un pequeño proceso dentiforme y con los ápices enteros y aguzados, y las uñas metatarsales profundamente bífidas. Las especies de Chirodines han sido recolectadas en México (Oaxaca y Chiapas), Guatemala y Costa Rica, dentro de ambientes situados entre los 200 y 1,500 m de altitud, con vegetación de bosque tropical caducifolio y subcaducifolio (Bates, 1888, Morón, 1991, Morón y Solís, 1994). Por su parte, siguiendo los criterios de Saylor (1942) y Morón (1986), Chlaenobia ha sido considerado un subgénero que incluye cerca de 20 especies de Phyllophaga (Morón, 2003, Morón, 2006). Los adultos se caracterizan por tener el cuerpo alargado y esbelto, de color amarillo pajizo; regiones dorsales glabras; frente sin carina occipital transversal; antenas con 9 o 10 artejos; tarsos usualmente muy largos, con cobertura setosa abundante en su parte ventral; uñas pro-, meso- y metatarsales hendidas en ambos sexos. Presentan dimorfismo sexual muy acentuado en antenas, abdomen, pigidio y patas. Se distribuyen entre el sureste de los Estados Unidos y Panamá, principalmente en ambientes situados por debajo de los 1,500 m de altitud, con climas cálidos o semicálidos (Aragón y Morón, 2003, Cano y Morón, 2002, Chapin, 1935, Morón, 1986, Morón, 1988, Morón, 1992, Morón y Robbins, 2004).
En 1850, Blanchard definió Chlaenobia (originalmente monoespecífico), basándose en Chlaenobia ciliatipes; sin embargo, no menciona su procedencia (Blanchard, 1850). Más tarde, Lacordaire (1856) consideró el género como válido, monoespecífico y procedente de Brasil, pero lo incluyó en la «subtribu iv. Macrodactylides, grupo iv. Philochlenides», debido a una falsa apreciación de la disposición de las procoxas, ya que las considera «cónicas y prominentes» en lugar de «transversales». El género también es considerado válido por Gemminger y Harold (1869), quienes lo vincularon con géneros macrodactiliformes, dentro de «Melolonthini», indicando «Brasilia» como localidad típica (Morón, 1986). Más adelante, en la Biologia Centrali-Americana, Bates (1888) revisó una serie de 22 especímenes, colocó el género encabezando la subfamilia «Rhizotroginae», presentó una diagnosis clara, describió 6 especies centroamericanas: Chlaenobia scabripyga, C. latipes, C. tumulosa, C. aequata, C. rodriguezi y C. unituberculata (considerada en el presente estudio como sinónimo de C. vexata Horn, 1885) y discutió sus características morfológicas. Posteriormente, Arrow (1933) y Chapin (1935) describieron 5 nuevas especies y una subespecie procedentes de México y Panamá, de las cuales 3 cambiaron de nombre por incurrir en homonimia (C. colimana Arrow, 1933 por C. peccata Blackwelder, 1944; C. arrowi Chapin, 1935 por C. instabilis Blackwelder, 1944 y C. dissimilis Chapin, 1935 por C. pilositarsis Blackwelder, 1944), 2 conservaron los nombres originales (C. panamana Chapin, 1935 y C. personata Chapin, 1935) y C. chiapensis Chapin, 1935 cambió de subespecie a especie.Chapin (1935) hizo notar la cercanía entre Chlaenobia, Chirodines y Phytalus.
Después de revisar las especies norteamericanas de Phytalus y Listrochelus Blanchard, 1850, Saylor, 1939, Saylor, 1940, Saylor, 1942 consideró necesario reducir Phytalus, Listrochelus, Chlaenobia y Chirodines al rango subgenérico dentro de Phyllophaga, pues expresó que no observó suficientes caracteres de valor con los cuales se pudieran apoyar su posición como géneros. Esta idea fue aceptada y promovida por Arnett (1973) y Morón, 1982, Morón, 1986. Posteriormente, Morón junto con otros autores, describieron 13 especies de Phyllophaga (Chlaenobia) de México, Guatemala, El Salvador y Honduras (Aragón y Morón, 2003, Cano y Morón, 2002, Morón, 1988, Morón, 1992, Morón y Robbins, 2004), de las cuales C. sandersoniana Morón, 1992, C. reyesiana Morón, 1992 y C. schusteriana Cano y Morón, 2002 fueron transferidas a Phyllophaga (Phytalus) (Morón, 2006).
Bates erigió Chirodines en 1888 basándose en 2 machos de Chirodines zunilensis, colectados en el cerro Zunil, Guatemala. Este género, monobásico en un principio, mantuvo su categoría taxonómica durante 54 años, hasta que Saylor (1940) lo ubicó a nivel de subgénero dentro de Phyllophaga. Posteriormente, Morón (1991) describió a Chirodines ome apoyándose en 2 machos colectados en Chiapas y a Chirodines yei basándose en 3 machos colectados en Oaxaca. Morón y Solís (1994) describieron a Chirodines nahui, utilizando 4 machos colectados en Guanacaste, Costa Rica. Cabe resaltar que, hasta el día de hoy, las hembras de las 4 especies de Chirodines son desconocidas para la ciencia.
Siguiendo los estudios filogenéticos recientes de Rivera-Gasperín y Morón, 2013, Rivera-Gasperín y Morón, 2017, en los cuales se utilizaron muestras representativas de la mayoría de los grupos de especies y subgéneros de Phyllophaga (s.l.), se dedujo que la reclasificación de todos sus subgéneros es necesaria. Los resultados de dichos análisis cladísticos mostraron evidencias para restituir Chlaenobia a nivel de género y considerar a Chirodines como su sinónimo. El objetivo del presente estudio fue utilizar caracteres morfológicos externos de adultos y la anatomía de los genitales masculinos y femeninos, para confirmar la validez del género Chlaenobia y evidenciar que Chirodines es su grupo hermano dentro del mismo clado.
Materiales y métodos
Se seleccionaron los taxones siguiendo la clasificación propuesta por Morón (1986). Se revisaron 364 adultos de 153 especies, depositados en las siguientes colecciones: Colección Entomológica del Instituto de Ecología A. C., Xalapa, Veracruz (IEXA); Museum für Naturkunde der Humboldt Universität, Berlín (ZMHU); Natural History Museum, Londres (MNHL) y Colección Miguel Ángel Morón, Xalapa (MXAL). Del número total de especies estudiadas, 10 pertenecen a Chlaenobia, 2 a Chirodines, 88 son de otros grupos de Phyllophaga y 55 conforman el grupo externo (Tabla 1).
Tabla 1 Lista de las especies usadas en el análisis cladístico.
| Especie | Autor | País | Colección | |
|---|---|---|---|---|
| 1 | Cyclocephala lunulata | Burmeister, 1847 | MX | IEXA |
| 2 | Serica porcula | Casey, 1884 | EE. UU. | MXAL |
| 3 | Hoplia squamifera | Burmeister, 1844 | MX | MXAL |
| 4 | Macrodactylus nigripes | Bates, 1887 | MX | IEXA |
| 5 | Isonychus piperitus | Bates, 1887 | MX | MXAL |
| 6 | Diplotaxis hirsuta | Vaurie, 1958 | MX | MXAL |
| 7 | Melolontha melolontha | (Linnaeus, 1758) | IT | MXAL |
| 8 | Polyphylla petiti | Guerin, 1830 | MX | MXAL |
| 9 | Cryptotrogus weisei | Kraatz, 1888 | JO | MXAL |
| 10 | Anoxia orientalis | (Krynicky, 1832) | BG | MXAL |
| 11 | Rhizotrogus aestivus | (Olivier, 1789) | RU | MXAL |
| 12 | Amphimallon solstitialis | (Linnaeus, 1758) | CZ | MXAL |
| 13 | Miltotrogus vernus | (Germar, 1824) | HU | MXAL |
| 14 | Haplidia transversa | (Fabricius, 1801) | YU | MXAL |
| 15 | Schizonycha mimocontinens | Pope, 1960 | ZA | MXAL |
| 16 | Schizonycha propinqua | (Kolbe, 1891) | TZ | ZMHU |
| 17 | Heptophylla picea | Motschulsky, 1857 | JP | MXAL |
| 18 | Triodonyx gigantissima | (Saylor, 1935) | MX | MXAL |
| 19 | Triodonyx woodruffi | (Warner y Morón, 1992) | MX | MXAL |
| 20 | Triodonyx bellamyi | (Warner y Morón, 1992) | EE. UU | MXAL |
| 21 | Triodonyx lalanza | (Saylor, 1941) | MX | IEXA |
| 22 | Triodonyx nogueirai | (Warner y Morón, 1992) | MX | MXAL |
| 23 | Holotrichia serrata | (Fabricius, 1787) | IN | MXAL |
| 24 | Holotrichia rufoflava | Brenske, 1894 | IN | MXAL |
| 25 | Holotrichia nigra | (Redtenbacher, 1868) | CN | MXAL |
| 26 | Holotrichia lata | Brenske, 1892 | TW | MXAL |
| 27 | Holotrichia angolensis | Decelle, 1977 | AO | MNHL |
| 28 | Sebakwe coriacea | (Péringuey, 1904) | ZW | MNHL |
| 29 | Octoplasia basilanensis | Itoh, 1996 | PH | ZMHU |
| 30 | Octoplasia pseudogigantea | Itoh, 1996 | MY | ZMHU |
| 31 | Microtrichia kawadai | Nomura, 1959 | JP | MXAL |
| 32 | Microtrichia densicollis | Moser, 1913 | IN | MNHL |
| 33 | Latipalpus occidentalis | Itoh, 1998 | ID | ZMHU |
| 34 | Latipalpus fujiokai | Itoh, 1999 | MY | ZMHU |
| 35 | Catrachia soctona | Morón, 2000 | MX | MXAL |
| 36 | Catrachia sp. | / | MX | MXAL |
| 37 | Trichesthes tristis | (Fabricius, 1781) | EE. UU. | MXAL |
| 38 | Trichesthes lenis | (Horn, 1887) | MX | IEXA |
| 39 | Trichesthes maximus | (Bates, 1888) | MX | MNHL |
| 40 | Listrochelus laportaei | Blanchard, 1850 | MX | MNHL |
