











 text new page (beta)
text new page (beta) Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
Permalink
Introducción
Las cactáceas tienen características biológicas y ecológicas que las hacen vulnerables a diversos factores de perturbación natural y antropogénica, se presentan en áreas de distribución restringida, y están limitadas a ambientes muy específicos. La familia Cactaceae comprende aproximadamente 2000 especies, y en México se registran alrededor de 850 en 55 géneros (Rzedowski, 1978; Anderson, 2001; Guzmán et al., 2003), por lo que es considerado el centro más importante de diversificación de cactáceas (Bravo-Hollis y Sánchez-Mejorada, 1991). Hernández y Godínez (1994) calculan que el grado de endemismo de las especies es del orden de 78% y de género, de 73%. Los miembros de esta familia ocupan el primer lugar en la lista de la Convención sobre Comercio Internacional de Especies Amenazadas de Fauna y Flora Silvestres (CITES, 2011) y en la International Union for Conservation of Nature (IUCN, 2016). Aproximadamente 260 especies de cactáceas mexicanas están en la Lista Roja (Sánchez-Mejorada, 1987 a, b; Hunt, 1999; Martínez-Ávalos, 2007; IUCN, 2016), 44 de éstas (si bien en realidad son 40, ya que 4 son sinónimos) están en el Apéndice I y toda la fa-milia está en el Apéndice II de CITES (Guzmán et al., 2003; Arias et al., 2005; CITES, 2011; 2016). En el listado de la Norma Oficial Mexicana se registran 260 taxa debido a que es una de las familias que más se detecta en el comercio nacional e internacional con origen silvestre (Glass, 1998; Arias et al., 2005; SEMARNAT, 2010).
Entre los factores reportados que hacen a las cactáceas vulnerables a diferentes tipos de perturbación humana destacan los siguientes:
Tienen tasas anuales de crecimiento individual y poblacional muy lentas (Jordan y Nobel, 1982; Zavala-Hurtado y Díaz-Solís, 1995; Suzán et al., 1998; Mandujano et al., 2001; Jiménez-Sierra et al., 2007).
Presentan patrones de distribución agregados relacionados con la asociación de nodricismo con rocas y/o plantas que proveen condiciones microclimáticas adecuadas para el establecimiento y la supervivencia de las plántulas (Turner et al., 1969; Jordan y Nobel, 1982; McAuliffe, 1984; Nobel, 1989; Suzán et al., 1994; 1998; Mandujano et al., 1998; Leirana y Parra, 1999), por lo que sin la protección de una cubierta vegetal o de una roca adecuada no se observa reclutamiento.
El tamaño y la estructura de tallas de las poblaciones tienden a fluctuar marcadamente a través del tiempo y los juveniles son escasos (Martínez-Ávalos et al., 1993, Mandujano et al., 2001).
La fragmentación del hábitat dada por actividades agrícolas, desarrollos urbanos, pecuarios y turísticos (Martínez-Ávalos et al., 1993; 1994; Zavala-Hurtado y Díaz-Solís, 1995; Suzán, 1997; Brys et al., 2005).
Marcada mortalidad de las plantas propiciada por el pastoreo por ganado caprino, vacuno y equino (Jordan y Nobel, 1982; Suzán, 1997; Huerta y Escobar, 1998; Jiménez-Sierra et al., 2007).
Sobrecolecta de ejemplares como recurso alimenticio para poblaciones humanas y por la colecta ilícita como plantas de ornato (Huerta y Escobar, 1998; Mandujano et al., 1998; Robbins 2003; Jiménez-Sierra et al., 2007; Martínez-Peralta y Mandujano, 2009). Por ejemplo, Robbins (2003) reporta que durante los últimos años se han colectado ilegalmente más de 60 especies de cactáceas mexicanas, principalmente especies de los géneros Astrophytum Lemaire, Ariocarpus Scheidweiler y Turbinicarpus (Backeb.) Buxb. & Backeb.
Los factores mencionados son presiones negativas para las especies de cactáceas que conllevan la reducción de las poblaciones. Se sabe que bajas densidades poblacionales pueden llevar al fracaso en la reproducción sexual en sus poblaciones (Schemske et al., 1994; Suzán et al., 1994; 1998; Leirana y Parra, 1999; Martínez-Peralta y Mandujano, 2011).
