










Services on Demand
Journal

Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
 Cited by SciELO
Cited by SciELO Related links
 Similars in
SciELO
Similars in
SciELO Share

 Permalink
PermalinkRevista mexicana de ciencias geológicas
On-line version ISSN 2007-2902Print version ISSN 1026-8774
Rev. mex. cienc. geol vol.31 n.3 Ciudad de México Dec. 2014
Artículo regular
The alluvial sequence of Santa Cruz Nuevo, Mexico: Paleoecological interpretation of fossil vertebrates and pedosediments from marine isotope stage 3
La secuencia aluvial de Santa Cruz Nuevo, México: Interpretación paleoecológica de vertebrados fosiles y pedosedimentos de la etapa isotópica marina 3
Rosa E. Tovar1*, Sergey Sedov2, Marisol Montellano-Ballesteros2, and Elizabeth Solleiro2
1 Posgrado en Ciencias de la Tierra, Instituto de Geología, Universidad Nacional Autónoma de México, Ciudad Universitaria, Delegación Coyoacán, 04510 Cd. de México, Mexico. *rosae.tovar@ciencias.unam.mx
2 Instituto de Geología, Universidad Nacional Autónoma de México, Ciudad Universitaria, Delegación Coyoacán, 04510 Cd. de México, Mexico.
Manuscript received: June 30, 2014
Corrected manuscript received: October 13, 2014
Manuscript accepted: October 14, 2014
ABSTRACT
The Pleistocene was climatically variable worldwide. In Mexico, climatic changes have been primarily studied in central and northern regions of the country, particularly in regard to marine isotope stage 2 (MIS 2). The paleoenvironments of the previous interglacial, MIS 3, remain less known, especially in the south. In the present work, the results for an alluvial pedosequence in Santa Cruz Nuevo, Puebla are presented; the large and small vertebrates recovered from this locality were used to reconstruct past paleoenvironments. However, it was found that the suite of fossil microvertebrates used here was more informative than that of the macrofossils because they are still extant, thus allowing insights into their ecology.
Radiocarbon dating of the pedosequence indicates it was formed during MIS 3, around 30,000 radiocarbon years before present. Four pedological units were recognized: stagnogleyic, carbonate, gley and humus. With the exception of the carbonate unit, fossils were retrieved from all pedological units. Paleosols varied between Fluvisols and Gleysols, neither of which is diagnostic of any particular climate. Nevertheless, they are both hydromorphic, a condition that could result from intermittent flooding.
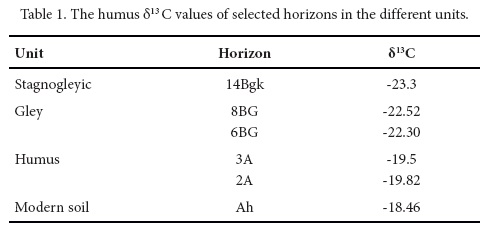
At the base of the section, in the stagnogleyic unit, the presence of mammoth bones in conjunction with negative values of δ13C (-23.3 ‰) in paleosols suggest the existence of grassland with a dominance of C3 grasses. The isotopic values are similar in the succeeding units (-23 to -22 ‰); however, the fossil association found in Santa Cruz Nuevo indicates the incorporation of shrubland faunas. It can thus be inferred that the plant composition in Santa Cruz Nuevo comprised C3 grasses and CAM metabolic succulents such as cacti and magueys. Hydrated CAM plants use the ribulose 1,5-biphosphate (RBP) carboxilase enzyme, which produces negative values comparable to those of C3 plants. The uppermost unit, the humus, has a similar fossil association, but in spite of this, the paleosol-based isotopic values suggest a vegetation mix, indicated by the presence of C4 plants such as grasses.
The comparison of our results with other records suggests that the end of MIS 3 was a dry phase which changed to humid, and in some places cool conditions, during MIS 2. No MIS 2 record is preserved in the Santa Cruz Nuevo pedosequence. The comparison of the modern soil and climate with our pedosequence shows more humid conditions during MIS 3. Finally, and although the microvertebrate associations are similar to extant ones, the macrovertebrates also encompass extinct taxa.
Key words: microvertebrates; alluvial sequence; paleosols; marine isotope stage 3; Puebla; Mexico.
RESUMEN
El Pleistoceno fue un periodo de variados cambios climáticos. En México, los cambios climáticos han sido más estudiados en las porciones centro y norte del país, particularmente los de la etapa isotópica marina 2. Los paleoambientes del periodo glacial previo, la etapa isotópica marina 3, son menos conocidos, especialmente en la porción sur. En este trabajo presentamos los resultados para una pedosecuencia aluvial en Santa Cruz Nuevo, Puebla, donde se han encontrado fósiles de vertebrados, con una gran proporción de microvertebrados asociados a paleosuelos. Los fósiles fueron usados para reconstruir los paleoambientes, sin embargo, los microvertebrados fósiles fueron más útiles debido a que continúan presentes en el área y a través de sus análogos actuales podemos reconocer sus requerimientos ecológicos.
La datación por radiocarbono de la pedosecuencia indica que se formó durante la etapa isotópica marina 3, alrededor de los 30,000 años de radiocarbono antes del Presente. Se reconocieron cuatro unidades pedológicas: estagnogleica, carbonatada, gleyca y húmica. En todas se encontraron fósiles con excepción de la unidad carbonatada. Los paleosuelos varían entre Fluvisoles y Gleysoles, los cuales no son diagnósticos para un clima en particular. Sin embargo, son indicativos de hidromorfismo que pudo ser resultado de inundaciones temporales.
Se usaron datos de δ13C de paleosuelos y las asociaciones de microvertebrados como los registros más importantes para las reconstrucciones paleoambientales. En la base, en la unidad estagnogleyca, la presencia de huesos de mamut y los valores negativos de δ13C (-23.3‰) sugieren la existencia de un pastizal con dominancia de pastos C3. Los valores isotópicos son similares en las siguientes unidades (-23 a -22 ‰), sin embargo, la asociación fósil indica la incorporación de habitantes de matorrales. Con base en ambos registros, inferimos que las plantas correspondieron a pastos C3 y suculentas como los cactus y magueyes que tienen metabolismo CAM. Las plantas CAM, que sí disponen de agua, usan la ribulosa 1,5-bifosfato carboxilasa dando valores negativos como lo hacen las plantas C3. La última unidad, la húmica, tiene una asociación fósil similar pero los valores isotópicos de los paleosuelos sugieren una mezcla en vegetación, indicativa de la incorporación de plantas C4 como los pastos C4.