| 41 | Listrochelus huachuca | (Saylor, 1940) | EE. UU. | MNHL |
| 42 | Listrochelus praesidii | Bates, 1888 | MX | MNHL |
| 43 | Listrochelus timidus | Horn, 1878 | MX | MXAL |
| 44 | Listrochelus nogueiranus | (Morón, 2002) | MX | IEXA |
| 45 | Listrochelus scoparius | LeConte, 1856 | EE. UU. | MXAL |
| 46 | Listrochelus cristagalli | Arrow, 1933 | MX | MNHL |
| 47 | Listrochelus mucoreus | LeConte, 1856 | MX | MXAL |
| 48 | Listrochelus arenicolus | (Howden, 1960) | EE. UU. | MXAL |
| 49 | Listrochelus granti | (Saylor, 1940) | EE. UU. | MNHL |
| 50 | Listrochelus cavatus | Bates, 1888 | MX | MNHL |
| 51 | Listrochelus cochisa | (Saylor, 1940) | MX | MXAL |
| 52 | Listrochelus densicollis | LeConte, 1863 | MX | MXAL |
| 53 | Listrochelus miraflora | (Saylor, 1940) | MX | MXAL |
| 54 | Chlaenobia ciliatipes | Blanchard, 1850 | MX | MNHL |
| 55 | Chlaenobia vexata | Horn, 1885 | MX | MNHL |
| 56 | Chlaenobia latipes | Bates, 1888 | MX | MNHL |
| 57 | Chlaenobia peccata | (Blackwelder), 1944 | MX | IEXA |
| 58 | Chlaenobia scabripyga | Bates, 1888 | MX | MNHL |
| 59 | Chlaenobia aequata | Bates, 1888 | NI | MNHL |
| 60 | Chlaenobia chiapensis | Chapin, 1935 | MX | MNHL |
| 61 | Chlaenobia halffteriana | (Morón, 1992) | MX | MXAL |
| 62 | Chlaenobia instabilis | (Blackwelder, 1944) | MX | MXAL |
| 63 | Chlaenobia sp. nov. | / | MX | MXAL |
| 64 | Chlaenobia zunilensis | (Bates, 1888) | HN | MXAL |
| 65 | Chlaenobia nahui | (Morón y Solís, 1994) | CR | MXAL |
| 66 | P. (Phytalus) puberea | (Mannerheim, 1829) | BR | MXAL |
| 67 | P. (Phytalus) pruinosa | (Blanchard, 1850) | MX | MXAL |
| 68 | P. (Phytalus) trichodes | (Bates, 1888) | MX | MNHL |
| 69 | P. (Phytalus) senicula | (Bates, 1888) | MX | MXAL |
| 70 | P. (Phytalus) balli | Morón y Woodruff, 2008 | MX | MXAL |
| 71 | P. (Phytalus) obsoleta | (Blanchard, 1850) | MX | IEXA |
| 72 | P. (Phytalus) cometes | (Bates, 1888) | MX | MXAL |
| 73 | P. (Phytalus) coahuayana | Morón, 2006 | MX | MXAL |
| 74 | P. (Phytalus) sonora | Saylor, 1939 | MX | MXAL |
| 75 | P. (Phytalus) pentaphylla | (Bates, 1888) | MX | MXAL |
| 76 | P. (Phytalus) bilobatata | Saylor, 1939 | EE. UU. | MXAL |
| 77 | P. (Phytalus) macrocera | (Bates, 1888) | MX | MNHL |
| 78 | P. (Phytalus) oaxaca | Saylor, 1940 | MX | MXAL |
| 79 | P. (Phytalus) fissilabris | (Bates, 1888) | MX | MXAL |
| 80 | P. (Phytalus) lulaana | Morón, 2000 | MX | MXAL |
| 81 | P. (Phytalus) lineata | (Bates, 1888) | MX | MNHL |
| 82 | P. (Phytalus) hirtifrons | (Moser, 1921) | MX | ZMHU |
| 83 | P. (Phytalus) heteroclita | (Burmeister, 1855) | BR | MXAL |
| 84 | P. (Phytalus) luridipennis | (Moser, 1918) | EC | MXAL |
| 85 | P. (Phytalus) nitididorsis | Frey, 1975 | BR | MNHL |
| 86 | P. (Phyllophaga) hirticula | Knoch, 1801 | EE. UU. | MXAL |
| 87 | P. (Phyllophaga) profunda | (Blanchard, 1850) | EE. UU. | MXAL |
| 88 | P. (Phyllophaga) fraterna | Harris, 1842 | EE. UU. | MXAL |
| 89 | P. (Phyllophaga) crassissima | (Blanchard, 1850) | EE. UU. | MXAL |
| 90 | P. (Phyllophaga) ravida | (Blanchard, 1850) | MX | ZMHU |
| 91 | P. (Phyllophaga) dentex | (Bates, 1888) | MX | MNHL |
| 92 | P. (Phyllophaga) dasypoda | (Bates, 1888) | MX | MNHL |
| 93 | P. (Phyllophaga) fulviventris | (Moser, 1918) | MX | ZMHU |
| 94 | P. (Phyllophaga) pastassana | (Moser, 1924) | EC | ZMHU |
| 95 | P. (Phyllophaga) nuda | (Moser, 1918) | GY | ZMHU |
| 96 | P. (Phyllophaga) heteronycha | (Bates, 1888) | MX | MNHL |
| 97 | P. (Phyllophaga) hoegella | Saylor, 1943 | MX | MXAL |
| 98 | P. (Phyllophaga) xanthe | (Bates, 1888) | MX | MNHL |
| 99 | P. (Phyllophaga) aff. xanthe | (Bates, 1888) | MX | MXAL |
| 100 | P. (Phyllophaga) collaris | Moser, 1921) | MX | MXAL |
| 101 | P. (Phyllophaga) pubicauda | (Bates, 1888) | MX | MNHL |
| 102 | P. (Phyllophaga) isabellae | Morón y Rivera, 2001 | MX | MXAL |
| 103 | P. (Phyllophaga) blanchardi | Blanchard, 1850 | MX | MNHL |
| 104 | P. (Phyllophaga) leonina | (Bates, 1888) | MX | MNHL |
| 105 | P. (Phyllophaga) beckeri | (Moser, 1921) | MX | ZMHU |
| 106 | P. (Phyllophaga) testaceipennis | (Blanchard, 1850) | MX | MXAL |
| 107 | P. (Phyllophaga) elenans | Saylor, 1938 | CR | MXAL |
| 108 | P. (Phyllophaga) parvisetis | (Bates, 1888) | MX | MNHL |
| 109 | P. (Phyllophaga) temora | Saylor,1943 | MX | IEXA |
| 110 | P. (Phyllophaga) martinezpalaciosi | Morón, 1988 | MX | MXAL |
| 111 | P. (Phyllophaga) setifera | (Burmeister, 1855) | MX | IEXA |
| 112 | P. (Phyllophaga) gigantea | (Bates, 1888) | PA | MNHL |
| 113 | P. (Phyllophaga) maculicollis | LeConte, 1863 | MX | MXAL |
| 114 | P. (Phyllophaga) schizorhina | (Bates, 1888) | PA | MNHL |
| 115 | P. (Phyllophaga) rolbakeri | Saylor, 1940 | MX | MXAL |
| 116 | P. (Phyllophaga) submucida | LeConte, 1856) | EE. UU. | MXAL |
| 117 | P. (Phyllophaga) torta | (LeConte, 1856) | MX | IEXA |
| 118 | P. (Phyllophaga) rugipennis | (Schauffus, 1858) | MX | MXAL |
| 119 | P. (Phyllophaga) tenuipilis | (Bates, 1888) | MX | ZMHU |
| 120 | P. (Phyllophaga) ephilida | (Saylor, 1825) | EE. UU. | MXAL |
| 121 | P. (Phyllophaga) glaberrima | (Blanchard, 1850) | EE. UU. | MNHL |
| 122 | P. (Phyllophaga) molopia | (Bates, 1888) | MX | MNHL |
| 123 | P. (Phyllophaga) segregans | (Bates, 1888) | MX | MXAL |
| 124 | P. (Phyllophaga) porodera | (Bates, 1888) | MX | MXAL |
| 125 | P. (Phyllophaga) eniba | Saylor, 1943 | MX | MXAL |
| 126 | P. (Phyllophaga) fucata | (Horn, 1887) | MX | MXAL |
| 127 | P. (Phyllophaga) opaca | (Moser, 1918) | MX | IEXA |
| 128 | P. (Phyllophaga) ignava | (Horn, 1887) | EE. UU. | MXAL |
| 129 | P. (Phyllophaga) scabrifrons | (Bates, 1888) | GU | MNHL |
| 130 | P. (Phyllophaga) serratipes | Morón and Cano, 2000 | HN | MXAL |
| 131 | P. incertae sedis crenonycha | Saylor, 1943 | MX | MXAL |
| 132 | P. incertae sedis integriceps | (Moser, 1918) | MX | MXAL |
| 133 | P. incertae sedis ilhuicaminai | Morón, 1998 | MX | MXAL |
| 134 | P. incertae sedis pubicollis | Blanchard, 1850 | MX | MXAL |
| 135 | P. incertae sedis sturmi | Bates, 1888 | MX | MNHL |
| 136 | P. incertae sedis stipitalis | (Blanchard, 1850) | MX | MXAL |
| 137 | P. incertae sedis aff. mexicana | (Blanchard, 1850) | MX | MXAL |
| 138 | P. incertae sedis brevipilosa | (Moser, 1918) | VE | ZMHU |
| 139 | P. incertae sedis caanchaki | Morón, 1998 | GT | ZMHU |
| 140 | P. incertae sedis densepunctata | (Moser, 1918) | CO | ZMHU |
| 141 | P. incertae sedis aff. marilucasana | Cano y Morón, 2002 | MX | MXAL |
| 142 | P. incertae sedis chamula | Morón, 1999 | MX | MXAL |
| 143 | P. (Tostegoptera) lanceolata | (Saylor, 1825) | EE. UU. | MXAL |
| 144 | P. (Eugastra) cribrosa | (LeConte, 1853) | EE. UU. | IEXA |
| 145 | P. (Eugastra) epigaea | (Wickham, 1903) | EE. UU. | MXAL |
| 146 | P. (Cnemarachis) plaei | (Blanchard, 1850) | PR | MXAL |
| 147 | P. (Cnemarachis) kenscoffi | Wolcott, 1928 | DO | MXAL |
| 148 | P. (Cnemarachis) mali | Wolcott, 1928 | DO | MXAL |
| 149 | P. (Cnemarachis) dissimilis | (Chevrolat, 1865) | CU | IEXA |
| 150 | P. (Cnemarachis) analis | (Burmeister, 1855) | CU | IEXA |
| 151 | P. (Cnemarachis) dubitata | García- Vidal, 1978 | CU | IEXA |
| 152 | P. (Clemora) smithi | (Arrow, 1912) | BB | MNHL |
| 153 | P. (Clemora) apicalis | (Blanchard, 1850) | PR | MNHL |
Los ejemplares seleccionados se reblandecieron dentro de una cámara húmeda durante un día y se expusieron al vapor de agua durante 30 min. Todos los ejemplares se disectaron para exponer sus piezas bucales, antenas, alas y genitales para su posterior revisión bajo un microscopio estereoscópico Leica Zoom 2000. Para poder observar los tipos de sensores antenales, se retiró la lamela central de un macho de cada especie y se lavó por separado con una solución caliente de KOH al 5% durante 15 min para eliminar residuos de grasa y partículas de polvo, posteriormente se enjuagó con abundante agua y se deshidrató en una solución acuosa de etanol al 70%. Una vez limpias las lamelas centrales, se pegaron por separado sobre soportes adhesivos, se les aplicó al vacío una capa de polvo de oro-paladio, se observaron bajo el microscopio electrónico de barrido Jeol JSM- 5600LV a 550x y 1,800x y se obtuvieron micrografías. Los caracteres morfológicos fueron fotografiados usando una cámara Leica DMC 2900, integrada a un microscopio Leica Z16 AP0A; las imágenes multifocales fueron procesadas con el software Leica Application Suite ver. 4.7.