A pesar de que se han realizado esfuerzos para la protección de algunas especies de cactáceas, muchas de ellas continúan siendo altamente vulnerables debido a que los planes de conservación y manejo no cuentan con información base sobre la biología de sus poblaciones y su viabilidad reproductiva (Esparza-Olguín et al., 2002; Mandujano et al., 2007). Debido a esta situación, es necesario obtener información sobre diversos aspectos ecológicos de las especies amenazadas a fin de contar con un panorama general de su estado actual de conservación (Palmer, 1987; Schemske et al., 1994). Algunos de estos aspectos ecológicos incluyen el patrón de distribución espacial, la estructura de tamaños de la población y la reproducción de los individuos. Por un lado, el patrón de distribución espacial refiere la relación que existe entre la especie y su ambiente (Krebs, 1985). La estructura de tamaños dentro de las poblaciones proporciona información sobre su composición en términos del sexo, la edad o tamaño y la reproducción de los individuos. Además, permite inferir si el reclutamiento en condiciones naturales ha ocurrido recientemente y sobre atributos de su historia de vida (Stearns, 1992; Caswell, 2001; Mandujano et al., 2001; 2007).
Los atributos de historias de vida son los caracteres relacionados con el crecimiento, la supervivencia y la reproducción, entre los que se encuentran las tasas de crecimiento corporal, el tamaño o la edad en el que se alcanza la madurez sexual y el número y tamaño de los vástagos producidos. El principio de asignación de los recursos sustenta que los organismos disponen de una cantidad finita de recursos que deben asignar entre los procesos fundamentales de crecer, sobrevivir y reproducirse (esfuerzo reproductivo), de tal manera que el número de descendencia viable que puedan aportar a las siguientes generaciones sea el mayor posible (Cody, 1966; Stearns, 1992). Por ello, el análisis de la reproducción contribuye a conocer los factores que podrían afectar el reclutamiento de los individuos (Álvarez et al., 2004). Godínez-Álvarez et al. (2003) reportan que existe información ecológica para menos de 30 especies de cactáceas, lo cual contrasta con las aproximadamente 2000 especies descritas en la familia. En este sentido, dada la carencia de estudios sobre ecología de poblaciones de este grupo, el presente trabajo pretende contribuir al conocimiento de las especies del género Astrophytum. El objetivo es evaluar la estructura poblacional, el patrón de distribución espacial y la reproducción de Astrophytum ornatum (DC.) Britton & Rose en una localidad en Cadereyta, Querétaro, México, como una medida del estado de conservación de la especie. Esta población es considerada como la más abundante en las pocas localidades que aún se conservan de la especie (Sánchez, 2006; Sánchez-Martínez et al., 2006).
Materiales y métodos
Área de estudio
La investigación se llevó a cabo en la localidad de Agua Salada en el municipio de Cadereyta de Montes (20.691°N, 99.819°W), ubicado en la parte central del estado de Querétaro, México. Presenta una vegetación de tipo matorral xerófilo, clasificado como matorral submontano (SPP, 1986; Zamudio et al., 1992), donde las especies identificadas más conspicuas fueron Agave difformis A. Berger, Bursera schlechtendalii Engl., Cylindropuntia imbricata (Haw.) DC., Ephedra compacta Rose, Jatropha dioica Sessé ex Cerv., Karwinskia mollis Schlecht., Mimosa aculeaticarpa Ortega, Mimosa depauperata Benth. y Prosopis laevigata (H. B. ex Willd.) Johnst. En esta zona predomina el clima de tipo semiseco templado con lluvias en verano (BS1kw) de la clasificación de Köppen modificada por García (1988), con temperatura mínima promedio de 12 ºC y una máxima promedio de 19.4 ºC, la temperatura media anual es de 16 ºC. La precipitación mínima promedio es de 309.3 mm y la máxima de 789.5 mm, la media anual es de 480 mm (SPP, 1986).
Especie estudiada
Astrophytum ornatum es una cactácea endémica de México, comúnmente conocida con los nombres de piojosa, biznaga algodoncillo o liendrilla. Desde el punto de vista de su comercialización es importante, principalmente por su atractivo ornamental, basado en las variaciones de los tricomas del tallo, la forma espiralada o recta de éste y su flor grande de color amarillo que aparece con precocidad (i.e., la reproducción sexual se inicia en plantas pequeñas (Arias, 1989)). El tallo es al principio esférico y después columnar. La epidermis es de color verde claro, oscuro o grisáceo con estigmas más o menos abundantes. Presenta 8 costillas rectas o espiraladas (Gómez-Sánchez, 2001). La flor es diurna, solitaria y nace en la zona apical de la planta. El fruto es globoso, semiseco, con dehiscencia apical. La semilla es navicular, de 2.5 a 3.1 mm de largo y de 1.5 a 1.7 mm de espesor (Arias, 1989). Presenta dos periodos de floración, de marzo a junio (primavera) y de agosto a septiembre (verano), y dos periodos de fructificación, de julio a septiembre y de enero a febrero. Es una especie con síndrome de polinización entomófila con vectores comúnmente del orden Hymenoptera (Arias, 1989; Zamudio et al., 1992). Se desarrolla en los matorrales desértico micrófilo y rosetófilo, presentándose también en el matorral submontano, crasicaule y en ciertas formaciones del tipo del bosque tropical caducifolio o matorral espinoso. Presenta afinidad por los lugares con humedad dentro de los cañones fluviales. Es una especie calcícola con apego por las calizas y lutitas de edades geológicas cretácicas. Se concentra en laderas de pendientes medias a pronunciadas, con mayor abundancia en la parte inferior; los suelos son someros (10 a 70 cm de profundidad) y pedregosos, de tipo Regosol calcárico, Litosol y Feozem (Arias, 1989; Ortega, 2004).