La comparación de nuestros resultados con otros registros sugiere que el final de la MIS 3 fue seco y que cambió, durante la etapa isotópica marina 2, a condiciones húmedas y templadas en algunas zonas. En la pedosecuencia estudiada no encontramos registro de la etapa isotópica marina 2. La comparación del suelo moderno y clima con respecto a nuestra pedosecuencia muestra condiciones más húmedas durante la etapa isotópica marina 3 y a pesar de que las asociaciones de microvertebrados son similares a las actuales, existen algunos elementos que se extirparon y la megafauna se extintinguió.
Palabras clave: microvertebrados; secuencia aluvial; paleosuelos; etapa isotópica marina 3; Puebla; México.
INTRODUCTION
Paleoecological interpretations, at least for the Phanerozoic and particularly for the Quaternary, are mainly based on the uniformitarianism and actualism principles. Uniformitarianism was proposed by James Hutton (1726–1797) and assumes that the geological processes that currently operate are the same that operated in the past. In other words, "the present is the key to the past". Actualism includes the direct comparison of different sources of information provided from fossils, sediments and paleosols with modern analogs.
During the Pleistocene, there was a rich biotic diversity of mammals in different regions of the world (North and South America, Europe and Australia), and it has been suggested that this biodiversity was richer than at present (Semken et al., 2010). However, the ecological requirements of macrovertebrates are not yet completely understood and paleoecological inferences from these fossils are still ambiguous and uncertain. On the other hand, microvertebrates, with their high reproductive rates and low extinction ratios, are ideal tools for paleoecological reconstructions. Furthermore, when small vertebrate remains are found along with megafauna, they ultimately contribute with valuable information on the paleoecology of the latter.
Although the use of fossil microvertebrates has many approaches in taxonomy, paleobiogeography, paleobiology and evolution (e.g., Sankey and Baszio, 2008), their main application in archaeology and Pleistocene paleontology is in the reconstruction of paleoecosystems. Through an analogy of archaeological and fossil microvertebrate remains it is possible to make paleoecological inferences because, in the modern natural contexts of the same species, they represent the local fauna and have narrow ecological requirements (Stahl, 1996).
The paleoecological analyses of many late Pleistocene faunal communities have established combinations of species which are ecologically incompatible at present. These assemblages are known as disharmonic, non-analog, or intermingled faunal associations (Fay, 1988; Graham, 1986; Graham and Lundelius, 1989). Evidence of disharmonic associations are numerous worldwide (Markova et al., 2002, 2010, Ceballos et al. 2010).
In this context, it can be said that microvertebrate fossil faunas are the key to the paleoecological reconstructions of disharmonic associations. In Mexico, fossil microvertebrates are found in caves (e.g., Arroyo-Cabrales and Polaco, 2003), ancient lakes (e.g., Álvarez and Hernández-Chávez, 1994), ancient springs (e.g., Álvarez and Álvarez-Castañeda, 1991) and ancient rivers (e.g., Van Devender et al., 1985) during the Pleistocene.
Soils developed over floodplains preserve a good record of the environmental conditions, geomorphology and/or local conditions. In alluvial sequences, microvertebrates are mainly found in the terrace sediments accumulated in earlier floodplains. Macromorphological characteristics are one of the many clues soils register in their soil memory. Soil memory is the ability of the soil system to record the environmental phenomena through pedogenetic processes that act in situ (Targulian and Goryachkin, 2004). Other soil memory elements in alluvial soils are humus δ13C, which are remains of organisms or their biotic activity such as feces, channels, phytoliths and magnetic susceptibility.
The use of combined records of soil memory and fossil vertebrates helps to interpret the environmental conditions that prevailed in an area (e.g., Solís-Castillo et al., 2012; Tovar et al., 2013, 2014). However, the recognition of disharmonic associations and the absence of knowledge on megafaunal habitats might be one of the reasons for a limited use of paleofaunas as paleoenvironmental indicators in Mexico. As a matter of fact, Mexican microvertebrate fossil remains are very little known and their use is restricted to taxonomic designation.
The goal of this article is to reconstruct the paleoenvironments of Santa Cruz Nuevo from the late Pleistocene to the present by studying macro and micro fossil vertebrates, paleosols and some elements of their soil memory.
STUDY AREA
The study area is located in the vicinity of the town of Santa Cruz Nuevo, southern Puebla, Mexico (Figure 1). Two different climatic settings are present in Santa Cruz Nuevo. The predominant climate is semi-warm subhumid with summer rains A(C)wO(w); the average annual temperature varies between 22 and 25 °C. The secondary climate, present in the northeast portion of the community, is subhumid temperate with summer rains C(w)O(W), and its average annual temperature varies between 17 and 19 °C. The vegetation is rosetophyl shrubland. A gallery forest borders the Acatlán river.
The Quaternary outcrops overlay the Mixteca terrain, which has been characterized by the Acatlán Complex. The Acatlán Complex comprises igneous and sedimentary rocks ranging from the Paleozoic to the Quaternary (Morán-Zenteno, 1994). The Santa Cruz Nuevo sediments belong to the latter.
In 2006, Tovar and Montellano reported the presence of 22 families, 24 genera and 11 species of fossil vertebrates in the study area.
MATERIAL AND METHODS
Four overlapping profiles (A-D) along the Acatlán river were described and measured. Six pedostratigraphic units were recognized on the basis of their macromorphological features and named according to their predominant pedological or sedimentological characteristics. The description of the soil features was achieved according to Birkeland (1999).
Bulk samples from every unit were taken for physical and chemical analyses, unaltered samples for thin sections, and samples presenting organic matter from selected horizons for radiocarbon dating.
Thin-sections (30 μm) were prepared from undisturbed soil samples, impregnated with the resin Cristal MC-40 at room temperature, studied under a petrographic microscope and described according to Bullock et al. (1985).
Humus and charcoal samples were sent to Beta Analytic Inc. for age estimation through Accelerator Mass Spectrometry (AMS). All dates were reported in radiocarbon ages before present (14C ages BP) and in calibrated ages according to <www.calpal.de>. Stable carbon isotopes, reported as δ13C, were obtained from organic matter in paleosols and sent to Laboratorio Universitario de Geoquímica Isotópica (LUGIS), UNAM.
To establish particle-size distribution quantitatively, sand (2–0.02 mm) was divided in fractions by sieving, while silt (0.02–0.002 mm) and clay (< 0.002 mm) were separated through the hydrometer method (Gee and Or, 2002).
Fossil vertebrates have been collected from the Santa Cruz Nuevo locality since 2001. Microvertebrate remains were recovered by the screen-washing technique, and megafaunal remains were recovered by using gypsum splints or toilet tissue wraps. The fossils collected were assigned to a pedologic unit or layer. All fossil material was prepared in the Laboratorio de Paleontología, Instituto de Geología, UNAM, and ultimately deposited in the Colección Nacional de Paleontología, at the same institute.