Análisis filogenético. Se utilizaron 285 caracteres, de los cuales 264 fueron morfológicos y 21 morfométricos (basados en medidas y proporciones), que se obtuvieron con un micrómetro ocular (posteriormente se hizo la transformación de estos caracteres continuos a discretos, los cuales son fácilmente codificables en los análisis filogenéticos). Los estados de carácter fueron codificados con números (0-6); 244 fueron binarios y 41 multiestado. Los caracteres inaplicables se codificaron como «−» y los caracteres que por pérdida de la estructura no se pudieron observar, como «?».
Se empleó el programa Mesquite ver. 3.03 (Maddison y Maddison, 1997) para elaborar la matriz de datos de 153 taxones y 285 caracteres morfológicos (ver material suplementario) que posteriormente se exportó al programa TNT ver. 1.1 (Goloboff, Farris y Nixon, 2008) para realizar el análisis cladístico basado en parsimonia. Todos los caracteres fueron tratados como desordenados (optimización de Fitch), no aditivos y con el mismo peso; los caracteres inaplicables fueron tratados como ausencias (−). La polarización de los estados de carácter se hizo por el método del grupo externo, utilizando a Cyclocephala lunulata Burmeister, 1847 para enraizar el cladograma.
Se hizo una búsqueda tradicional con 1,000 réplicas, usando la estrategia TBR, con las opciones «Wagner trees» y «replace existing trees» habilitadas, con espacio en la memoria para almacenar 1,000 árboles, guardando 10 árboles por cada réplica y con «random seed = 1». Este procedimiento se repitió 10 veces para corroborar que siempre se obtuvieran los mismos valores estadísticos (índice de consistencia e índice de retención). Todos los árboles obtenidos fueron resumidos en un árbol de consenso estricto (Nelsen). Finalmente, el árbol de consenso estricto se sometió a una prueba de bootstrap con 1,000 réplicas para evaluar el soporte de las ramas.
Lista de caracteres morfológicos
Cabeza. 1. Margen apical del prementón: 0) bilobulado, 1) recto; 2. Vestidura del prementón: 0) con sedas sencillas, 1) con sedas escamiformes; 3. Prementón: 0) sin surco longitudinal, 1) con surco longitudinal; 4. Textura de la superficie del mentón: 0) pulida-punteada, 1) rugosa-punteada; 5. Línea transversal en mentón: 0) presente, 1) ausente; 6. Proporciones del labio: 0) semicuadrado, 1) claramente más largo que ancho; 7. Elevaciones laterales en el mentón: 0) presentes, 1) ausentes; 8. Dimensión de la galea maxilar: 0) amplia, 1) reducida; 9. Número de dientes maxilares: 0) 6 dientes, 1) 5 dientes, 2) 4 dientes, 3) 3 dientes, 4) un diente, 5) 7 o más dientes; 10. Diente maxilar distal: 0) entero, 1) astillado; 11. Ápice del diente maxilar distal: 0) entero, 1) bifurcado; 12. Forma del diente maxilar distal: 0) recto, 1) torcido; 13. Ápice de los dientes maxilares: 0) enteros, 1) con helus; 14. Base de la galea cercana a la lacinia: 0) sin un conjunto de 3 dientes pequeños, 1) con un conjunto de 3 dientes pequeños; 15. Base del diente maxilar proximal, observando a la maxila en vista ventral: 0) entero, 1) bifurcado anteriormente; 16. Diente maxilar proximal, observando a la maxila en vista ventral: 0) entero, 1) bifurcado posteriormente; 17. Acomodo de la lacinia maxilar: 0) libre y proyectada, 1) fusionada con la galea; 18. Vestidura de la base de la lacinia maxilar: 0) con sedas muy finas, 1) glabra; 19. Borde dorsal interno de la maxila: 0) con sedas, 1) glabro; 20. Membrana en el borde dorsal interno de la maxila: 0) con pequeña proyección basal de ápice semicuadrado, 1) sin proyección basal; 21. Base del esclerito estipital ii: 0) elevado cerca del esclerito estipital i, 1) nivelado; 22. Esclerito estipital ii: 0) corto, 1) muy alargado; 23. Forma de los palpos maxilares: 0) delgados, 1) engrosados; 24. Forma del sensillum maxilar masculino: 0) circular, 1) alargado, 2) dividido en 4 sets; 25. Sensillum maxilar en hembras: 0) nivelado y simétrico, 1) expuesto y asimétrico; 26. Vestidura del 4.o artejo de los palpos maxilares: 0) con menos de 15 sedas, 1) con más de 20 sedas; 27. Cara exterior del 4.o artejo de los palpos maxilares: 0) sin área sensorial diferenciada, 1) con área sensorial diferenciada; 28. Longitud del último artejo de los palpos maxilares respecto a los 2 anteriores: 0) más largo, 1) no más largo; 29. Posición de las mandíbulas con respecto a los bordes del clípeo en vista dorsal: 0) ocultas, 1) expuestas; 30. Dimensión del lóbulo incisivo mandibular: 0) desarrollado, 1) muy reducido; 31. Dientes en el lóbulo incisivo mandibular: 0) ausentes, 1) presentes y muy bien marcados; 32. Diente interno del lóbulo incisivo mandibular: 0) sin foseta apical, 1) con foseta apical; 33. Margen del lóbulo incisivo mandibular: 0) con muesca, 1) sin muesca; 34. Cara dorsal del lóbulo incisivo mandibular izquierdo: 0) con surco diagonal, 1) sin surco diagonal; 35. Cara dorsal del lóbulo incisivo mandibular derecho: 0) cóncavo, 1) nivelado; 36. Borde externo del lóbulo incisivo mandibular derecho: 0) con proyección cerca del ápice, 1) sin proyección cerca del ápice; 37. Cara interna del lóbulo incisivo mandibular derecho: 0) sin proyección curvada, 1) con proyección curvada; 38. Seno postincisivo: 0) presente, 1) ausente; 39. Coloración del seno postincisivo masculino: 0) del mismo color que el resto de la mandíbula, 1) más clara que el resto de la mandíbula; 40. Dimensión del lóbulo molar mandibular: 0) amplio, 1) reducido; 41. Lóbulo molar de la mandíbula derecha: 0) con escasas estrías muy gruesas, con sus extremos laterales separados, 1) con varias estrías muy paralelas y fusionadas casi por completo, 2) sin un arreglo definido, 3) con un pliegue central; 42. Estrías basales del lóbulo molar de la mandíbula derecha: 0) sin una estría diagonal, 1) con una estría diagonal; 43. Textura de la región media del lóbulo molar mandibular derecho: 0) estriada, 1) lisa; 44. Área proximal del lóbulo molar de la mandíbula derecha: 0) con 2 a 4 quillas proyectadas, 1) con 5 o más quillas proyectadas, 2) con una quilla proyectada; 45. Área distal del lóbulo molar de la mandíbula izquierda: 0) con 2 a 11 quillas proyectadas, 1) con 12 o más quillas proyectadas, 2) con una quilla proyectada; 46. Quilla basal del área distal del lóbulo molar de la mandíbula izquierda: 0) fusionada, 1) libre; 47. Textura de las estrías del lóbulo molar mandibular derecho: 0) lisa, 1) con patrones de pequeños agujeros; 48. Textura de las estrías del lóbulo molar mandibular izquierdo: 0) lisa, 1) con patrones de pequeños agujeros; 49. Lóbulo molar de la mandíbula derecha con 8 o más estrías paralelas: 0) ausente, 1) presente; 50. Forma del borde externo mandibular: 0) curvado o sinuado, 1) recto; 51. Base del labro. 0) articulada con el clípeo, 1) fusionada con el clípeo; 52. Forma del borde antero-basal del labro. 0) homogéneamente bilobulado, 1) heterogéneamente bilobulado, 2) semicircular; 53. Vestidura del borde anterior del labro en vista ventral. 0) con sedas libres dirigidas hacia el centro, 1) con sedas mezcladas con las sedas de la epifaringe; 54. Perfil del borde anterior del labro en vista lateral. 0) vertical, 1) disminuido, 2) diagonal; 55. Número de artejos antenales. 0) 10, 1) 9, 2) 8; 56. Número de lamelas en el macho. 0) solo 3, 1) más de 3; 57. Número de lamelas en la hembra. 0) solo 3, 1) más de 3; 58. Longitud de la maza antenal respecto al funículo en machos. 0) similar o ligeramente más larga, 1) más del doble de larga; 59. Dimorfismo sexual en la longitud de la maza antenal. 0) nulo, 1) fuertemente acentuado; 60. Densidad de sensores antenales a 550x (en la parte media de la cara interna de la lamela central). 0) de 100 a 300, 1) de 0 a 99, 2) más de 400; 61. Porcentaje de sensores basicónicos con respecto al total observado a 550x (en la parte media de la cara interna de la lamela central). 0) de 8 al 50%, 1) de 0 al 7%, 2) más del 51%; 62. Lamela central con superficie estriada. 0) ausente, 1) presente; 63. Cara interna de la lamela central con una línea longitudinal de agujeros grandes e irregulares («entradas de bolsillo»). 0) ausente, 1) presente; 64. Sensilas placodeas. 0) ausentes, 1) presentes; 65. Sensilas auricílicas. 0) ausentes, 1) presentes; 66. Sensilas basicónicas. 0) ausentes, 1) presentes; 67. Sensilas coelocónicas. 0) ausentes, 1) presentes; 68. Poros en la superficie de la lamela. 0) ausentes, 1) presentes; 69. Superficie del centro de la lamela. 0) con textura, 1) lisa; 70. Borde del poro. 0) entero, 1) desvanecido; 71. Sensores sin orden y espacios grandes entre ellos. 0) ausentes, 1) presentes; 72. Franja de sensilas placodeas deformadas. 0) ausentes, 1) presentes; 73. Forma del borde anterior del clípeo en vista dorsal. 0) bilobulado o circular, 1) rectangular o trapezoidal; 74. Parte central del borde anterior del clípeo. 0) recto, 1) inclinado, retraído; 75. Forma de los ángulos latero-apicales del clípeo. 0) redondeados, 1) proyectados; 76. Forma del contorno del clípeo en vista dorsal. 0) subtrapezoidal, 1) subrectangular; 77. Vestidura de la superficie del clípeo. 0) glabro, 1) setoso; 78. Sedas escamiformes en el clípeo. 0) ausentes, 1) presentes; 79. Margen laterobasal del clípeo en vista dorsal. 0) entero, 1) proyectado hacia arriba y afuera; 80. Base del clípeo. 0) sin carina transversal, 1) con carina transversal; 81. Grosor del borde anterior del clípeo en vista lateral. 0) grueso, 1) adelgazado; 82. Forma de la sutura frontoclipeal. 0) sinuada, 1) recta, 2) trapezoidal, 3) en «V»; 83. Continuidad de la sutura frontoclipeal. 0) completa, 1) incompleta; 84. Elevación de la sutura frontoclipeal. 0) nivelada, deprimida o profundamente impresa, 1) ligeramente elevada (solo se aprecia en vista lateral), 2) evidentemente elevada; 85. Textura de la frente. 0) punteada, 1) rugoso-punteada; 86. Vestidura del centro de la frente. 0) glabro, 1) setosa; 87. Vestidura de la región paraocular. 0) glabro, 1) setosa; 88. Zona paraocular. 0) nivelada, 1) elevada; 89. Superficie de la frente. 0) uniforme, 1) irregular; 90. Vértice. 0) sin carina, 1) con carina. 91. Área detrás del vértice. 0) con banda ancha transversal con puntuación, 1) con banda ancha transversal sin puntuación; 92. Porción anterior del ojo en vista dorsal. 0) oculta por el canto ocular, 1) visible por adelante del canto ocular; 93. Forma de los ojos femeninos en vista dorsal. 0) redondeados, 1) alargados; 94. Distancia interocular (ancho de la frente entre el diámetro de los ojos). 0) angosta, caben menos de 5 diámetros oculares (ojos grandes), 1) amplia, caben más de 5 diámetros oculares (ojos chicos); 95. Canto ocular en vista dorsal. 0) formando un ángulo con el clípeo, 1) continuo con el clípeo; 96. Vestidura del canto ocular. 0) setoso, 1) glabro; 97. Cobertura setosa del canto ocular. 0) escasa, menos de 25 sedas, 1) abundante, más de 50 sedas.