Respecto a la distribución geográfica de Astrophytum, se considera como un género endémico de México (Bravo-Hollis y Sánchez-Mejorada, 1991; Bravo-Hollis y Scheinvar, 1995; Anderson, 2001). Anderson (2001) reconoce conservadoramente cuatro especies, Astrophytum asterias (Zucc.) Lem., A. capricorne (A. Dietr.) Britton & Rose, A. myriostigma Lem. y A. ornatum, y Digitostigma caputmedusae Velazco-Macias & Nevaréz ha sido incluida dentro del género. Todas las especies de Astrophytum tienen actualmente el estatus de amenazadas y A. asterias en peligro (SEMARNAT, 2010). En particular, Astrophytum ornatum ha sido con-siderada históricamente como una especie queretano-hidalguense (Sánchez, 2006). Arias (1989) reconoce a A. ornatum como una especie endémica de México, cuya distribución se extiende en dirección Sureste-Noroeste, casi paralela a la Sierra Madre Oriental, ocupando los valles y barrancas de Hidalgo, Querétaro y Guanajuato (20º30'10" y 21º18'40"N y entre 98º21'10" y 100º30'10"W). En los últimos 10 años se ha reportado la presencia de A. ornatum más hacia el Norte, en las cuencas del río Santa María, abarcando las porciones más septentrionales de Guanajuato y los territorios inmediatos de la parte media-sur del estado de San Luis Potosí (Bárcenas, 1999). Biogeográficamente es importante por ser la especie de distribución más meridional del género Astrophytum (Arias, 1989; Vázquez Lobo et al., 2015; Carrillo-Angeles, et al., 2016).
Censo y mapeo de los individuos
En el sitio de estudio se establecieron dos parcelas per-manentes para muestrear a la población; la primera con una dimensión de 16 × 30 m ubicada a los 20º53'7.6"N y 99º41'29.7"W, la cual presentaba 20º de inclinación; la segunda de 30 × 40 m ubicada a los 20º52'50.6"N y 99º41'29.7"W con 10º de inclinación; ambas a 1856 m s.n.m. La elección de las parcelas se hizo después de recorridos intensivos en la localidad donde se reporta la ubicación de la población de A. ornatum, y a la accesibilidad al sitio; ya que la pendiente y el sustrato de roca suelta impiden el acceso a algunos de ellos. Además, en las zonas planas solamente quedan cultivos o matorrales dominados por Mimosa aculeaticarpa (uña de gato) y no se observó a ningún individuo de A. ornatum. La población de A. ornatum se considera remanente, debido a que la distribución de la especie ha quedado relegada a zonas inaccesibles, que no pueden transformarse para aprovechamientos agrícolas y cuya pendiente dificulta el paso del ganado caprino (Sánchez, 2006; Sánchez-Martínez et al., 2006).
En cada parcela se realizó el censo de los individuos por dos años consecutivos (2008 y 2009). Cada individuo fue etiquetado con alambre y chaquiras de vidrio para indicar una serie numérica y a cada uno de éstos se le midió, con ayuda de vernier y flexómetro, el diámetro en la parte más ancha de la planta y la altura en centímetros. Con estos datos se estimó el tamaño medido como el volumen del cactus utilizando la fórmula:
Donde V es el volumen (cm3), r es el radio (diámetro/2 (cm)) y h la altura (cm). Posteriormente, la población se dividió en una categoría de semillas y siete categorías de volumen para determinar la estructura de tamaños de la población (Caswell, 2001; Zepeda-Martínez et al., 2013), y se compararon mediante una prueba de chi-cuadrada para evaluar si había diferencias en el número de individuos por categoría en los años de estudio. Las semillas de más de un año de edad tienen la capacidad de germinar hasta en 70%, por lo que se considera que tienen el potencial de formar un banco de semillas (Zepeda-Martínez et al., 2013).