RESULTS
Stratigraphy and soil characteristics
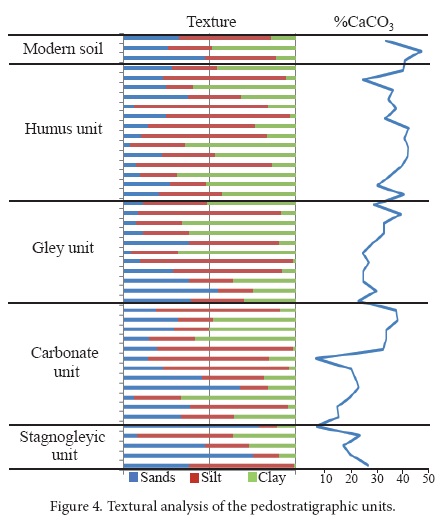
Two pedostratigraphic sections were analyzed and correlated through marker soils. Four pedostratigraphic units were recognized in the study area of Santa Cruz Nuevo, Puebla, Mexico, and were identified as stagnogleyic, carbonate, gley and humus (Figures 2 and 3). Among the different units, layers with cross-stratification, lenses or small channels were observed. Fossils were found in all units, with the exception of the carbonate unit.
According to the stratigraphy, the stagnogleyic unit (2461-2680 cm.) is the oldest fluvial deposit in the study area (Figures 2 and 3). Inside this unit, at least three sedimentary beds with evidence of fluvial processes can be identified. The most recent bed probably includes several other beds because of their thickness. However, this bed could not be studied further due to its inaccessibility. The three sedimentary beds correspond to sands with planar cross-stratification and lenticular channels.
Three paleosols were recognized (14Bgk, 13G, 12Bg) in the stagnogleyic unit by means of their color pattern and pedogenic structure. Moreover, paleosol 14Bgk is separated from paleosol 13G by a sedimentary layer.
In general, paleosols of the stagnogleyic unit are poorly developed and their texture is mainly sandy (Figure 4), with a variable proportion of silt and mud and, a considerable content of gravels. Manganese (Mn) dendrites are found in the entire unit, even within fossils. Paleosols developed over sediments, filling old channels that are still visible at present (Figure 3b). Munsell color varies between 2.5 Y in the first 219 cm and 10 YR in the upper 30 cm.
Paleosol 14Bgk presents carbonate rhizoconcretions (Figure 3c) embedded in a brown matrix with columnar structure and numerous yellow-brown patches and black dendrite-manganese mottles (paleosol 14Bgk). A sand bed with disperse gravels (pebbles and cobbles) and channels with imbricated gravels overlies this paleosol.
Paleosol 13 G is incipient and some of its channels are preserved. Columnar structure is not well developed and is slightly friable. This paleosol contains remains of fossil mammoth (ribs, vertebrae and a defense).
Paleosol 12 Bg presents a high proportion of CaCO3 associated with Mn channels, it is structured in blocks and, as in paleosol 13 G, is slightly friable. Over the stagnogleyic unit lies a 367 cm sandstone bed with numerous channels. However, its position made it challenging to study.
At a micromorphological level, cavities, channels (some with chambers and fillings), manganese and iron (Fe) oxides are common in the stagnogleyic unit (Figure 5a). Little mesofauna evidence was found (Figure 5b).
The carbonate unit (1559–1973 cm) has brown-red colors (Munsell color corresponds to 10YR) with little alluvial evidence. Three poorly differentiated paleosols (11Bw-11Bk-11C, 10Bw-10Bkm, 9BCkg-9Bw) were recognized and separated by sedimentary beds. The 11Bw-11Bk-11C paleosol has a silt-clay texture, with prismatic blocks that can be broken into small blocks. Over this paleosol, the sedimentary bed corresponds to channeled sands.
Paleosol 10Bw displays a clay texture with small pebbles, several yellow-brown patches, CaCO3 in pores, and an angular block structure. Paleosol 10Bkm presents similar characteristics; however, it is more compact due to a higher proportion of CaCO3 and a smaller number of patches. The subsequent layer is a silt layer identified as a weathered ash layer but with no visible structure.
Horizon 9BCkg presents a sand-silt texture with yellow-brown patches, pebbles and subangular blocks. Horizon 9Bw exhibits a silt-clay texture with a structural tendency to form blocks.
At a micromorphological level, high proportions of Mn and Fe oxides are evidenced in the carbonate unit. Although few channels and cavities can be observed, feeding evidence is common.
The gley unit (928–1559 cm) comprises three paleosols (8CG-8BG-, 7BCg-7BG, 6C-6B-6Bg) and a gleyic sediment. This unit is characterized by a toneless brown to greenish color (Figure 3d) and brown-yellow patches. The Munsell color is 10YR except in paleosol 8CG-8BG, in which it is 2.5Y. The Mn dendrites are not as common as in the stagnogleyic unit.
Horizons 8CG-8BG displays a silt-clay texture, being finer in 8BG. The structure in 8CG is organized in blocks, whereas in 8BG it is granular. Horizon 7BCg presents a similar proportion of sands, silt and clay to that of 8CG-8BG. The structure consists of small angular blocks. Horizon 7Bg has a clay-silt texture and consists of small angular blocks than can be broken into granules. The texture of paleosol 6C-6B-6Bg is silt-muddy with an overlying clay layer; its structure is mainly organized in blocks.
This block structure is also evident at a micromorphological level (Figure 5c), in which fractures and Mn dendrites are evident, but not Fe oxides. No evidence of mesofaunal activity is observed, but a plant stem with secondary carbonate was preserved (Figure 5d).
The humus unit (143–828 cm) comprises four paleosols (5C-5B-5AB-5A, 4C-4Bg2-4Bg1, 3C-3BC-3A, 2C-2B-2A), three of which present a superficial Ah horizon. The Ah horizon is approximately 10-15 cm deep (Figure 3e). The structure consists of small blocks with a loamy texture (Figure 4) and scattered gravels. Sedimentary evidence of small channels and lamination can be observed. At the top of the humus unit, lenticular channels and parallel cross-stratification and through cross stratification are present. The Munsell color is 10YR; however a strong pigmentation with dark humus is characteristic of the Ah horizons of this unit (Figure 3e).
The micromorphology evidences the presence of pedofeatures of Fe and Mn oxides, the latter being the least abundant. Unlike in the overlying gley unit, biogenic pores, some of them filled with mesofaunal feces and charcoal, were observed in this unit (Figures 5e and 5f).
The modern soil is poorly developed. The Ah horizon is 10 cm deep and presents a granular structure, a silt texture, and a great number of plant roots. The vegetation is primarily composed of cacti and magueys. Fractures and some pores can be appreciated at a micromorphological level. An absence of the ferruginous pedofeatures and Mn oxides present in all paleosols is noteworthy.
Two radiocarbon ages were obtained from the gley and humus units, 30,380±220 14C years BP (34,512±191 cal. years BP) and 31,680±120 14C years BP (35,610±403 cal. years BP), respectively. Both ages indicate that the deposit was formed at the end of marine isotope stage 3 (MIS 3).