Protórax, mesotórax y metatórax. 98. Sedas escamiformes en el canto ocular. 0) ausentes, 1) presentes; 99. Proporciones del pronoto en vista dorsal. 0) subrectangular o casi tan largo como ancho, 1) claramente más largo que ancho; 100. Textura de la superficie dorsal del pronoto. 0) punteada, 1) rugoso-punteada; 101. Puntuación secundaria en el pronoto. 0) ausente, 1) presente; 102. Micropuntuación brillante y superficial entre puntuación primaria. 0) ausente, 1) presente; 103. Puntuación del pronoto. 0) puntuación sin textura interna, 1) puntuación con textura interna; 104. Textura del borde anterior del pronoto. 0) sin puntuación, 1) punteada; 105. Vestidura del borde anterior del pronoto. 0) glabro o con una línea de sedas, 1) con varias hileras de sedas; 106. Textura basal del borde anterior del pronoto. 0) sin puntos grandes que lo penetran casi por completo, 1) con puntos grandes que lo penetran casi por completo; 107. Parte mesial del pronoto, cercana al borde posterior. 0) con puntuación continua, 1) con una línea longitudinal sin puntación (se ve a simple vista); 108. Franja longitudinal con puntuación abundante y pequeña en centro del pronoto. 0) ausente, 1) presente; 109. Región mesial del borde posterior del pronoto. 0) sin puntos grandes con sedas muy largas, 1) con puntos grandes con sedas muy largas; 110. Textura de la zona mesial del borde posterior del pronoto. 0) lisa, 1) finamente punteada; 111. Regiones laterales del borde posterior del pronoto. 0) sin puntos anulares, ovalados y ordenados, 1) con puntos anulares, ovalados y ordenados; 112. Vestidura de la superficie dorsal del pronoto. 0) glabro, 1) setosa; 113. Sedas escamiformes en el pronoto. 0) ausentes, 1) presentes; 114. Patrones longitudinales de sedas blanquecinas en el pronoto. 0) ausentes, 1) presentes; 115. Vestidura del borde lateral del pronoto. 0) con sedas insertadas en la arista del borde lateral, 1) con sedas insertadas dorsalmente, cerca de la arista del borde lateral, 2) con sedas insertadas ventralmente, cerca de la arista del borde lateral; 116. Regiones pruinosas en el pronoto. 0) ausentes, 1) presentes, 117. Margen anterior del pronoto. 0) entero, 1) membranoso; 118. Borde anterior del pronoto. 0) definido, 1) no definido; 119. Grosor del borde anterior en el pronoto masculino. 0) continuo, sin ensanchamiento brusco, 1) engrosado en la zona mesial; 120. Ángulos anteriores del pronoto en vista dorsal. 0) visibles, 1) no visibles; 121. Forma de los ángulos anteriores del pronoto. 0) agudos, 1) redondeados; 122. Formaciones en el margen lateral del pronoto. 0) onduladas o crenuladas, 1) liso; 123. Ángulos posteriores del pronoto. 0) dirigidos hacia los lados, 1) dirigidos hacia la región pleural; 124. Pronoto con bisel posterior. 0) ausente, 1) presente; 125. Borde del pronoto en todo el perímetro. 0) con margen discontinuo, 1) con margen continuo; 126. Borde posterior del pronoto. 0) sin borde, o borde romo, 1) borde con canto plano y puntos con sedas, 2) borde con canto plano y sin puntos; 127. Margen anterior del escutelo. 0) deprimido, pero sin formar una depresión de 90°, 1) formando una depresión de 90° muy bien marcada; 128. Textura del escutelo. 0) punteada, 1) lisa; 129. Región mesial del escutelo. 0) sin quilla tenue longitudinal, 1) con quilla tenue longitudinal; 130. Vestidura del escutelo. 0) glabro, 1) setosa; 131. Sedas escamiformes en el escutelo. 0) ausentes, 1) presentes; 132. Textura elitral. 0) punteada, 1) estriada; 133. Superficie elitral. 0) sin puntuación brillante y superficial, 1) con puntuación brillante y superficial; 134. Vestidura de la superficie elitral. 0) glabra, 1) setosa; 135. Sedas escamiformes en los élitros. 0) ausentes, 1) presentes; 136. Patrones longitudinales de sedas blanquecinas en los élitros. 0) ausentes, 1) presentes; 137. Regiones pruinosas en los élitros. 0) ausentes, 1) presentes; 138. Élitros fusionados en machos. 0) ausentes, 1) presentes; 139. Élitros fusionados en hembras. 0) ausentes, 1) presentes; 140. Relieve de la interestría sutural. 0) elevado, 1) apenas marcado; 141. Textura de la interestría sutural. 0) punteada, 1) rugosa; 142. Densidad de la puntuación o rugosidad de la interestría sutural. 0) igual que en los élitros, 1) menos densa que en los élitros; 143. Textura del margen membranoso epipleural. 0) sin margen membranoso, 1) lisa, 2) corrugada; 144. Filo del margen epipleural. 0) entero, 1) aterciopelado; 145. Textura de la zona media epipleural. 0) lisa, 1) con pars stridens; 146. Vestidura de la epipleura. 0) setosa, 1) glabra; 147. Acomodo de sedas en la epipleura. 0) sedas en orden, de mayor a menor, 1) sedas largas mezcladas con cortas; 148. Sedas en la epipleura. 0) en todo el margen, 1) solo hasta la mitad anterior, 2) cubren 3/4 del margen (a partir de la base), 3) solo en la base; 149. Sedas en el ápice de la epipleura. 0) ausentes, 1) presentes; 150. Sedas entre los élitros, naciendo en el borde externo. 0) presentes, 1) ausentes; 151. Sedas entre los élitros, naciendo en el borde interno. 0) ausentes, 1) presentes; 152. Forma del perfil elitral en vista lateral. 0) aplanado, 1) convexo, 2) ligeramente deprimido al centro; 153. Dimensión de las alas membranosas en los machos. 0) desarrolladas, 1) reducidas; 154. Forma de las alas membranosas. 0) sin lóbulo inferior, 1) con lóbulo inferior; 155. Color de alas membranosas. 0) ámbar, 1) ahumadas; 156. Relación de proporción de largo-ancho de las alas membranosas. 0) cortas (razón menor a 3), 1) largas (razón mayor a 3); 157. Ápice de la vena J. 0) terminando antes del margen, 1) uniéndose al margen; 158. Forma de la vena A2. 0) «Y», 1) «T»; 159. Región proximal de la vena A2. 0) curvada, pero sin formar un ángulo de 90°, 1) formando un ángulo de 90°; 160. Proyección media de la vena A2. 0) corta, semiperpendicular a la vena A1, 1) prolongada, semiparalela a la vena A1; 161. Delimitación de la celda basal. 0) casi cerrada entre A1 y A2, 1) cerrada entre A1 y A2; 162. Vena espuria entre cubital y A1. 0) presente, sin tocar el margen alar, 1) presente, tocando el margen alar, 2) ausente; 163. Ápice de la vena cubital. 0) ligeramente curvado, 1) ligeramente sinuado; 164. Forma de la vena M. 0) sencilla, 1) ligeramente curvada en el ápice; 165. Sutura entre el propleurón y el basiesternón. 0) apenas marcada y nivelada, 1) muy visible y elevada; 166. Solapa ocular ventral. 0) ausente, 1) presente; 167. Regiones pruinosas en la propleura. 0) ausentes, 1) presentes; 168. Basiesternón. 0) con 2 quillas, 1) liso, 2) con una quilla; 169. Vestidura del basiesternón. 0) setosa, 1) glabra; 170. Proceso proesternal 0) poco proyectado, 1) proyectado hasta la altura de los trocánteres anteriores; 171. Forma del proceso proesternal. 0) semicircular, agudo, bilobulado o bidentado, 1) bidentado en el ápice y proyectado diagonalmente en el centro; 172. Dimensión del metaesternón en machos. 0) alargado, 1) corto; 173. Sedas centrales del metaesternón. 0) finas y sedosas, 1) como espinas muy largas, 2) escamiformes.
