










Services on Demand
Journal
Article
 text in
text in  English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista mexicana de fitopatología
On-line version ISSN 2007-8080Print version ISSN 0185-3309
Rev. mex. fitopatol vol.41 n.3 Texcoco Sep. 2023 Epub Oct 13, 2023
https://doi.org/10.18781/r.mex.fit.2210-2
Artículos científicos
Identificación de especies y razas fisiológicas de Xanthomonas aisladas de jitomate (Solanum lycopersicum) y chile (Capsicum annuum) en Sinaloa, México
1 Centro de Investigación en Alimentación y Desarrollo, Coordinación Regional Culiacán, Carretera a ElDorado km 5.5, Culiacán, Sinaloa, CP 80110, México.
La mancha bacteriana del jitomate y chile, causada por cuatro especies de Xanthomonas y diversas razas, es una de las enfermedades de mayor impacto en la horticultura a nivel mundial. El objetivo de este estudio fue identificar a las especies y razas fisiológicas de Xanthomonas spp. presentes en los cultivos de jitomate (Solanum lycopersicum) y chile (Capsicum annuum) en Sinaloa, México. Para ello, se recolectaron muestras con síntomas de mancha bacteriana en campos comerciales tanto de jitomate como de chile, distribuidos en diferentes municipios de Sinaloa. Noventa y tres bacterias se aislaron en los medios agar nutritivo (AN) y agar levadura-dextrosa-carbonato de calcio (YDC). Un total de 47 aislados bacterianos se identificaron como Xanthomonas mediante la combinación de pruebas morfológicas, patogénicas, bioquímicas, fisiológicas y moleculares. Además, para la caracterización de razas fisiológicas de cada una de las cepas, se utilizaron cuatro líneas diferenciales de jitomate y seis de chile. El análisis realizado mediante iniciadores específicos para su diagnóstico molecular confirmó que, el 83% correspondió a X. euvesicatoria, donde se detectaron las razas P0, P3, P6, y P10; el 10.6% a X. perforans con las razas fisiológicas T1, T2, T3 y T5, y el 6.4% a X. vesicatoria, detectándose las razas T1, T3 y T5. Entretanto, con las líneas diferenciales de jitomate se detectaron a las razas T1, T2, T3 y T5; mientras que con las líneas diferenciales de chile se detectaron las razas P0, P3, P6, P8 y P10. Esta información actualiza los datos previos sobre la distribución de razas de Xanthomonas que infectan al chile en Sinaloa, debido a que se reporta por primera vez a las razas P6 y P10.
Palabras clave: mancha bacteriana; líneas diferenciales; reacción de hipersensibilidad
Bacterial spot of tomato and chili pepper, caused by four species of Xanthomonas and various races, is one of the diseases with the greatest impact on horticulture worldwide. The aim of this study was to identify the species and physiological races of Xanthomonas present in tomato (Solanum lycopersicum) and chili pepper (Capsicum annuum) crops in Sinaloa, Mexico. For this purpose, samples with typical symptoms of bacterial spot were collected in commercial fields of the different municipalities in the state of Sinaloa. Ninety-three bacteria were isolated on semi-selective medium as nutrient agar and yeast extract-dextrose. A total of 47 bacteria were identified as Xanthomonas by a combination of morphological, pathogenic, biochemical, physiological, and molecular tests. In addition, to characterize the morphological race of each strain, differential tomato (four) and chili pepper (six) lines were used. Molecular diagnostic using specific primers indicated that 83% corresponded to X. euvesicatoria, 10.6% to X. perforans, and 6.4% to X. vesicatoria. At the physiological race level, races T1, T2, T3, and T5 were detected in tomato; while the Xanthomonas races detected in chili pepper were P0, P3, P6, P8 and P10. This information updates previous data on the distribution of races of Xanthomonas that infect chili peppers in Sinaloa, since races P6 and P10 are reported for the first time.