The δ13C values of humus and CaCO3 (Table 1 and 2) are more negative at the base, and become less negative at the top of the pedostratigraphic section.
Fossil findings
Three of the four recognized pedostratigraphic units in Santa Cruz Nuevo yielded fossil vertebrate remains. Mammoth remains (ribs, vertebrae, tusks and other unidentified anatomical parts) have been found in paleosol 3G of the stagnogleyic unit since 2002. Interestingly, fossils from this unit are dark, presumably due to an assimilation of manganese oxide.
The richest fossiliferous unit is the gley unit, where many microvertebrate fossils have been collected (Tovar and Montellano, 2006), particularly from horizon 8CG (Figure 2) and correlative beds. Fossils like salamanders (Ambystoma sp., Plethodontidae), toads (Incilius sp. and Incilius occidentalis), frogs (Rana sp.), turtles (Gopherus sp. cf. G. berlandieri), lizards (Sceloporus sp. cf. S. grammicus, Sceloporus sp. cf. S. horridus, Scincidae), snakes (Conopsis sp., Lampropeltis sp., Leptodeira sp., Senticolis sp., Thamnophis sp.), rabbits (Sylvilagus floridanus) and mice (Neotoma mexicana, Peromyscus difficilis) were commonly found in lenses throughout this unit.
A fossil horse tibia was collected from a channel in horizon 6Bg, at the top of the gley unit. However, an accurate dating could not be obtained due to low collagen content. At the same level but in lateral outcrops, several remains of fossil toads (Incilius sp. and Incilius occidentalis), spade foot toads (Scaphiopodidae), turtles (Kinosternon sp.), snakes (Conopsis sp., Lampropeltis sp., Trimorphodon sp., Crotalus sp.), giant armadillos (Glyptotherium sp.), rabbits (Sylvilagus floridanus), mice (Neotoma mexicana, Reithrondomys sp.), bears (Ursidae), cacomistles (Bassariscus sp. cf. B. sumichrasti), horses (Equus conversidens) and mammoths (Mammuthus sp.) have been recovered.
Toad (Incilius sp., Incilius occidentalis), turtle (Kinosternon sp.), snake (Lampropeltis sp., Thamnophis sp., Crotalus sp.), bird (Cyrtonyx sp., Catharus sp. cf. C. occidentalis, Columba fasciata, Icterus sp., Pipilo sp., Salpinctes sp. cf. S. obsoletus, Dumetella sp. cf. D. carolinensis, Campylorhynchus sp. cf. C. rufinucha, Callipepla sp. cf. C. squamata), giant armadillo (Glyptotherium sp.), rabbit (Sylvilagus floridanus), mice (Neotoma mexicana) and deer (Odocoileus sp.) remains were collected from lenses in the sedimentary beds of the humus unit (Figure 2).
Considering all four units together, microvertebrates were more abundant than macrovertebrates: the number of megafaunal elements represents only 13% of the fossil bones recovered. The scarce megafaunal taxa recovered from the Santa Cruz Nuevo locality correspond to the Orders Cingulata, Artiodactyla, Perissodacctyla, Proboscidea and the Ursidae family.
Most of the megafaunal bones are fragmented and therefore, non diagnostic. The most abundant elements are isolated osteoderms, antlers and teeth. The most complete remains include jaws and a fragmentary horse skull (Figure 6).
On the other hand, microvertebrate fossil bones are better preserved; a great number of limb bones are complete. However, the most abundant microvertebrate elements correspond to vertebrae. In addition, incomplete jaws with dental series and isolated teeth have also been found.
Finally, a turtle carapace scute with gnawing marks (Figure 7a) and root marks on frog bones (Figure 7b) are some of the most remarkable evidence of biological disturbance on the fossil remains of Santa Cruz Nuevo.
DISCUSSION
Paleosols and the environmental setting
The paleosols of Santa Cruz Nuevo provide evidence of incipient pedogenesis and sedimentary structures such as lenses or stratification. These sedimentary structures are common in alluvial soils, mainly in soils developed in floodplains (Gerrard, 1992).
The Mn dendrites, mainly found in the stagnogleyic unit but present in all units, are supporting evidence of temporary water saturation, probably originated from flooding (Gerrard, 1992; Schaetzl and Anderson, 2005). The poor development of paleosols, the presence of alluvial sedimentary structures (e.g., planar cross stratification and channels, combined with Mn dendritic mottles and gleyic color pattern) indicate that the paleosols of the stagnogleyic unit correspond to Gleyic Fluvisols (WRB, 2006).
Paleosols of the gley unit are Fluvisols and Gleysols. However, the exposition to floods appears to have been prolonged because no macroscopic evidence of Mn, a more mobile element than Fe (Gerrard, 1992; Schaetzl and Anderson, 2005), was found.
The incipient extent of differentiation in the carbonate unit suggests that these paleosols are Cambisols. On the other hand, the humus unit maintains theirs superficial A horizons suggesting a stability in the landscape. At the top of this unit, the presence of several small channels indicates the return to a fluvial activity. Above this unit there is a gap with no MIS 2 sediments preserved. The modern soil corresponds to Cambisol (WRB, 2006) originated in a xeric climate. It must be stressed that all paleosols show redoximorphic features, which are indicative of temporary water saturation. This kind of features is absent in the modern soil. Such absence points to a less drained environment or more frequent floods in the Pleistocene compared to the Holocene.
Fluvisols are not diagnostic of a particular climate (WRB, 2006) and composition of the vegetation was impossible to be determined because of an absence of phytoliths and pollen. However, past environments can also be reconstructed by means of stable carbon isotopes (e.g., Lounejeva-Baturina et al., 2006). In plants, carbon isotopic fractionation is associated with photosynthesis. Higher plants have three different metabolic pathways: C3, C4 and CAM. Values of δ13C for C3 plants fall between -25 and -35‰, those for C4 range from -10 to -17‰, and CAM plants have a range of δ13C values between those typical of C3 and C4 plants because they are facultative. CAM plants have values similar to those of C4 plants during active acidification, i.e., when plants are not well watered (Edwards and Walker, 1983). C3 plants are typical of humid climates, whereas C4 plants characterize warm and dry areas.
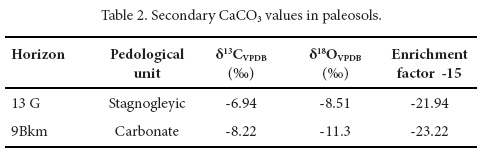
The analysis of δ13C in paleosol humus and secondary carbonates shows a trend to less negative values at the top of the pedosequence (Tables 1 and 2), with values ranging between -22‰ and -23‰ in the lower three units (stagnogleyic, gley and carbonate) to -19.82‰ and -19.5‰ in the humus unit. The δ13C of modern soil is -18.46. Thus, we observe a tendency towards less negative δ13C values in the modern soil compared to Pleistocene paleosols. In Santa Cruz Nuevo, this tendency towards drier environments is less conspicuous than in other Pleistocene-Holocene sequences of central and southern Mexico (e.g., Canul-Montañez, 2008, Lounejeva-Baturina et al., 2006, Lozano-García et al., 2005, Tovar et al., 2013, 2014).