Abdomen. 174. Regiones pruinosas en los esternitos. 0) ausentes, 1) presentes; 175. Esternitos masculinos con tumescencias paramesiales suavemente redondeadas. 0) ausentes, 1) presentes; 176. Región mesial del primer esternito masculino. 0) liso, 1) con 2 quillas simétricas en su base; 177. Sutura de los esternitos en su región paramesial. 0) continua, 1) con un surco transversal, ligeramente ensanchado, rugoso y oscurecido; 178. Suturas en los esternitos. 0) funcionales, 1) no funcionales; 179. Región mesial de los esternitos. 0) con sutura visible, 1) con sutura desvanecida; 180. Últimos esternitos modificados. 0) ausentes, 1) presentes; 181. Regiones laterales del último esternito. 0) sin depresión, 1) con surco transversal amplio; 182. Borde posterior del quinto esternito masculino con una angosta franja esclerosada. 0) ausente, 1) presente; 183. Borde posterior del quinto esternito femenino con una angosta franja esclerosada. 0) ausente, 1) presente; 184. Borde del último espiráculo. 0) no definido, 1) definido; 185. Forma del último espiráculo. 0) circular, 1) ovalado; 186. Suturas del espiráculo. 0) presentes, 1) ausentes; 187. Puntuación de la región mesial de la placa anal masculina. 0) uniforme, 1) con una pequeña zona multipunteada o rugosa; 188. Placa anal masculina. 0) sin surco transversal, 1) con surco transversal; 189. Zona mesial de la placa anal masculina. 0) sin surco longitudinal, 1) con surco longitudinal; 190. Región mesial de la base de la placa anal masculina. 0) sin un proceso bidentado, 1) con un proceso bidentado; 191. Región mesial del ápice de la placa anal masculina. 0) sin un proceso bidentado/unidentado, 1) con un proceso unidentado o bidentado; 192. Ápice de la placa anal femenina. 0) entero, 1) con escotadura apical, 2) ligeramente proyectado; 193. Ápice de la placa anal masculina. 0) entero, 1) con escotadura apical; 194. Placa anal muy expandida. 0) ausente, 1) presente; 195. Ápice del propigidio. 0) expuesto, 1) oculto por el ápice de los élitros; 196. Propigidio. 0) sin surco mesial, 1) con surco mesial; 197. Unión entre propigidio y placa pigidial en machos. 0) cada estructura separada claramente de la otra, 1) casi fusionadas; 198. Borde lateroapical de la placa pigidial. 0) con margen continuo, 1) con margen laminar y respingado, 2) sin margen; 199. Borde basal de la placa pigidial. 0) con margen interrumpido, 1) con margen continuo; 200. Vestidura del disco pigidial. 0) setosa, 1) glabra; 201. Sedas escamiformes en el pigidio. 0) ausentes, 1) presentes; 202. Disco pigidial en hembras. 0) simple, 1) con prominencias; 203. Textura pigidial. 0) punteada, 1) rugosa; 204. Perfil lateral del pigidio masculino. 0) convexo, 1) casi plano; 205. Perfil lateral del pigidio femenino. 0) convexo, 1) casi plano, 2) cóncavo; 206. Sedas apicales del pigidio en hembras. 0) insertadas en la parte externa del borde, 1) insertadas en la parte interna del borde, 2) insertadas antes del borde.
Genitales masculinos. 207. Perímetro dorsal del tecto. 0) similar o más angosto que el de la pieza basal, 1) más ancho que el de la pieza basal; 208. Ápice del tecto con carina transversal. 0) ausente, 1) presente; 209. Regiones condilares apicales del tecto. 0) sin desarrollo, 1) desarrolladas; 210. Regiones condilares basales del tecto. 0) sin desarrollo, 1) desarrolladas; 211. Longitud del tecto respecto a la pieza basal. 0) similar o más largo, 1) más corto; 212. Forma de la pieza basal. 0) sencilla, 1) semiesférica, 2) esbelta y aplanada; 213. Continuidad de la pieza basal. 0) continua con el tecto, 1) separada del tecto cerca del ápice de la cápsula genital, 2) separada del tecto cerca de la mitad de la cápsula genital; 214. Dimensión de los cóndilos laterobasales en la pieza basal. 0) no desarrollados, 1) desarrollados; 215. Forma de los cóndilos laterobasales en la pieza basal. 0) simétricos, 1) asimétricos; 216. Reborde del foramen basal en la pieza basal. 0) sin reborde, 1) con reborde marcado; 217. Falobase más elevada que el tecto. 0) ausente, 1) presente; 218. Falobase ensanchada dorso-ventralmente. 0) ausente, 1) presente; 219. Falobase y parámeros fusionados dorsalmente en su extremo basal. 0) presente, 1) ausente; 220. Falobase y parámeros formando una acanaladura longitudinal dorsal. 0) ausente, 1) presente; 221. Falobase y parámeros formando un tubo muy alargado, estrecho y curvado hacia arriba. 0) ausente, 1) presente; 222. Falobase y parámeros formando un tubo engrosado y curvado hacia abajo, con el ápice ancho y comprimido. 0) ausente, 1) presente; 223. Simetría de la falobase y parámeros. 0) simétricos, 1) asimétricos; 224. Ápice de los parámeros como placas anchas y simétricas. 0) ausente, 1) presente; 225. Parámeros excavados, muy extendidos y asimétricos. 0) ausente, 1) presente; 226. Razón de longitud de la pieza basal-tecto respecto a la longitud de la falobase-parámeros. 0) más de 1 (parámeros cortos), 1) 1 o menos (parámeros largos); 227. Forma del spiculum gastrale. 0) «Y», 1) «V», 2) «U»; 228. Longitud de la porción basal del spiculum gastrale.0) larga (su longitud es mayor o igual que la longitud de los brazos), 1) corta (su longitud es menor que la longitud de los brazos), 2) ausente.
Genitales femeninos. 229. Placas dorsales de los genitales femeninos. 0) separadas, 1) fusionadas entre sí; 230. Proceso púbico. 0) ausente, 1) con formas lobuladas variables, 2) agudo y muy alargado; 231. Tamaño de las placas dorsales o superiores. 0) pequeñas, 1) grandes; 232. Ápice lateral interno de las placas ventrales o inferiores. 0) entero, sin proyecciones, 1) con una proyección alargada y setosa; 233. Vestidura de las placas ventrales o inferiores. 0) con sedas escasas, 1) glabras; 234. Placas superiores e inferiores. 0) ambas presentes, 1) solo las inferiores.
Extremidades. 235. Espolón apical protibial en machos. 0) presente, 1) ausente; 236. Espolón apical protibial en hembras. 0) presente, 1) ausente; 237. Forma del espolón apical protibial. 0) recto, 1) ligeramente curvado; 238. Alcance del espolón protibial en machos. 0) rebasando el ápice interno de la protibia, 1) sin rebasar el ápice interno de la protibia; 239. Alcance del espolón protibial en hembras. 0) rebasando el ápice interno de la protibia, 1) sin rebasar el ápice interno de la protibia; 240. Número de dentículos bien definidos en las protibias de los machos. 0) 2 o más, 1) menos de 2; 241. Número de dentículos bien definidos en las protibias de las hembras. 0) 3, 1) 2; 242. Dimorfismo sexual en las protibias (diferencia en número de dientes externos y presencia de espolón protibial). 0) nulo, 1) marcado; 243. Protibias en vista ventral. 0) con carina longitudinal, 1) sin carina longitudinal; 244. Región interna de las protibias en vista ventral, en ambos sexos. 0) con sedas delgadas, largas y erectas, 1) con 2 a 7 sedas espiniformes; 245. Coxas anteriores. 0) transversales y poco sobresalientes, 1) semicónicas y sobresalientes; 246. Carina mesotibial. 0) completa, 1) incompleta; 247. Quilla corta incompleta mesotibial. 0) presente, 1) ausente; 248. Carina metatibial. 0) completa, 1) incompleta; 249. Quilla corta incompleta metatibial. 0) presente, 1) ausente; 250. Lado externo de las meso y metatibias (con el ejemplar en vista lateral). 0) entero, 1) con dentículos, 2) aserrado; 251. Borde del lado externo de las metatibias. 0) romo, 1) marcado; 252. Ápice de las metatibias masculinas. 0) con 2 espolones, 1) sin espolones; 253. Ápice de las metatibias con una corona de sedas gruesas, largas y fuertes (como espolones). 0) ausente, 1) presente; 254. Tamaño del espolón metatibial interno respecto al externo. 0) notablemente mayor, 1) más corto, similar o ligeramente mayor; 255. Espolón metatibial externo masculino. 0) articulado, 1) fusionado con el borde apical de la metatibia; 256. Extremo distal de la metatibia masculina. 0) sin una carina cercana a la base del espolón metatibial externo, 1) con una carina cercana a la base del espolón metatibial externo; 257. Vestidura de la base del espolón metatibial externo masculino. 0) glabro, 1) setoso; 258. Vestidura de la base del espolón metatibial externo femenino. 0) glabro, 1) setoso; 259. Sedas espiniformes apicales de metatibia femenina. 0) ordenadas, 1) encimadas; 260. Metafémur con plectrum dorsoapical. 0) ausente, 1) presente; 261. Protarsómeros masculinos, 0) alargados, 1) cortos; 262. Ápice interno de los protarsómeros. 0) entero, 1) proyectado, formando un pequeño espolón fusionado; 263. Forma del ápice interno de los protarsómeros. 0) romo, 1) agudo; 264. Quilla estrecha longitudinal en la región ventral de los protarsómeros 1.o-4.o. 0) formando una línea longitudinal, 1) formando 2 líneas longitudinales, 2) regiones ventrales modificadas por la abundancia de sedas, 3) región ventral pulida; 265. Forma de la quilla estrecha longitudinal en la región ventral de los tarsos. 0) entero, 1) aserrado-pectinado; 266. Región ventral de los protarsómeros en machos. 0) lisos, 1) con textura estrigada en los tarsómeros 1.o-4.o, 2) con textura estrigada en los tarsómeros 3.o-4.o, 3) con textura estrigada en el basitarso; 267. Protarsómeros masculinos muy ensanchados ventralmente. 0) ausente, 1) presente; 268. Ápice interno de los mesotarsómeros. 0) entero, 1) proyectado, formando un pequeño espolón fusionado; 269. Región ventral de los mesotarsómeros en machos. 0) lisos, 1) con textura estrigada en los tarsómeros 1.o-4.o, 2) con textura estrigada en los tarsómeros 3.o-4.o; 270. Sedas de la región ventral de los metatarsómeros. 0) delgadas, sin orden, 1) gruesas y ordenadas, formando un patrón pectinado; 271. Sedas dorsales de los metatarsómeros. 0) más abundantes que las ventrales, 1) igual de abundantes que las ventrales, 2) solo hay una línea de sedas, 3) menos abundantes que las ventrales; 272. Ápice interno del primer metatarsómero en vista ventral. 0) entero, 1) con un espolón fusionado; 273. Tamaño del primer metatarsómero masculino. 0) más corto que el segundo, 1) igual de corto que el segundo; 274. Cobertura setosa de la región ventral de todos los tarsos. 0) escasa, 1) moderadamente densa, 2) muy abundante; 275. Forma de las uñas protarsales masculinas. 0) bidentadas, 1) tridentadas, 2) bífidas o hendidas, 3) pectinadas, 4) bipectinadas, 5) sencillas, 6) bífidas con dentículo basal; 276. Margen inferior del dentículo intermedio de las uñas protarsales. 0) entero, 1) finamente aserrado, 2) pectinado; 277. Similitud entre uñas protarsales. 0) similares, 1) parecidas, 2) diferentes; 278. Forma de las uñas protarsales entre macho y hembra. 0) similares, 1) diferentes; 279. Cara interna de las uñas protarsales. 0) roma, 1) aplanada; 280. Forma de las uñas pro y mesotarsales masculinas. 0) similares, 1) diferentes; 281. Uñas metatarsales. 0) ornamentadas, 1) sencillas; 282. Número de uñas metatarsales. 0) 2 uñas, 1) 1 uña; 283. Forma de las uñas metatarsales masculinas. 0) bidentadas, 1) tridentadas, 2) bífidas o hendidas, 3) pectinadas, 4) bipectinadas, 5) sencillas, 6) bífidas con dentículo basal; 284. Forma de las uñas pro y metatarsales masculinas. 0) similares, 1) más o menos diferentes, 3) completamente diferentes; 285. Razón corporal (determinado como la longitud total, desde el ápice del clípeo hasta el ápice de los élitros, dividido entre el ancho máximo elitral) 0) robusto (razón menor a 2), 1) esbelto (razón de 2 o más).