Keywords: Bacterial spot; differential lines; hypersensitivity reaction
La mancha bacteriana en jitomate (S. lycopersicum) y chile (C. annuum) causada por Xanthomonas spp. es una de las enfermedades más importantes y destructivas en estos cultivos a nivel mundial (Potnis et al., 2015; Sharma y Bhattarai, 2019; Bogatzevska et al., 2021; Rotondo et al., 2022). Aunque esta enfermedad ocurre en todas las regiones productoras de jitomate y chile, es más severa en regiones subtropicales y tropicales donde el patógeno es favorecido por la alta temperatura y lluvias constantes (Koike et al., 2007).
Los síntomas foliares iniciales causados por Xanthomonas spp. consisten en manchas irregulares acuosas que posteriormente se tornan de color marrón oscuro a negro y por lo general miden <5 mm de diámetro. A medida que avanza la enfermedad, las manchas pueden coalecer y dar lugar a áreas necróticas grandes en las hojas. En infecciones avanzadas y severas, las plantas pueden deshojarse, especialmente en la parte inferior. Las infecciones en la fruta se observan de manera irregular como costras ásperas, levantadas, de color marrón, que miden de 2-5 mm de diámetro. Las manchas en los frutos se agrupan cerca del extremo del pedúnculo (Koike et al., 2007).
A nivel mundial existen cuatro especies de Xanthomonas (X. euvesicatoria, X. vesicatoria, X. perforans y X. gardneri) asociadas comúnmente a la enfermedad conocida como mancha bacteriana en jitomate y chile (Jones et al., 2004). Actualmente, se reportan cinco razas de Xanthomonas spp. para jitomate: raza T1 reportada en X. euvesicatoria, raza T2 en X. vesicatoria y X. gardneri, razas T3 y T4 reportadas en X. perforans; así como, la raza T5, la cual se reportó como una recombinación entre la raza T3 y T4 de X. perforans (Adhikari et al., 2020; Jibrin et al., 2022). Mientras que, en el caso de chile, se han reportado 11 razas (P0, P1, P2, P3, P4, P5, P6, P7, P8, P9 y P10). La determinación de las razas en las cepas de Xanthomonas spp. se basa en la reacción de hipersensibilidad en respuesta a las proteínas efectoras mediadas a través del sistema de secreción tipo III en las células del hospedante y con reconocimiento por proteínas de resistencia específicas en plantas diferenciales de jitomate y chile (Stall et al., 2009).
En jitomate, existen estudios previos donde se determinó la diversidad de razas y/o especies de Xanthomonas en cepas colectadas en EE. UU. (Bouzar et al., 1994; Rotondo et al., 2022), India, Argentina, España, Nueva Zelanda, Francia, Hungría (Bouzar et al., 1994), Taiwán (Hartman y Yang, 1990; Bouzar et al., 1994; Lue et al., 2010), México (Bouzar et al., 1994; Bouzar et al., 1996), Islas del Caribe (O’Garro, 1998), Brasil (Bouzar et al., 1994; Quezado-Duval y Camargo, 2004), Canadá (Bouzar et al., 1994; Abbasi et al., 2015), Australia (Bouzar et al., 1994; Roach et al., 2018), Tailandia (Sitthitanasin et al., 2020), Bulgaria (Bogatzevska et al., 2021) y África (Jibrin et al., 2022).
Para el cultivo de chile se han identificado razas y/o especies de Xanthomonas con cepas provenientes de Argentina, Brasil, Hungría, Senegal, España, Tongo (Bouzar et al., 1994), EE. UU. (Bouzar et al., 1994; Kousik y Ritchie 1995; Rotondo et al., 2022), Taiwán (Hartman y Yang, 1990; Bouzar et al., 1994; Lue et al., 2010), México (Bouzar et al., 1994; 1996; Carrillo-Fasio et al., 2001), Islas del Caribe (Bouzar et al., 1994; O’Garro, 1998), Australia (Bouzar et al., 1994; Roach et al., 2018), Tailandia (Sitthitanasin et al., 2020), Bulgaria (Bogatzevska et al., 2021) y África (Jibrin et al., 2022).