The δ13C values correspond to a mixed vegetation with a preponderance of C3 plants at the base (stagnogleyic unit) and an increase in C4 plants at the top (humus unit). At present, modern soil in Santa Cruz Nuevo supports a shrubland vegetation with numerous CAM plants such as cacti and magueys with no evidence of grasses, except for the cultivated areas.
Fossil vertebrates of Santa Cruz Nuevo
The fossil vertebrates of Santa Cruz Nuevo were highly useful in the reconstruction of past environments in the study area because they presented clues to the conditions in which the organisms lived, their position in the food chain and their interaction with other species. In addition, the analysis of skeletal elements traces the history of the individual from its death, transport, burial, and finally, the discovery of the site where the remains lay (Behrensmeyer, 1984).
The paleoenvironment reconstruction in the study area was developed according to the ecological requirements of modern analogs and the published data on habitat reconstructions of extinct fauna. Fossil vertebrates were found in three paleopedological units (stagnogleyic, gley and humus units) and were more abundant in the last two.
Bone elements of Mammuthus sp. most likely belonging to a single specimen (since 2002, different parts of the skeleton have been discovered) were uncovered in the stagnogleyic unit. These remains have not yet been removed from the site due to extraction difficulties. The animal probably died close to the study area, because most of the skeletal elements remain together, although not articulated. Agenbroad (1984) suggested that the preferred habitat of mammoths was open areas as grasslands. The present work suggests that the grassland composition had a considerable proportion of C3 grasses as evidenced by a δ13C signature.
In the gley and humus units, fossils are associated to fluvial sediments or incipient paleosols. The most abundant orders correspond to animals of amphibian habits or those living close to rivers.
Behrensmeyer (1982) stated that fossils found inside channels can be interpreted as allochtonous (outside the animal distribution area), whereas fossils found in a finely grained matrix characteristic of floodplains are inferred as autochthonous (within the area where the animal lived). Because microvertebrates have been found in fine sediments and fragile bones break easily during water transportation (Korth, 1979), it is assumed that the microvertebrate association in Santa Cruz Nuevo is para-autochthonous, i.e., within the animal distribution area. As previously mentioned, megafaunal remains are less abundant than those of microvertebrates, with only four mammal orders present (Xenarthra, Uranotheria, Perissodactyla and Carnivora). Most of the macrovertebrate fossils correspond to skull fragments, jaws and isolated teeth, which, according to Voorhies (1969), are not transported for long distances.
A large sample of microvertebrates was collected from the gley unit (Figure 8). Some of the identified taxa, Ambystoma sp. and Kinosternon sp. are strictly associated with fresh water (Zug et al., 2001); other taxa such as Scaphiopodidae, Gopherus sp. cf. G. berlandieri, Sceloporus sp. cf. S. horridus, Senticolis sp., Peromyscus difficilis, Equus conversidens and Mammuthus sp. are restricted to dry areas in ecosystems such as shrublands, dry spiny forests, grasslands or deserts (Smith, 1939; Hibbard and Taylor, 1960; Ceballos-González and Galindo-Leal, 1984; Benítez-Gálvez, 1997; Agenbroad, 1984); and others like Incilius occidentalis, Sceloporus sp. cf. S. grammicus, Conopsis sp., Leptodeira sp., Neotoma mexicana, Bassariscus sp. cf. B. sumichrasti inhabit dry or temperate areas. Dry ecosystems include shrublands, spiny forests and grasslands; temperate ecosystems are coniferous and/or oak forests (Duellman, 1966; Ceballos-González and Galindo-Leal, 1984; Benítez-Gálvez, 1997). Some eurytopic taxa are present in the sample: Lampropeltis sp., Thamnophis sp., Crotalus sp., Sylvilagus floridanus, Reithrondomys sp. (Lee, 1996; Benítez-Gálvez, 1997). Due to their high tolerance to wide ecological variations, eurytopic taxa are not useful in environment determination.
According to McNab (1985), Glyptotherium, the giant armadillo, inhabited tropical to warm-temperate areas, eating leaves along water bodies (Gillete and Ray, 1981). However, the δ13C analysis of glyptodont teeth from El Cedral, San Luis Potosí, Mexico, exhibited a mixed diet (C3/C4) of the members of this taxon. The large C4 plant consumption of the El Cedral Glyptotherium suggests that this genus was, preferentially, a grassland grazer (Pérez-Crespo et al., 2012). The study of the fossil giant armadillos of Santa Cruz Nuevo supports the interpretation of Pérez-Crespo et al., (2012).
The humus unit shares many species with the gley unit (Figure 8). The species or genera inhabiting dry spiny forests, shrublands and/or grasslands are Salpinctes sp. cf. S. obsoletus, Dumetella sp. cf. D. carolinensis, Campylorhyncus sp. cf. C. rufinucha, Callipepla sp. cf. C. squamata, and Glyptotherium sp. (Peterson and Chalif, 2008; Pérez-Crespo et al., 2012). The temperate species are Catharus sp. cf. C. occidentalis and Columba fascista (Peterson and Chalif, 2008). The species distributed in dry and temperate areas are Incilius occidentalis, Cyrtonyx sp., Neotoma mexicana and Odocoileus sp. (Duellman, 1966; Ceballos-González and Galindo-Leal, 1984; Peterson and Chalif, 2008). The eurytopic species are Lampropeltis sp., Thamnophis sp., Crotalus sp., Icterus sp., Pipilo sp., Sylvilagus floridanus (Benítez-Gálvez, 1997; Peterson and Chalif, 2008). In the humus unit only one aquatic genus (Kinosternon sp.) is found.
The assemblages of both gley and humus units present similar fauna. Although some temperate forest elements are present in both groupings, most of the identified taxa inhabited dry areas such as xeric shrublands or grasslands, suggesting that these kinds of vegetation were predominant during the end of the Pleistocene, although the idea of a mosaic habitat is not dismissed. With the use of both fossil and recent microvertebrates it was noticed that there are no extinct microvertebrates in the area and that the eradicated fauna belong to herpetofauna (Tovar and Montellano, 2006). Salamanders (Families Ambystomatidae and Phletodontidae), skinks (Family Scincidae) and two kinds of turtles (Families Bataguridae and Testudinidae) are absent in the study area at present.
Although several fossil taxa have been eradicated from the Santa Cruz Nuevo area, it is worth mentioning that most of the microvertebrate fossils in particular, still have extant representatives in the study area, and that this pattern has also been observed in other localities (Parmley and Pfau, 1997).