Resultados
Se obtuvieron 10 cladogramas igualmente parsimoniosos, con una longitud de 2,802 pasos, IC = 0.12 e IR = 0.56. En cada una de las 10 repeticiones se obtuvieron los mismos valores de los estadísticos descriptivos y los mismos árboles. El cladograma de consenso estricto se muestra en las Figura 1, Figura 2. En primera instancia se observa que el agrupamiento tradicionalmente denominado Phyllophaga sensu lato es polifilético, lo cual confirma los resultados obtenidos por Rivera-Gasperín y Morón, 2013, Rivera-Gasperín y Morón, 2017, y fundamenta la necesidad de reestructurar su clasificación supragenérica. En esta topología (Figura 1, Figura 2), el clado en donde se encuentra Chlaenobia + Chirodines, junto con P. Phytalus lineata Bates, 1888, Phyllophaga aff. marilucasanaCano y Morón, 2002 y P. chamula Morón, 1999 se encuentra relacionado con el linaje de especies que contiene a los subgéneros Phytalus, Phyllophaga, Tostegoptera Blanchard, 1850, Eugastra LeConte, 1856 y Listrochelus y a los géneros Trichesthes Erichson, 1847 y Catrachia Coca-Abia y Robbins, 2006, con los cuales comparte el sensillum maxilar masculino alargado, borde posterior del quinto esternito masculino con una angosta franja esclerosada y protibias en vista ventral con una carina longitudinal (Fig. 3, nodo 1, caracteres 24:1, 182:1 y 243:0).
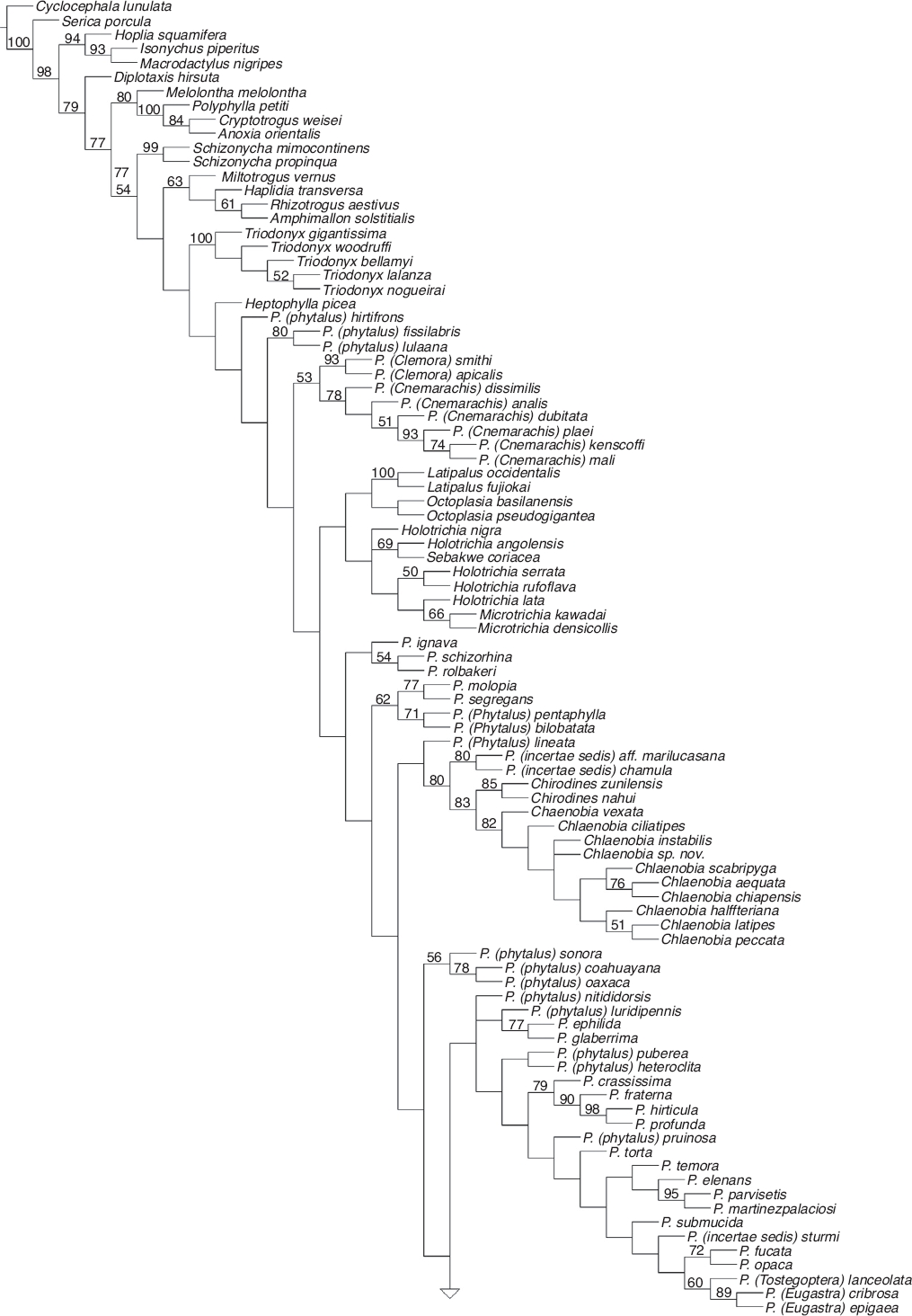
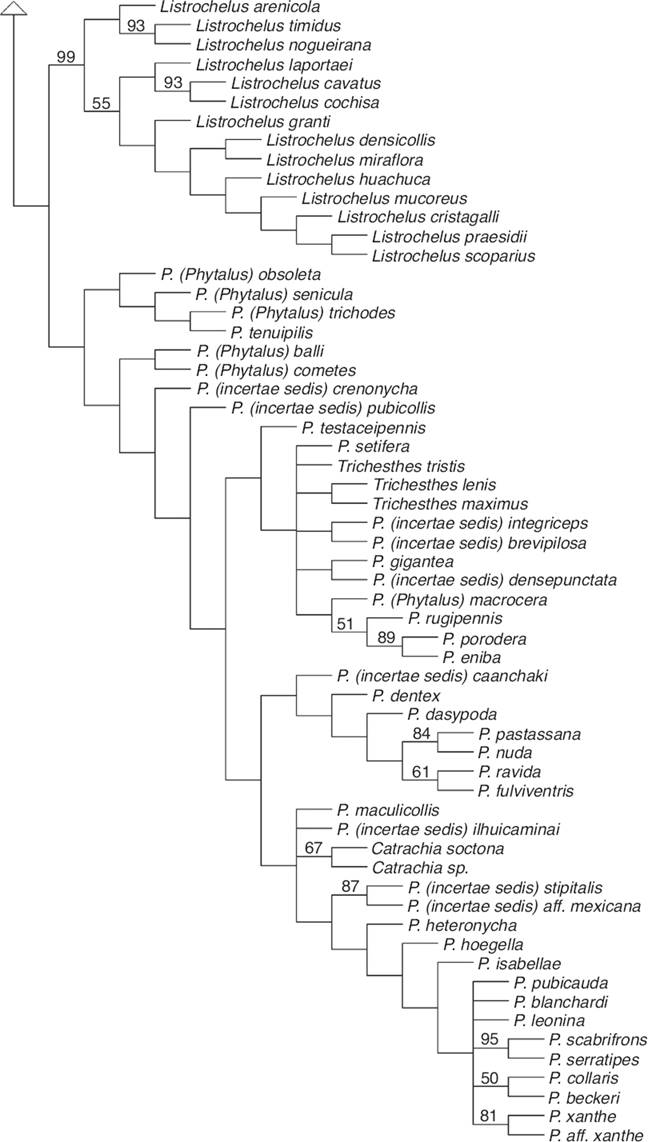
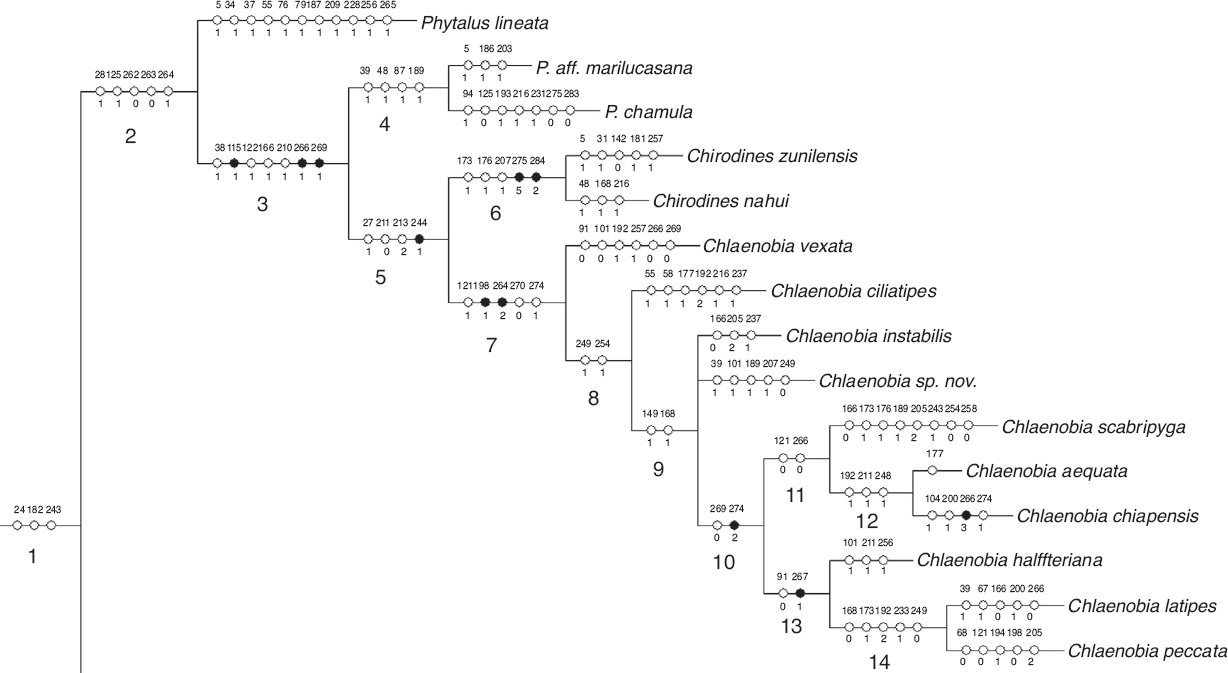
Figura 3 Clado de interés. El número de carácter se muestra sobre las líneas, el estado de carácter bajo las líneas (0-5); los círculos negros representan sinapomorfías, los círculos blancos son homoplasias; y los números grandes (1-14) enumeran los nodos.