Patrón de distribución espacial
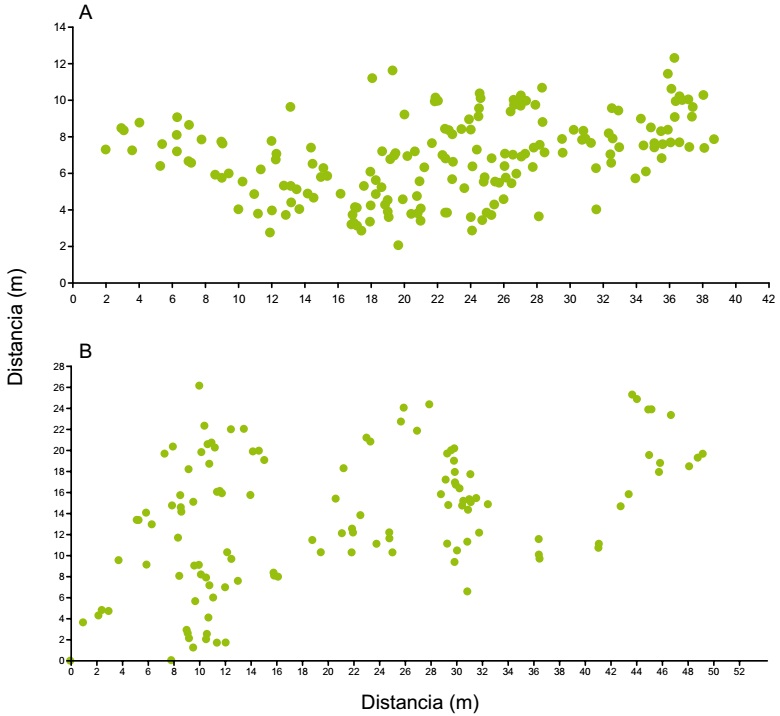
Con el fin de determinar qué patrón de distribución presenta A. ornatum, los individuos que se encontraban en las parcelas de estudio se localizaron en un mapa usando un sistema de coordenadas polares. Para ello, en ambas parcelas se tomó un punto de referencia y se midió el ángulo y la distancia, por medio de una brújula y cinta métrica, hacia el individuo más cercano y después de ese individuo al siguiente y así sucesivamente para posteriormente transformar esos datos en coordenadas x, y (Cuadro 1).
Cuadro 1 Fórmulas usadas para transformar sistema de coordenadas polares (ángulos y distancias entre individuos) en coordenadas x, y.
| Coordenadas | X | y |
|---|---|---|
| Planta 1 (valores arbitrarios para partir de este punto) | xi = 0 | y1 =0 |
| Planta 2 | x2= x1 + Sen(ángulo × distancia) | y2=y1+ Cos(ángulo × distancia) |
| Plata n | xn+1= xn+ Sen(ángulo × distancia) | yn+1= yn+ Cos(ángulo × distancia) |
La prueba de Hopkins se usó para analizar el patrón de distribución espacial de los individuos de la población muestreada (Krebs, 1985). Este análisis trabaja con el es-tadístico h, que se calcula como:
Donde Xi es la distancia de un punto aleatorio i al individuo más cercano y ri es la distancia de un organismo aleatorio i al vecino más cercano. El estadístico h se distribuye como una F con 2n grados de libertad en el numerador y lo mismo en el denominador, donde n para los grados de libertad es el número de puntos muestreados al azar. Si los organismos se encuentran agregados, la distancia de un punto al azar al organismo más cercano será mayor que la distancia entre individuos cercanos. Lo contrario ocurre si el patrón espacial es uniforme. Así, la prueba de F de dos colas, en la cual cuando h es muy pequeña, indica con certeza estadística que el patrón espacial es uniforme. El índice del patrón de distribución (IH) con un intervalo de variación de 0 a 1 puede ser estimado por:
Este índice se aproxima a la unidad conforme se incrementa la agregación y a cero cuando la uniformidad es máxima. Bajo la hipótesis nula de aleatoriedad, el índice del patrón de distribución es de 0.5 (Krebs, 1985).
Fecundidad
La fecundidad de los individuos agrupados en cada categoría de tamaño se calculó a partir de una colecta de un total de 55 frutos de plantas diferentes. Se obtuvieron de 6 a 9 frutos completos para estimar el número de semillas en cada una de las diferentes categorías de tamaño. Colectamos un fruto por individuo en 29 plantas (36 frutos dentro de las parcelas). Para aumentar la muestra se colectaron 19 frutos completos, en 15 plantas localizadas fuera de las parcelas de estudio. A las plantas fuera de las parcelas se les midió el tamaño para asignar la producción de semillas al tamaño correspondiente. La muestra de frutos se colectó en julio de 2008 y los censos del número de estructuras reproductivas (botones, flores, frutos) por planta reproductiva se hicieron durante las temporadas reproductivas (28 de febrero y 16 de julio de 2008). En cada planta dentro de las parcelas se contabilizó el número de frutos totales; el número de frutos por planta reproductiva fue multiplicado por el número promedio de semillas que contenían. El total de semillas por planta de cada clase de tamaño fue sumado para todas las plantas y dividido entre el número de plantas analizadas para la clase de tamaño. En esta forma, la medida de esfuerzo reproductivo incluye la producción de las estructuras reproductivas y el número de semillas promedio por individuo, que pertenece a cada clase de tamaño. La cantidad de frutos y semillas que produjeron los individuos reproductivos fue comparada mediante análisis lineales generalizados (GLIM) del tipo Poisson, con ajuste de sobredispersión, con el programa JMP versión 10 (SAS Institute, 2007).