En Sinaloa, principal estado productor de jitomate y chile en México, la mancha bacteriana presenta una amplia distribución y se observa continuamente en forma epidémica, ocasionando pérdidas importantes en la producción de ambos cultivos (Carrillo-Fasio et al., 2001). En este sentido, algunos estudios previos que estudiaron la diversidad de razas de Xanthomonas en estos cultivos, se basaron en cepas aisladas de campos de Sinaloa y Sonora (Bouzar et al., 1994; 1996; Carrillo-Fasio et al., 2001). No obstante, la información de la distribución y presencia de razas de Xanthomonas spp. en los cultivos de jitomate y chile debe actualizarse con base en estudios complementados con datos moleculares. Por lo que, el objetivo de este estudio fue identificar a las razas y especies de Xanthomonas que infectan a los cultivos de jitomate y chile en el estado de Sinaloa, México mediante la combinación de pruebas morfológicas, patogénicas, bioquímicas, fisiológicas y moleculares.
Materiales y métodos
Recolección de muestras. Durante octubre de 2016 a marzo de 2017, se recolectaron hojas de jitomate y chile con síntomas típicos de mancha bacteriana en 47 campos comerciales distribuidos los municipios de Ahome (8), Guasave (7), Navolato (7), Culiacán (9), Elota (5) y Escuinapa (11), en Sinaloa, México. Los cultivos se encontraban en diversas etapas de desarrollo; desde la etapa inicial hasta la etapa de producción.
Aislamiento de bacterias. Las hojas colectadas se desinfestaron superficialmente en alcohol al 70% por 30 segundos, se cortaron en pequeños trozos a partir de los márgenes de la lesión y se trituraron en agua destilada estéril. La suspensión se sembró por estrías sobre placas Petri con medio de cultivo Agar nutritivo (AN) y medio de levadura-dextrosa-carbonato de calcio (YDC) Las placas se incubaron a 28 °C durante 48 h en oscuridad continua, y posteriormente se seleccionaron y purificaron las colonias redondas, convexas, amarillas y de consistencia mucosa. Los cultivos puros se conservaron en tubos con agua a 4 °C para su almacenamiento a corto plazo y en glicerina al 20% a temperatura de -20 °C para su almacenamiento a largo plazo (Schaad et al., 2001).
Reacción de hipersensibilidad. Con el fin de corroborar que las cepas aisladas fueran patógenas, se realizó la prueba de hipersensibilidad, utilizando plantas de tabaco (Nicotiana tabacum) de dos meses de edad. Se preparó una solución bacteriana con una concentración de 3 ´ 108 UFC mL-1 según la escala de McFarland´s (Kyraly et al., 1974) y con una jeringa hipodérmica se infiltró la solución en el envés de la hoja, identificando cada área infiltrada con el dato o registro de la cepa. Las plantas se colocaron en condiciones de invernadero y se monitorearon durante las primeras 48 h (Király et al., 1974). Aquellas cepas que indujeron una reacción de hipersensibilidad positiva fueron las que se seleccionaron para continuar con su estudio.
Pruebas bioquímicas y fisiológicas. De un total de 93 cepas aisladas, solo 47 (39 de jitomate y 8 de chile) presentaron las características fenotípicas reportadas para Xanthomonas spp., y con estas se realizaron las pruebas bioquímicas y fisiológicas (tinción de Gram, metabolismo oxidativo/fermentativo de los carbohidratos, oxidasa y almidón) descritas por Schaad et al. (2001). Todas las pruebas se realizaron en tres ocasiones.