The results herein presented suggest that the stagnogleyic unit (δ13C -23.3‰ to - 21.94‰) corresponds to a grassland with an abundance of C3 grasses and a minor proportion of C4 grasses and other C4 plants. Both kinds of grasses might have represented a food source for the mammoth, which has been reported as a mixed-feeder (Pérez-Crespo et al., 2009; Nunez et al., 2010).
The gley unit (30,380±220 14C years BP) presents δ13C values (-22.52‰ to - 22-3‰) similar to those of the stagnogleyic unit. The occurrence of certain taxa suggests the possible presence of either grassland or shrubland in the area. The evidence assembled here leads to interpret the paleoenvironment as a grassland area, with a great proportion of C3 grasses and patches of shrubland, in which the dominant vegetation was composed by CAM plants such as cacti, magueys and other succulent plants. CAM plants are particularly adapted to semi-arid environments and when the water supply is abundant, they directly fix atmospheric CO2 during the day through ribulose 1,5-biphosphate (RBP) carboxilase as C3 plants do (Edwards and Walker, 1983).
Water may not have been an abundant resource in the area; however, it is well documented that flooded water can generate a temporary wetland environment in lowlands, regardless the climate. Gleysols, soils characteristic of the gley unit, are formed in nearly all climates ranging from perhumid to arid and are associated with topographic lows (WRB, 2006) where water is available and exhibits a negative signal.
The humus unit (31,680±120 14C years BP) shows higher δ13C values (-19.82‰ to -19.5‰), supporting an increase of C4 plants and probably warmer conditions alongside a decrease in water supply.
Chronology of Santa Cruz Nuevo and correlation with other records at the end of MIS 3 in south-central Mexico
The pedosequence of Santa Cruz Nuevo comprises the end of MIS 3, around 30,000 14C years BP. The analysis of the environmental conditions, as previously mentioned, suggested a transition from dry to drier conditions.
Santa Cruz Nuevo is located close to the locality of Axamilpa, Tepexi de Rodríguez, Puebla, Mexico. Axamilpa is younger than Santa Cruz Nuevo and the prevailing environmental conditions, at the end of MIS 3, was a temperate weather (a forest) changing to a semi-arid climate (Tovar et al., 2014) as observed at present, demonstrating a dry phase in the region.
However, most publications, either on lakes or paleosol profiles, have previously suggested in their climatic interpretations the presence of dry periods around >30,000 years BP (Figure 9, Table 3). According to δ18O and δ13C values in snail shells, before 36,000 14C BP the paleoclimate in the Upper Barranca Caulapan in Valsequillo was slightly drier than at present. After that date, weather conditions became drier up to the 28,000 to 20,000 14C years BP interval, when conditions became cooler and wetter (Stevens et al., 2012). Lozano-García and Xelhuantzi-López (1997) suggested a subhumid cooler climate with drier periods during the interval of ca. 35,000 to ca. 20,000 14C years BP in the Texcoco Lake.
The analyses of paleosols, δ13C, and fossil pollen from the subunit TX2 (30,189 to 51,798 cal. years BP) in Huexoyucan and the sections Tlalpan and Mammut (38,000 to 29,000 14C years BP), in Tlaxcala, Mexico, suggest the presence of a forest vegetation with a mean annual temperature of 7-8 °C, with cold and humid conditions (Sedov et al. 2009; Solís-Castillo et al., 2012). In the Teotihuacan Valley, Estado de Mexico, the pedostratigraphic level SP4 is correlated with TX2. There, the pedogenesis was humid, indicating a temperate forest environment. However, there is also evidence of dry seasons such as vertic processes, carbonate precipitation and bioturbation (Solleiro-Rebolledo et al., 2011).
At approximately ~52,000 cal. years BP, a forest of Quercus, Alnus, Cupressaceae and Fraxinus was established in the Zacapu basin, Michoacán, Mexico; however, around 42,000 cal. years BP and until a hiatus around 30,000 years cal. BP, drier conditions took place according to the reported presence of high percentages of Poaceae, Asteraceae, Amaranthaceae and Cyperaceae (Correa-Metrio et al., 2012).
For the Chalco lake, Estado de Mexico, during 34,000 and 31,500 14C years BP, the diatom assemblage suggests that the lake was shallow and alkaline. After 31,500 14C years BP, the lake experienced a reduction in its water level becoming alkaline and saline (Caballero-Miranda, 1997). While for the Cuitzeo lake, Michoacán, the absence of diatoms in the sedimentary record at around 35,000 years BP suggests a very low water level in the lake (Israde-Alcántara et al., 2002).
In the Zacango sequence, northern flank of Nevado de Toluca, Mexico, the pollen record suggests that after the 37,000 years BP eruption of the Nevado de Toluca volcano, there was a shallow and stagnic pond surrounded by grassland (Caballero et al., 2001). However, the pedological features of pedocomplex PT3 in Nevado de Toluca, dated at 35,650 ± 1200 14C years BP, indicate processes typical of Andosols which are formed under forest vegetation (Sedov et al., 2001).
Most MIS 3 records come from lakes and paleosols. Pleistocene vertebrate fossils were very abundant in Mexico however the lack of instrumental dating and stratigraphic control for many localities make the chronological comparison between localities of the same age practically impossible, particularly when we try to compare them in a millennial scale.
On the other hand, lake records are especially useful because their continuous entry allows researchers to recognize regional climatic changes in a known scale, while paleosols are mere glimpses of specific moments in time. However, the study of paleosols also gives clues on local processes and help in the understanding of the dynamics of local communities.
ACKNOWLEDGMENTS
This work was supported by PAPIIT-UNAM, grants No. IN117709, IN110710 and IN108714 (Paleosuelos asociados a registros paleontológicos y arqueológicos: desarrollo de proxies paleoambientales integrales) and CONACYT- 166878 (E. Solleiro-Rebolledo). Authors acknowledge the support of Eligio Jiménez and Jaime Díaz for the preparation of thin sections, René Alcalá for his support in the grain size determination of paleosols and Kumiko Shimada for her help in TOC determination. Pedro Morales and Edith Cienfuegos (Laboratorio de Espectrometría de Isótopos Estables [LUGIS], Universidad Nacional Autónoma de México) contributed with the stable carbon isotope composition of paleosol humus.
We thank Pilar Melgarejo for her help in the language correction and Maria Bronnikova for the revision of the work.