En la Figura 3 se numeraron los nodos del 1 al 14, para facilitar su descripción detallada. En el nodo 2 se ubican P. Phytalus lineata, Phyllophaga aff. marilucasana y P. chamula junto con las especies de Chlaenobia y Chirodines por presentar una combinación de 5 estados de carácter homoplásicos: último artejo de los palpos maxilares más corto o similar respecto a los 2 anteriores, borde del pronoto con margen continuo en todo el perímetro, ápice interno de los protarsómeros entero y ensanchado y región ventral de los protarsómeros 1.o-4.ocon quilla longitudinal doble (caracteres 28:1, 125:1, 262:0, 263:0 y 264:1).
En el nodo 3 se ubican Phyllophaga aff. marilucasana y P. chamula junto con las especies de Chlaenobia y Chirodines. Este nodo se encuentra soportado por un alto valor de bootstrap del 80% y 3 sinapomorfías: borde lateral del pronoto con sedas insertadas dorsalmente, cerca de la arista del borde lateral (Fig. 4) y región ventral de los protarsómeros y mesotarsómeros masculinos (del 1.o-4.o) con superficie estrigada (microestrías formando un patrón semejante a una huella digital) (Figs. 5 y 6) (caracteres 115:1, 266:1 y 269:1); además, este grupo también comparte otros 4 caracteres que no son exclusivos para el clado: borde externo del lóbulo incisivo mandibular derecho sin proyección cerca del ápice, margen lateral del pronoto liso, solapa ocular ventral presente y calla basales del tecto desarrolladas (caracteres 36:1, 122:1, 166:1 y 210:1). En el nodo 4 se encuentra el grupo hermano de Chlaenobia + Chirodines, representado en este estudio por las especies P. aff. marilucasana y P. chamula. Este clado presenta un valor de bootstrap del 80% y 4 caracteres homoplásicos: coloración del seno postincisivo masculino más clara que el resto de la mandíbula, estrías del lóbulo molar mandibular izquierdo con patrones de pequeños agujeros, región paraocular setosa y zona mesial de la placa anal masculina con un surco longitudinal (caracteres 39:1, 48:1, 87:1 y 189:1).

Figura 4-11 Caracteres morfológicos sinapomórficos: 4) pronoto en vista lateral de C. peccata, con detalle de las sedas insertadas dorsalmente; 5) protarsómero en vista ventral de C. zunilensis, con detalle de la textura estrigada; 6) mesotarsómero en vista ventral de C. nahui; 7) protibia en vista ventral de C. halffteriana; 8) uñas protarsales de C. zunilensis; 9) uñas metatarsales de C. zunilensis; 10) placa pigidial de C. halffteriana; 11) protarsómeros en vista lateral de C. latipes. Líneas = 0.5 mm.
En el nodo 5 se reúnen todas las especies que corresponden al concepto tradicional de Chlaenobia y Chirodines (bootstrap = 83%). Estas especies comparten una sinapomorfía: región interna de las protibias en vista ventral con 2 a 7 sedas espiniformes (Fig. 7, carácter 244:1); y 3 caracteres que no son únicos para el clado: cara exterior del 4.o artejo de los palpos maxilares con área sensorial diferenciada, longitud del tecto similar o más largo respecto a la pieza basal y pieza basal separada del tecto cerca de la mitad de la cápsula genital (caracteres 27:1, 211:0 y 213:2).
Con un valor de bootstrap del 85%, en el nodo 6 se hermanan las especies tradicionalmente tratadas como Chirodines por compartir 2 sinapomorfías: uñas protarsales masculinas sencillas (Fig. 8) y uñas pro- y metatarsales masculinas completamente diferentes entre ellas (Figs. 8 y 9) (caracteres 275:5 y 284:2); y 3 caracteres no exclusivos para el clado: sedas centrales del metaesternón como espinas muy largas, región mesial del primer esternito masculino con 2 quillas simétricas en su base y perímetro dorsal del tecto más ancho que el de la pieza basal (caracteres 173:1, 176:1 y 207:1).
El nodo 7 (bootstrap = 82%) contiene a las especies tradicionalmente definidas como Chlaenobia. Estas 10 especies comparten 2 estados de carácter sinapomórficos: borde lateroapical de la placa pigidial con margen laminar y respingado (Fig. 10) y quilla estrecha longitudinal en la región ventral de los protarsómeros ausente (Fig. 8) (caracteres 198:1 y 264:2); además de 3 homoplasias: ángulos anteriores del pronoto redondeados, sedas de la región ventral de los metatarsómeros delgadas y sin orden y cobertura setosa moderadamente densa en la región ventral de todos los tarsos (caracteres 121:1, 270:0 y 274:1). En el nodo 8 se agrupan las especies que comparten 2 estados de carácter homoplásicos: quilla metatibial corta incompleta ausente y espolón metatibial interno más corto, similar o ligeramente mayor con respecto al externo (caracteres 249:1 y 254:1). En el nodo 9 se observa una politomía que involucra a C. instabilis, C. sp. nov. y las especies clasificadas por Morón (1986) dentro del «grupo latipes», las cuales comparten 2 estados de carácter que no son exclusivos para el clado: ápice de la epipleura con sedas y basiesternón liso (caracteres 149:1 y 168:1).
En el nodo 10 se encuentran las especies del «grupo latipes», las cuales comparten una sinapomorfía: cobertura setosa muy abundante en la región ventral de todos los tarsos (Fig. 11, carácter 274:2); y un estado de carácter no exclusivo: mesotarsómeros masculinos con región ventral lisa (carácter 269:0). En el nodo 11, se agrupan las especies C. scabripyga, C. aequata y C. chiapensis, pues comparten 2 estados de carácter que también se observan en otros clados: pronoto con ángulos anteriores agudos y protarsómeros masculinos con región ventral lisa (caracteres 121:0 y 266:0). En el siguiente nodo (12) y con un valor de bootstrap de 76%, se hermanan las especies C. aequata y C. chiapensis, estas especies comparten 3 caracteres homoplásicos: ápice de la placa anal femenina con escotadura apical, tecto más corto respecto a la pieza basal y carina metatibial incompleta (caracteres 192:1, 211:1 y 248:1).
Chlaenobia halffteriana Morón, 1992, C. latipes y C. peccata se reúnen en el nodo 13, pues comparten una sinapomorfía: protarsómeros masculinos muy ensanchados ventralmente (Fig. 11, carácter 267:1); y un carácter no exclusivo para el clado: área detrás del vértice con banda ancha transversal con puntuación (carácter 91:0). Por último, en el nodo 14 (bootstrap = 51%), se relacionan C. latipes y C. peccata, por compartir una combinación de 5 estados de carácter: basiesternón con 2 quillas, sedas centrales del metaesternón como espinas muy largas, ápice de la placa anal femenina ligeramente proyectado, placas ventrales de la genitalia femenina glabras y quilla corta incompleta metatibial presente (caracteres 168:0, 173:1, 192:2, 233:1 y 249:0).
Discusión
En 1940, Saylor decidió tratar a Chlaenobia y Chirodines como subgéneros de (Phyllophaga), pues consideró que la estructura de las uñas y la vestidura setífera densa de los tarsos no eran caracteres suficientemente fuertes para mantenerlos como géneros válidos. Sin embargo, el cladograma obtenido en este estudio muestra que Chlaenobia-Chirodines es un grupo monofilético, soportado por un valor alto de bootstrap = 80% y 7 caracteres morfológicos, de los cuales 3 son sinapomorfías para el grupo (caracteres 115:1, Fig. 4; 266:1, Fig. 5; 269:1, Fig. 6) y 4 no son exclusivos para el clado (caracteres 36:1, 122:1, 166:1 y 210:1).