Mortalidad
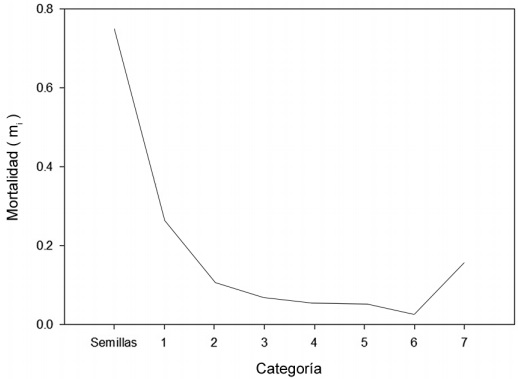
La probabilidad de muerte se estimó a partir de los censos poblacionales realizados en 2008 y 2009. La probabilidad de mortalidad en cada categoría de tamaño del ciclo de vida se calculó de la siguiente manera: mi= 1-px donde mi es la probabilidad de mortalidad en la clase i y px es la suma de probabilidades de permanencia, retrogresión y crecimiento en cada categoría (Caswell, 2001). En el caso de las semillas, la probabilidad de mortalidad de las mismas se calculó con el promedio de la proporción de semillas que en diversos ensayos de germinación germinan y el reclutamiento observado en campo (Zepeda-Martínez et al., 2013).
Resultados
Patrón de distribución espacial
La densidad poblacional promedio de A. ornatum en la población estudiada fue de 0.22 ind/m2, es decir de un individuo por cada 4.5 m2 (Cuadro 2), con un total de 373 Individuos. El índice de agregación obtenido para toda la población a partir de la prueba de Hopkins fue I H=0.88 (n=40, h=7.46, F 0.01=0.588 y F 0.975=1.7 con 60 grados de libertad (g.l.) en el numerador y 120 g.l. en el denominador), por lo que se rechaza la hipótesis nula de aleatoriedad, determinando que A. ornatum presenta un patrón de distribución de tipo agregado (Figs. 1A y B).
Cuadro 2: Densidad poblacional de Astrophytum ornatum (DC.) Britton & Rose en las dos parcelas de estudio localizadas en Cadereyta, Querétaro, México.
| Área m 2 | No. De individuos | Densidad ind/m 2 | |
|---|---|---|---|
| Parcela 1 | 480 | 240 | 0.5 |
| Parcela 2 | 1200 | 133 | 0.11 |
| Total | 1680 | 373 | 0.22 |
Estructura poblacional y reproducción
El ciclo de vida comprende una categoría del estado de semillas y siete categorías de tamaño (volumen en cm3, Fig. 2). El análisis de la distribución de frecuencias, sin considerar a las semillas, muestra que las categorías uno, tres y cuatro presentan un mayor porcentaje de individuos con respecto al resto. En el año 2008, 27.69% de los individuos se encontraba en la categoría tres (30.001-600 cm3), 19.62% en la categoría cuatro (600.001-2500 cm3) y 12.37% en la uno (no reproductivos, de 0.006-7cm3). Esta estructura no difirió significativamente en el año 2009 (χ2=12.36, g.l.=6, P=0.0542; Fig. 2).
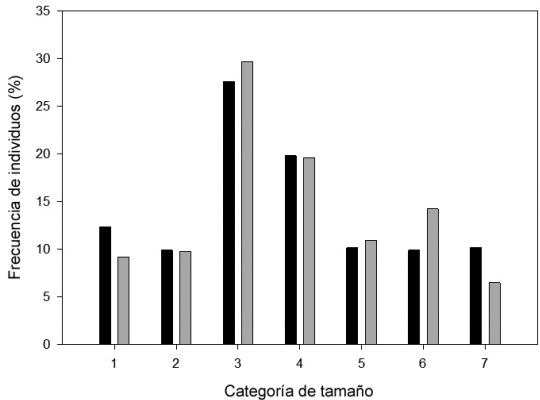
Figura 2 Estructuras poblacionales de Astrophytum ornatum (DC.) Britton & Rose observadas en una población localizada en Cadereyta, Querétaro, México. Barras negras corresponden al censo en el año 2008 (N=373 individuos) y las grises al de 2009 (N=337). Los intervalos de clase para las categorías de tamaño: 1=0.006-7 cm3, 2≥7≤30 cm3, 3≥30≤600 cm3, 4≥600≥2500 cm3, 5≥2500≤4100 cm3, 6≥4100≤9200 cm3, 7≥9200 cm3.