REFERENCES
Agenbroad, L., 1984, New World mammoth distribution, in Martin, P.S., Klein, R.G. (eds.), Quaternary extinction: Tucson, The University of Arizona Press, 90-108. [ Links ]
Álvarez, T., Álvarez-Castañeda, S.T., 1991, Análisis de la fauna de roedores del área de El Cedral, San Luis Potosí, México: Anales del Instituto de Biología, Universidad Nacional Autónoma de México, Serie Zoología, 62, 169-180. [ Links ]
Álvarez, T., Hernández-Chávez, J.J., 1994, Estudio de los restos de Neotoma (Rodentia: Cricetidae) del pleistoceno-holoceno de Tlapacoya, Estado de México, México, con descripción de dos nuevos taxa: Revista de la Sociedad Mexicana de Paleontología, 7(2), 1-11. [ Links ]
Arroyo-Cabrales, J., Polaco, O. 2003. Caves and the Pleistocene vertebrate paleontology of Mexico, in Schubert, B.W., Mead, J.I., Graham, R.W. (eds.), Ice Age Cave Faunas of North America: U.S.A., Indiana University Press, 273-291. [ Links ]
Behrensmeyer, A.K., 1982, Time resolution in fluvial vertebrate assemblages: Paleobiology, 8, 211-227. [ Links ]
Behrensmeyer, A.K., 1984, Taphonomy and the fossil record: American Anthropologist, 72, 558-566. [ Links ]
Benítez-Gálvez, J.E., 1997, Los ofidios de Puebla: México, Fotograbado ETA, S.A. de C.V., 122 pp. [ Links ]
Birkeland, P.W., 1999, Soils and Geomorphology: New York, Oxford University Press, 430 pp. [ Links ]
Bullock, P., Fedoroff, N., Jongerius, A., Stoops, G., Tursina, T., Babel, U., 1985, Handbook for Soil Thin Section Description: Wolverhampton, Waine Research Publications, 152 pp. [ Links ]
Caballero, M., Macías, J.L., Lozano-García, S., Urrutia-Fucugauchi, J., Castañeda, B.R., 2001, Late Pleistocene/Holocene volcanic stratigraphy and paleoenvironments of the upper Lerma basin, Mexico: Special Publications of the International Association of Sedimentologists, 30, 247-261. [ Links ]
Caballero-Miranda, M., 1997, Reconstrucción paleolimnológica del Lago de Chalco durante el último máximo glaciar, el registro de diatomeas entre 34,000 y 15,000 años A.P.: Revista Mexicana de Ciencias Geológicas, 14, 91-100. [ Links ]
Canul-Montañez, M., 2008, Reconstrucción paleoclimática (Cuaternario tardío) de la porción occidental del Valle de Tehuacán, Puebla, México: Estudio palinológico: México, Universidad Nacional Autónoma de México, M.Sc, Thesis, 79 pp. [ Links ]
Ceballos, G., Arroyo-Cabrales, J., Ponce, E., 2010, Effects of Pleistocene environmental changes on the distribution and community structure of the mammalian fauna of Mexico: Quaternary Research; 73, 464-473. [ Links ]
Ceballos-González, G., Galindo-Leal, C., 1984, Mamíferos silvestres de la cuenca de México: México, Limusa, 299 pp. [ Links ]
Correa-Metrio, A., Lozano-García, S., Xelhuantzi-López, S., Sosa-Nájera, S., Metcalfe, S.E., 2012, Vegetation in western Central Mexico during the last 50 000 years: modern analogs and climate in the Zacapu Basin: Journal of Quaternary Science, 27, 509-518. [ Links ]
Duellman, W.E., 1966, The Central American Herpetofauna: An ecological perspective: Copeia, 1966(4), 700-719. [ Links ]
Edwards, G., Walker, D.A., 1983, C3, C4: Mechanisms, and Cellular and Enviromental Regulation of Photosynthesis: Great Britain, Blackwell Scientific Publications, 552 pp. [ Links ]
Fay, L.P., 1988, Late Wisconsinan Appalachian herpetofaunas: relative stability in the midst of change: Annals of Carnegie Museum, 57(9), 189-220. [ Links ]
Gee, G.W., Or, D., 2002, Particle-Size Analysis, in Dane, A.H., Topp, G.C. (eds.), Methods of Soil Analysis, Part 4, Physical Methods: U.S.A., Soil Science Society of America, USA, 255-293. [ Links ]
Gerrard, J., 1992, Soil geomorphology. An integration of pedology and geomorphology: London, Chapman & Hall, 288 pp. [ Links ]
Gillete, D.D., Ray, C.E., 1981, Glyptodonts of North America: Smithsonian Contributions to Paleobiology, 40, 1-255. [ Links ]
Graham, R.W., 1986, Response of mammalian communities to environmental changes during the Late Quaternary, in Diamond, J., Case, T.J. (eds.), Community ecology: U.S.A., Harper and Row publishers, 300-313. [ Links ]
Graham, R.W., Lundelius, E.L., 1989, Coevolutionary disequilibrium and Pleistocene extinctions, in Martin, P.S., Klein, R.G. (eds.), Quaternary extinctions: Tucson, The University of Arizona Press, 223-249. [ Links ]
Hibbard, C.W., Taylor, D.W., 1960, Two late pleistocene faunas from southwestern Kansas: The University of Michigan, Contributions from the Museum of Paleontology, 16(1), 1-223. [ Links ]
Israde-Alcántara, I., Garduño-Monroy, V.H., Ortega-Murillo, R., 2002, Paleoambiente lacustre del Cuaternario Tardío en el centro del lago de Cuitzeo: Hidrobiológica, 12(1), 61-78. [ Links ]
Korth, W.W., 1979, Taphonomy of microvertebrate fossil assemblages: Annals of Carnegie Museum, 48(15), 235-285. [ Links ]
Lee, J.C., 1996, The amphibians and reptiles of the Yucatán Peninsula: U.S.A., Cornell University Press, 500 pp. [ Links ]
Lounejeva-Baturina, E., Morales-Puente, P., Cabadas-Báez, H.V., Cienfuegos-Alvarado, E., Sedov, S., Vallejo-Gómez, E., Solleiro-Rebolledo, E., 2006, Late Pleistocene to Holocene environmental changes from δ13C determinations in soils at Teotihuacan, Mexico: Geofísica Internacional, 45, 85-98. [ Links ]
Lozano-García, S., Xelhuanzti-López, .M.S., 1997, Some problems in the late Quaternary pollen records of central Mexico: Basin of Mexico and Zacapu: Quaternary International, 43-44, 117-123. [ Links ]
Lozano-García, S., Sosa-Nájera, S., Sugiura, Y., Caballero, M., 2005, 23,000 yr of vegetation history of the Upper Lerma, a tropical high-altitude basin in Central Mexico: Quaternary Research, 64, 70-82. [ Links ]
Markova, A.K., Simakova, A.N., Puzachenko, A.Y., Kitaev, L.M., 2002, Environments of the Russian Plain during the Middle Valdai Briansk Interstade (33,000–24,000 yr B.P.) Indicated by Fossil Mammals and Plants: Quaternary Research, 57, 391-400. [ Links ]