Basándonos en los resultados obtenidos y siguiendo los lineamientos establecidos en el Capítulo 6, Artículo 23 (Principio de Prioridad) del Código Internacional de Nomenclatura Zoológica (ICZN, 1999), se propone la restitución del género Chlaenobia Blanchard, 1850, e incluir a las especies originalmente descritas como Chirodines Bates, 1888. De las especies analizadas, P. chamula y P. aff. marilucasana se encuentran estrechamente relacionadas con el clado de interés, pero sus semejanzas con P. Phytalus lineata y otras especies no incluidas en el presente análisis podrían justificar la formación de otro grupo de especies hermano de Chlaenobia, cuyo estudio se encuentra en proceso junto con la revisión formal del género Chlaenobia (Morón, observ. pers.).
Chlaenobia Blanchard, 1850, estatus revalidado
Chlaenobia Blanchard, 1850: 116
Especie tipo: Chlaenobia ciliatipes Blanchard, 1850, monotípico (Museum d’Histoire Naturelle de Paris). Subsecuentemente confirmado por Saylor (1942. p.161).
Phyllophaga (Chlaenobia), Aragón y Morón (2003. p. 560), Arnett (1973. p. 404), Cano y Morón, 1998, Cano y Morón, 2002, Coca-Abia et al. (1993. p. 268), Morón, 1986, Morón, 1988, Morón, 1992, Morón, 2003, Morón y Robbins (2004. p. 331), Saylor, 1940, Saylor, 1942.
Redescripción
Los adultos de Chlaenobia pueden separarse de otros géneros por presentar la siguiente combinación de caracteres: cuerpo alargado y esbelto, nunca ensanchado hacia atrás; de color amarillento pajizo a rojizo o blanquecino; regiones dorsales glabras y brillantes, sin sedas escamiformes ni zonas pruinosas. Mentón rugoso o punteado, con sedas escasas, disco cóncavo y ápice bilobulado. Maxilas con 5 dientes, diente maxilar proximal entero; galea amplia; base del esclerito estipital ii elevado cerca del esclerito estipital i; palpos maxilares delgados, el cuarto artejo con vestidura escasa, más corto o similar con respecto a los 2 anteriores; área sensorial diferenciada, sensillum alargado. Lóbulo incisivo mandibular con muesca; cara dorsal del lóbulo incisivo mandibular izquierdo con surco diagonal. Seno postincisivo presente. Lóbulo molar de la mandíbula derecha con escasas estrías gruesas, región media semiplana o estriada; algunas especies presentan patrones de pequeños agujeros bajo las estrías de ambas mandíbulas. Antenas con 9 o 10 artejos; maza antenal con 3 lamelas. Clípeo glabro, subtrapezoidal, con ápice bilobulado o circular y ángulos lateroapicales redondeados. Sutura frontoclipeal sinuada, completa y nivelada. Frente glabra, punteada, con superficie uniforme, sin carina transversal. Canto ocular con cobertura setosa escasa. Pronoto con puntuación fina y uniforme, en ocasiones con puntación secundaria; borde lateral liso, sin ondulaciones, con sedas insertadas dorsalmente, cerca de la arista; borde posterior romo, sin puntuación anular ni sedas largas; ángulos posteriores obtusos, redondeados o casi rectos, en ocasiones replegados hacia la región pleural. Escutelo punteado, con margen anterior ligeramente deprimido. Élitros libres, punteados, sin estrías; interestría sutural elevada; perfil elitral aplanado. Epipleura setosa, con margen liso, sin textura aterciopelada o corrugada. Alas membranosas desarrolladas, sin lóbulo inferior; celda basal casi cerrada entre A1 y A2; vena espuria bien marcada entre cubital y A1, sin tocar el margen alar inferior. Solapa ocular ventral presente. Basiesternón con una o 2 quillas. Metaesternón alargado, en ocasiones con sedas centrales espiniformes. Esternitos inarticulados; región mesial de los esternitos con suturas visibles; últimos esternitos sencillos, sin modificaciones; algunas especies presentan el borde posterior del quinto esternito con una franja esclerosada angosta. Placa anal con puntuación uniforme, sin surco transversal, en ocasiones con surco longitudinal. Propigidio sin surco mesial. Pigidio glabro o setoso; con margen apical continuo o laminar y respingado; borde basal con margen continuo o interrumpido. Protibias con 2 a 7 sedas espiniformes en la región ventral. Espolón metatibial interno más corto, similar o ligeramente mayor respecto al externo. Región ventral de los protarsómeros 1.o-4.ocon quilla longitudinal doble. Uñas tarsales hendidas o sencillas, nunca bidentadas o pectinadas. Genitales masculinos con pieza basal sencilla, continua o separada del tecto cerca de la mitad de la cápsula genital, con cóndilos laterobasales desarrollados y simétricos; calla basales del tecto desarrolladas; parámeros cortos. Genitales femeninos formados por cuatro placas independientes, sin proceso púbico. Dimorfismo sexual marcado: machos con maza antenal más larga que en hembras; protibias con 2 o 3 dientes externos; región ventral de los pro- y mesotarsómeros lisa o estrigada; todos los artejos tarsales provistos con una cobertura setífera moderadamente densa o muy abundante; uñas protarsales hendidas o sencillas. Hembras con pigidio usualmente excavado o tuberculado; ápice de la placa anal entero, con escotadura o ligeramente proyectado; base del espolón metatibial externo con sedas.
Especies incluidas en Chlaenobia Blanchard, 1850.
1. Chlaenobia aequata Bates, 1888. Chlaenobia aequata Bates, 1888. Biologia Centrali-Americana, 2: 168. Distribución: Costa Rica, Nicaragua.
2. Chlaenobia chiapensis Chapin, 1935. Chlaenobia chiapensis Chapin, 1935. Smithsonian Miscellaneous Collections, 94(9): 18. (Morón, 2003). Distribución: Costa Rica, El Salvador, Guatemala, Honduras, México.
3. Chlaenobia ciliatipes Blanchard, 1850. Chlaenobia ciliatipes Blanchard, 1850. Catalogue de la Collection Entomologique. Classe des insectes: 116. Distribución: Guatemala, México. Chlaenobia aegrota Bates, 1888 (Morón, 2003).
4. Chlaenobia etabatesiana (Morón, 1992). Phyllophaga (Chlaenobia) etabatesiana Morón, 1992. Giornale Italiano di Entomologia, 6(30): 28. Distribución: México.
5. Chlaenobia halffteriana (Morón, 1992). Phyllophaga (Chlaenobia) halffteriana Morón, 1992. Giornale Italiano di Entomologia, 6(30): 16. Distribución: Costa Rica, Honduras, México.
6. Chlaenobia howdeniana (Morón, 1992). Phyllophaga (Chlaenobia) howdeniana Morón, 1992. Giornale Italiano di Entomologia, 6(30): 22. Distribución: México.
7. Chlaenobia instabilis (Blackwelder, 1944). Phyllophaga instabilis Blackwelder, 1944. United States National Museum Bulletin, 185(2): 225. Distribución: México. Chlaenobia arrowi Chapin, 1935 (Phyllophaga instabilis fue el nombre de reemplazo para Chlaenobia arrowi, homónimo secundario de Lachnosterna arrowi Saylor, 1935).
8. Chlaenobia latipesBates, 1888. Chlaenobia latipes Bates, 1888. Biologia Centrali-Americana, 2: 167. Distribución: El Salvador, Guatemala, México. Chlaenobia bicallosa Bates, 1888 (Chapin, 1935).
9. Chlaenobia lempira (Morón y Robbins, 2004). Phyllophaga (Chlaenobia) lempira Morón y Robbins, 2004. Anales del Instituto de Biología, UNAM, Serie Zoología, 75(2): 332. Distribución: Honduras.
10. Chlaenobia nahui (Morón y Solís, 1994). Phyllophaga (Chirodines) nahui Morón y Solís, 1994. Giornale Italiano di Entomologia, 7: 182. Distribución: Costa Rica.
11. Chirodines ome (Morón, 1991). Phyllophaga (Chirodines) ome Morón, 1991. Giornale Italiano di Entomologia, 5: 400. Distribución: México.
12. Chlaenobia panama Chapin, 1935. Chlaenobia panama Chapin, 1935. Smithsonian Miscellaneous Collections, 94(9): Distribución: Costa Rica, Panamá.
13. Chlaenobia peccata (Blackwelder, 1944). Phyllophaga peccatus Blackwelder, 1944. United States National Museum Bulletin, 185(2): 225. Distribución: México. Chlaenobia colimana Arrow, 1933 (Phyllophaga peccatus fue el nombre de reemplazo para Chlaenobia colimana, homónimo secundario de Lachnosterna colimana Moser, 1921).
14. Chlaenobia personataChapin, 1935. Chlaenobia personata Chapin, 1935. Smithsonian Miscellaneous Collections, 94(9): 12. Distribución: México.
15. Chlaenobia pilositarsis (Blackwelder, 1944). Phyllophaga pilositarsis Blackwelder, 1944. United States National Museum Bulletin, 185(2): 225. Distribución: México. (Phyllophaga pilositarsis fue el nombre de reemplazo para Chlaenobia dissimilis, homónimo secundario de Ancylonycha dissimilis Chevrolat, 1865).
16. Chlaenobia rodriguezi Bates, 1889. Chlaenobia rodriguezi Bates, 1889. Biologia Centrali-Americana, 2: 399. Distribución: Guatemala.
17. Chlaenobia rzedowskiana (Aragón y Morón, 2003). Phyllophaga (Chlaenobia) rzedowskiana Aragón y Morón, 2003. Proceedings of the Entomological Society of Washington, 105(3): 560. Distribución: México.
18. Chlaenobia scabripyga Bates, 1888. Chlaenobia scabripyga Bates, 1888. Biologia Centrali-Americana, 2: 167. Distribución: Costa Rica, El Salvador, Guatemala, México.
19. Chlaenobia tumulosa Bates, 1888. Chlaenobia tumulosa Bates, 1888. Biologia Centrali-Americana, 2: 168. Distribución: Belice, El Salvador, Guatemala, Honduras, México, Nicaragua.
20. Chlaenobia vexata (Horn, 1885). Phytalus vexatus Horn, 1885. Transactions of the American Entomological Society, 12: 121. Chlaenobia vexata (Horn); Chapin, 1935. Smithsonian Miscellaneous Collections, 94(9): 13. Distribución: Guatemala, México, Estados Unidos. Phytalus cavifrons Linell, 1896 (Chapin, 1935). Chlaenobia vexata unituberculata Bates, 1889 (Chapin, 1935)
21. Chlaenobia yei (Morón, 1991). Phyllophaga (Chirodines) yei Morón, 1991. Giornale Italiano di Entomologia, 5: 402. Distribución: México.
22. Chlaenobia zunilensis (Bates, 1888). Chirodines zunilensis Bates, 1888. Biologia Centrali-Americana, 2: 169. Distribución: Guatemala, Honduras, México.