Solamente 34% del total de individuos son re-productivos. El promedio de flores por planta en la población (N=328 individuos en etapas reproductivas) es bajo (0.82 ± 1.6, desviación estándar (d.e.) flores/ind) con un rango amplio a nivel individual de 0 hasta 11 flores/ind. El número de frutos que en promedio produce un individuo reproductivo también es reducido (0.25 ± 1.02 d.e.) con un rango amplio de 0 a 10 frutos/ind. De acuerdo con el número de flores y frutos observados, el asentamiento de frutos (proporción de flores que forman frutos) fue de 31% en 2008 y de 2.5% en la temporada reproductiva de 2009. El número de semillas por fruto es de 120 ± 49 d.e. y hay una amplia variación en el número de semillas que producen las plantas en las diferentes categorías de tamaño (Fig. 3, Cuadro 3). El esfuerzo reproductivo es mayor en cuanto al número de estructuras reproductivas (botones, flores y frutos) en las últimas tres categorías del ciclo de vida (Fig. 3). La producción de frutos difiere entre las categorías de tamaño (Cuadro 3, χ2=12.85, g.l.=5, P=0.024; n=84 plantas reproductivas), donde los individuos de las categorías 6 y 7 son los que producen el mayor número de frutos (contraste de clases 2 a la 5 contra la 6 y 7, χ2=4.92, g.l.=1, P=0.026). La producción de semillas también difiere entre las categorías de tamaño (Cuadro 3, χ2=14.15, g.l.=5, P=0.014; n=84 plantas reproductivas), en este caso, las categorías dos y tres presentan un mayor número promedio de semillas por fruto (Fig. 3, Cuadro 3, χ2=10.69, g.l.=1, P=0.0017).

Figura 3 Distribución del esfuerzo reproductivo en términos del número de botones florales, flores y frutos, y promedio de semillas (± error estándar) por fruto de los individuos de Astrophytum ornatum (DC.) Britton & Rose; en cada una de las categorías de tamaño de las plantas (1=0.006-7 cm3, 2≥7≤30 cm3, 3≥30≥600 cm3, 4≥600≤2500 cm3, 5≥2500≤4100 cm3, 6≥4100≤9200 cm3, 7≥9200 cm3, volumen). Los datos de fecundidad corresponden a 2008.
Cuadro 3 Número de frutos y semillas per cápita (promedio ± desviación estándar) en los individuos reproductivos de Astrophytum ornatum (DC.) Britton & Rose de diferentes categorías de tamaño (1=0.006-7 cm3, 2≥7≤30 cm3, 3≥30≤600 cm3, 4≥600≤2500 cm3, 5≥2500≤4100 cm3, 6≥4100≤9200 cm3, 7≥9200 cm3). Los individuos de la primera categoría no son reproductivos. Datos obtenidos en dos parcelas de estudio localizadas en Cadereyta, Querétaro, México, durante 2008 (N=número de individuos).
| Categoría de tamaño | N | Número de frutos | Número de semillas |
|---|---|---|---|
| 1 | 0 | ||
| 2 | 2 | 2.5 (0.70) | 188 (43.8) |
| 3 | 5 | 2.2 (1.3) | 276.5 (271.4) |
| 4 | 14 | 2.1 (1.3) | 50.9 (86.5) |
| 5 | 19 | 2.7 (1.9) | 36.6 (67.1) |
| 6 | 22 | 2.9 (2.3) | 31.25 (87.5) |
| 7 | 22 | 4.1 (3.5) | 71.1 (145.2) |
En el censo correspondiente al año 2009 no se encontró reclutamiento de plántulas y se observó mortalidad en todas las categorías de tamaño. Las primeras categorías presentan una mayor probabilidad de mortalidad, la cual va disminuyendo en aquellas con individuos de mayor tamaño. La categoría de semillas presentó el valor más alto (0.72), seguido de la categoría uno (0.26) y las categorías siete (0.13) y dos (0.11) (Fig. 4).