Markova, A.K., Puzachenko, A.Y., Kolfschoten, T., 2010, The North Eurasian mammal assemblages during the end of MIS 3 (Brianskian–Late Karginian–Denekamp Interstadial): Quaternary International, 212, 149-158. [ Links ]
McNab, B., 1985, Energetics, population biology, and distribution of Xenarthrans, living and extinct, in Montgomery, G.G. (ed.), The Evolution and Ecology of Armadillos, Sloths, and Vermilinguas: Washington, D.C., Smithsonian Institution Press, 219-232. [ Links ]
Morán-Zenteno, D., 1994, The Geology of the Mexican Republic: U.S.A., The American Association of Petroleum Geologists, 160 pp. [ Links ]
Nunez, E.E., MacFadden, B.J., Mead, J.I., Baez, A., 2010, Ancient forests and grasslands in the desert: Diet and habitat of Late Pleistocene mammals from Northcentral Sonora, Mexico: Palaeogeography, Palaeoclimatology, Palaeoecology, 297, 391-400. [ Links ]
Parmley, D., Pfau, R.S., 1997, Amphibians and reptiles of the late Pleistocene Tonk Creek local fauna, Stonewall County, Texas: The Texas Journal of Science, 49, 151-158. [ Links ]
Pérez-Crespo, V.A., Sánchez-Chillón, B., Arroyo-Cabrales, J., Alberdi, M.T., Polaco, O.J., Santos-Moreno, A., Benammi, M., Morales-Puente, P., Cienfuegos-Alvarado, E., 2009, La dieta y el hábitat del mamut y los caballos del Pleistoceno tardío de El Cedral con base en isótopos estables (δ13C, δ18O): Revista Mexicana de Ciencias Geológicas, 26(2), 347-355. [ Links ]
Pérez-Crespo, V.A., Arroyo-Cabrales, J., Alva-Valdivia, L.M., Morales-Puente, P., Cienfuegos-Alvarado, E., 2012, Diet and habitat definitions for Mexican glyptodonts from Cedral (San Luis Potosí, Mexico) based on stable isotope analysis: Geological Magazine, 149(1), 153-157. [ Links ]
Peterson, R.T., Chalif, E.L., 2008, Aves de México, Guía de campo: México, Editorial Diana, 473 pp. [ Links ]
Sankey, J.T., Baszio, S. (eds.), 2008, Vertebrate microfossil assemblages their role in paleoecology and paleobiogeography: U.S.A., Indiana University Press, 278 pp. [ Links ]
Schaetzl, R., Anderson, S., 2005, Soils. Genesis and Geomorphology: New York, Cambridge University Press, 827 pp. [ Links ]
Sedov, S., Solleiro-Rebolledo, E., Terhorst, B., Solé, J., Flores-Delgadillo, M., Werner, G., Poetsch, T., 2009, Paleosol sequence in Tlaxcala Basin: a multiscale proxy of the Middle to Late Quaternary environmental change in Central Mexico: Revista Mexicana de Ciencias Geológicas, 26(2), 448-465. [ Links ]
Sedov, S., Solleiro-Rebolledo, E., Gama-Castro, J., Vallejo-Gómez, E., González-Velásquez, A., 2001, Buried palaeosols of the Nevado de Toluca: an alternative record of Late Quaternary environmental change in central Mexico: Journal of Quaternary Science, 16(4): 375-389. [ Links ]
Semken, H.A., Graham, R.W., Stafford, T.W., 2010, AMS C-14 analysis of Late Pleistocene non-analog faunal components from 21 cave deposits in southeastern North America: Quaternary International, 217, 240-255. [ Links ]
Smith, H.M., 1939, The Mexican and Central American lizards of the genus Sceloporus: U.S.A., Field Museum of Natural History 26, 397 pp. [ Links ]
Solís-Castillo, B., Solleiro-Rebolledo, E., Sedov, S., Salcido-Berkovich, C., 2012, Paleosuelos en secuencias coluvio-aluviales del Pleistoceno-Holoceno en Tlaxcala: registros paleoambientales del poblamiento temprano en el centro de México: Boletín de la Sociedad Geológica Mexicana, 64(1), 91-108. [ Links ]
Solleiro-Rebolledo, E., Sycheva, S., Sedov, S., McClung de Tapia, E., Rivera-Uria, Y., Salcido-Berkovich, C., Kuznetsova, A., 2011, Fluvial processes and paleopedogenesis in the Teotihuacan Valley, Mexico: Responses to late Quaternary environmental changes: Quaternary International, 233(1), 40-52. [ Links ]
Stahl, P.W., 1996, The recovery and interpretation of microvertebrate bone assemblages from archaeological contexts: Journal of Archaeological Method and Theory, 3, 31-75. [ Links ]
Stevens, R.E., Metcalfe, S.E., Leng, M.J., Lamb, A.L., Sloane, H.J., Naranjo, E., González, S., 2012, Reconstruction of late Pleistocene climate in the Valsequillo Basin (Central Mexico) through isotopic analysis of terrestrial and freshwater snails: Palaeogeography, Palaeoclimatology, Palaeoecology, 319-320, 16-27. [ Links ]
Targulian, V.O., Goryachkin, S.V., 2004, Soil memory: types of record, carriers, hierarchy and Diversity: Revista Mexicana de Ciencias Geológicas; 21(1), 1-8. [ Links ]
Tovar, R.E., Montellano, M., 2006, Pleistocene microvertebrates from Santa Cruz Nuevo, Puebla, Mexico: Current Research in the Pleistocene, 23, 72-74. [ Links ]
Tovar, R.E., Sedov, S., Solís, B., Solleiro, E., 2013, Dark humic alluvial paleosols in Central and Southern Mexico: micromorphological indicators of Late Pleistocene megafauna habitats: Spanish Journal of Soil Science, 3, 217-235. [ Links ]
Tovar, R.E., Sedov, S., Montellano-Ballesteros, M., Solleiro, E., Bennami, M., 2014, Paleosols, bones, phytoliths, and δ13C signatures of humus and teeth in the alluvial sequence of Axamilpa, Puebla: Inferences for landscape evolution and megafauna paleoecology during MIS 3–2 in Southern Mexico: Catena, 112, 25-37. [ Links ]
Van Devender, T.R., Rea, A.M., Smith, M.L., 1985, The Sangamon interglacial vertebrate fauna from Rancho La Brisca, Sonora, Mexico: Transactions of the San Diego Society of Natural History, 21(2), 23-55. [ Links ]
Voorhies, M., 1969, Taphonomy and population dynamics of an early Pliocene vertebrate fauna, Knox County, Nebraska: Contributions to Geology Special Paper No. 1, 1-69. [ Links ]
WRB, 2006, World reference base for soil resources 2006. A framework for international classification, correlation and communication: Rome, Food and Agriculture Organization of the United Nations, 128 pp. [ Links ]
Zug, G.R., Vitt, L.J., Caldwell, J.P., 2001, Herpetology. 2nd edition: U.S.A., Academic Press, 630 pp. [ Links ]