Discusión
El tamaño y la dinámica de las poblaciones de A. ornatum no son conocidos en conjunto. Arias (1989) estimaba que aún existen varias poblaciones estables, aunque se aprecia un deterioro general de las mismas. No obstante, varias poblaciones se han perdido (Sánchez, 2006; Sánchez-Martínez et al., 2006). Según lo observado por Ortega (2004) en individuos de A. ornatum en la zona de la Presa de Zimapán, Hidalgo, México, se producen 17 frutos viables por año en esa población. Éstos a su vez producen en promedio 42 (± 24.2 d.e.) semillas por fruto, mientras que el porcentaje de germinación reportado es de 56%. En el caso de la población estudiada en Cadereyta, encontramos tres veces más frutos producidos y semillas que lo reportado para la especie en otras poblaciones (Arias, 1989). Sin embargo, el asentamiento (fruit set, proporción de flo-res que pasan a formar fruto) de frutos es de los más bajos que se han registrado si se le compara con lo reportado para otras especies de cactáceas, porque el promedio estimado es de 75% (Mandujano et al., 2010). Aunado a ello, como las semillas son la única forma de dispersión de la especie y se ha detectado granivoría por parte de hormigas del género Iridomyrmex que se alimentan de éstas, principalmente de su funículo después de que ha ocurrido la dehiscencia (Arias, 1989), deben ser pocas las semillas que pueden llegar a establecerse.
El establecimiento de las semillas en A. ornatum sigue la relación de nodriza protegido, es decir, ocurre bajo la protección de arbustos, huecos en las rocas o bajo plantas adultas a las que se agregan, en donde se reduce la temperatura y la radiación (Arias, 1989; Ortega, 2004; Zepeda-Martínez et al., 2013). El patrón de distribución espacial agregado es común en especies que presentan asociación nodriza protegido, porque las zonas favorables para el establecimiento se reducen a las que aportan la protección contra los factores abióticos y bióticos (Jordan y Nobel, 1982; Godínez-Álvarez et al., 2003; Mandujano et al., 2007).
Arias (1989) confirma que el desarrollo inicial de los individuos de la especie es lento y comprueba que en la mayoría de las poblaciones estudiadas existen reemplazos en la población. Además, menciona que, durante sus recorridos en campo, en 16 de las 19 localidades valoradas, se presentan plántulas e individuos inmaduros. Ortega (2004) menciona que A. ornatum mantiene su crecimiento en forma globosa hasta los 15 a 25 cm y posteriormente se desarrolla como columnar. Diez localidades estudiadas durante el año 2005 por el personal del Jardín Botánico Regional de Cadereyta, en el centro de Querétaro, arrojaron tamaños poblacionales pequeños (menos de 50 individuos por localidad), pero en todas se encontraron plántulas (Sánchez, 2006; Sánchez-Martínez et al., 2006), por lo que la población que estudiamos es actualmente la más grande que queda de la especie. García-Manjares et al. (2008) indican, en un estudio realizado en la Barranca de Metztitlán, que las alturas de los individuos oscilan entre 1 y 119 cm, lo que implica reclutamiento reciente. El intervalo de altura con mayor frecuencia difiere entre las poblaciones de A. ornatum, en una fue de 50 a 60 cm y en la otra de 30 a 40 cm. En el caso de la población de Cadereyta, encontramos que la categoría de los más pequeños ocupa el tercer lugar en frecuencia de individuos, donde la altura registrada de A. ornatum en la clase 1 de volumen fue de 0.2 a 2 cm y se registraron casi 40 plantas, por lo que la estructura poblacional sugiere que sí ha habido reclutamiento. Sin embargo, en los dos años de este estudio no se registraron plántulas recién establecidas en la población de A. ornatum; lo que sugiere que es necesaria una observación estrecha de los procesos de reclutamiento, porque las plantas pueden mantenerse en el mismo estado de desarrollo por varios años, en este caso, como juvenil o plántula (Caswell, 2001).
La estructura de tamaños observada en una población ayuda a comprender los procesos históricos y probablemente actuales que determinan la dinámica poblacional de una especie. Así, la forma de estructura poblacional refleja el reclutamiento de varios años atrás, aunque no podemos datar cuándo sucedió. La variedad de estructuras poblacionales que se pueden encontrar en la naturaleza es resultado de diferencias entre la producción de semillas, la tasa de germinación, la formación de bancos de semillas, el crecimiento anual dado por abundancia o escasez de recursos, el crecimiento clonal, la supervivencia y la fecundidad de los individuos, dependiendo así de la interacción de todos estos procesos (Mandujano et al., 2001; Martínez-Ávalos, 2007).
En el caso de la población estudiada de A. ornatum, la alta proporción de individuos en las tres primeras categorías de tamaño sugiere que ha habido reclutamiento reciente. La forma de la curva de la estructura de tamaños de A. ornatum sugiere este patrón, como se ha reportado para otras especies perennes, ya que, por el contrario, la ausencia de ciertas tallas o la presencia de valles en las gráficas de estructura poblacional indican el reclutamiento escaso o en pulsos temporales (Mandujano et al., 2001; Godínez-Álvarez et al., 2003). La evidencia de reclutamiento concuerda con lo reportado para una población de la misma especie en Zimapán (Ortega, 2004) y otra en Metztitlán con tamaños poblacionales grandes (García-Manjares et al., 2008), ambas en el estado de Hidalgo. Sin embargo, Arias (1989) reporta en dos de tres poblaciones de A. ornatum estudiadas en la zona árida queretano-hidalguense una relación inversa a la mencionada anteriormente, lo cual sugiere que las poblaciones pequeñas no presentan reclutamiento.
La época reproductiva de A. ornatum es larga, extendiéndose de noviembre a mayo. En este estudio se observó que solo 34% de los individuos de la población son reproductivos y que se presentan una mayor cantidad de estructuras reproductivas (botones florales, flores y frutos) en las últimas categorías de la estructura de tamaños, pero, en promedio, un mayor número de semillas por fruto en las primeras categorías reproductivas (dos y tres). Esto refleja que la historia de vida de la especie en la localidad de estudio tiene una asignación de recursos diferencial en las distintas categorías del ciclo de vida. La historia de vida de un organismo puede definirse como la historia evolutiva de las características que componen su ciclo de vida, las cuales sintetizan los procesos de reproducción, crecimiento y supervivencia, y determinan su adecuación en el ambiente que habita (Cody, 1966; Stearns, 1992). En el caso de la especie de estudio y bajo el contexto descrito anteriormente, si los individuos de mayor tamaño asignan más recursos a la producción de estructuras reproductivas entonces, el tamaño de los frutos disminuirá, mientras que en las categorías en las que en promedio se producen menor número de estructuras reproductivas, la asignación de recursos estará enfocada a producir frutos de mayor tamaño y por lo tanto una mayor producción de semillas.
Los factores que condicionan el desarrollo natural de A. ornatum son principalmente de origen humano. El género entero sigue siendo predilecto de los coleccionistas. Particularmente esta especie atrae mucho interés debido, entre otros factores, a los nuevos hallazgos de poblaciones realizados durante el 2004 en la frontera guanajuatense-potosina que han permitido localizar formas intermedias entre A. ornatum y A. myriostigma. Las variaciones híbridas (consideradas variedades por los coleccionistas y en algunos tratados como especies) de Astrophytum ornatum (conocidas como A. glabrescens, A. niveum, A. mirbelii y A. virens) presentan una alta demanda en el mercado (Sánchez, 2006). El saqueo de plantas, plántulas y semillas podría estar activo en estas poblaciones por lo que es necesario un monitoreo permanente (Sánchez, 2006). Como ya se hizo evidente, la producción de frutos y semillas es muy baja y el reclutamiento no ocurre todos los años.
Los peligros originados localmente están también presentes y de ellos, el pastoreo caprino es frecuente, aunque no existen estudios que correlacionen la perturbación de este tipo de ganado con los disturbios de consecuencias para las poblaciones, en todo su ámbito de distribución (Arias, 1989). La carga animal aún en algunos sitios ya declarados como reservas ecológicas (Reservas de la Biosfera de Metztitlán y Sierra Gorda) es alta (Sánchez, 2006). Adicionalmente, existe desde varios años atrás un problema con los burros ferales que también destruyen las biznagas (SEMARNAP, 1999).
De acuerdo con la evaluación de la condición local de riesgo, A. ornatum es una especie relacionada con la Zona Árida Querétaro-Hidalguense que presenta especificidad de hábitat y se distribuye preferentemente en las cañadas de los ríos locales, y se asocia con el matorral micrófilo, rosetófilo y submontano de la región. Dada la extensión de los cauces de los ríos es posible que la especie encuentre sitios de resguardo para su establecimiento, pero algunos de ellos ya se encuentran muy deteriorados o amenazados por ser zonas de extracción del mármol y otros materiales (Sánchez-Martínez et al., 2006). Las poblaciones más numerosas se han registrado en sitios con mayor pendiente, como es el caso en este estudio. En poblaciones ubicadas en sitios de menor pendiente es notable el grado de deterioro que presentan ya que son dañadas por el sobrepastoreo, por la extracción de leña y de plantas de A. ornatum, y por el ganado equino (burros) (Sánchez-Martínez et al., 2006). Cabe señalar que en los dos años de estudio no se registró saqueo de plantas en los lugares censados, dada la dificultad para acceder al sitio y la cercanía a ejidos. En síntesis, es primordial que se preserve el hábitat relicto que mantiene la población de A. ornatum, dado que la información generada en esta investigación y en un estudio demográfico en el sitio indican que es una población frágil, que presenta individuos reproductivos, la producción de semillas viables, pero su dinámica poblacional sugiere que tiende a decrecer, dado que en los años de estudio no se observó reclutamiento y se encontró baja precipitación (Zepeda-Martínez et al., 2013). Se sabe que hay limitaciones de su muestreo y en especies perennes es deseable obtener información por periodos amplios de tiempo para incrementar la certidumbre de los resultados. No obstante, esta población es la más grande que se mantiene en la región, por lo que deben concretarse los esfuerzos para su preservación.