Extracción de ADN y amplificación por PCR. La lisis celular de la bacteria se realizó por medio de choque térmico a partir de colonias jóvenes cultivadas por dos días en medio YDC. Para ello, se colocó una colonia pura en un tubo de microcentrífuga de 1.5 mL, conteniendo 100 µL de agua inyectable y se homogenizó la solución. Posteriormente, se colocó durante 15 min a 95 °C en un termoblock (Labnet) y después se transfirió inmediatamente a un recipiente con hielo durante 7 min. Una vez transcurrido el tiempo, se centrifugó a 10 000 x g por 15 min, se decantó el sobrenadante y se resuspendió la pastilla en 50 µL de agua inyectable.
Cuadro 1 Iniciadores específicos para la identificación de especies de Xanthomonas en cepas obtenidas de jitomate y chile en Sinaloa, México.
| Species | Primer name | Gene sequence | Amplicon (pb) |
|---|---|---|---|
| Xanthomonas spp. | RST 65 | 5’-GTCGTCGTTACGGCAAGGTGGTCG-3’ | 420 |
| RST 69 | 3’-TCGCCCAGCGTCATCAGGCCATC-5’ | ||
| X. euvesicatoria | Bs-XeF | 5′CATGAAGAACTCGGCGTATCG-3′ | 173 |
| Bs-XeR | 3′-GTCGGACATAGTGGACACATAC-5′ | ||
| X. vesicatoria | Bs-XvF | 5′-CCATGTGCCGTTGAAATACTTG-3’ | 138 |
| Bs-XvR | 3′-ACAAGAGATGTTGCTATGATTTGC-5´ | ||
| X. perforans | Bs-XpF | 5′-GTC GTG TTG ATG GAG CGT TC-3′ | 197 |
| Bs-XpR | 3′-GTGCGAGTCAATTATCAGAATGTGG-5′ | ||
| X. gardneri | Bs-XgF | 5′-TCAGTGCTTAGTTCCTCATTGTC-3’ | 154 |
| Bs-XgR | 3′-TGACCGATAAAGACTGCGAAAG-5′ |
La amplificación por PCR se llevó a cabo con cinco pares de iniciadores (Sigma) que flanquean al gen HrpB2, especificados en el Cuadro 1 (Obradovic et al., 2004), y una mezcla de reacción indicada en el Cuadro 2. Los reactivos pertenecen al kit GoTaq® PCR Core System I. Las condiciones de amplificación fueron: una desnaturalización inicial a 95 ºC por 5 min, 35 ciclos de 94 ºC por 1 min, 56 ºC por 1 min, 75 ºC por 1 min; y una extensión final a 75 ºC por 10 min. Las reacciones se llevaron a cabo en un termociclador Proflex PCR system (Applied Biosystem).
Cuadro 2 Mezcla de reacción para la PCR para la identificación de especies de Xanthomonas.
| Reactive (Initial concentration) | Final Volume (µL) | Final Concentration |
|---|---|---|
| Buffer (10X) | 2.5 | 1 X |
| MgCl2 (50mm) | 1.2 | 2.5 mM |
| Taq Pol (5 u/mL) | 0.125 | 0.25 U |
| Primer RST65 (10 mM) | 1.25 | 0.5 mM |
| Primer RST 69 (10 mM) | 1.25 | 0.5 mM |
| Primer Bs-XeF (10 mM) | 0.5 | 0.2 mM |
| Primer Bs-XeR (10 mM) | 0.5 | 0.2 mM |
| Primer Bs-XvF (10 mM) | 1.0 | 0.4 mM |
| Primer Bs-XvR (10 mM) | 1.0 | 0.4 mM |
| Primer Bs-XpF (10 mM) | 0.75 | 0.3 mM |
| Primer Bs-XpR (10 mM) | 0.75 | 0.3 mM |
| Bs-XgF (10 mM) | 0.75 | 0.3 mM |
| Bs-XgR (10 mM) | 0.75 | 0.3 mM |
| DNTP’s (10 mM) | 0.5 | 0.2 mM |
| DNA | 1 | |
| H2O cbp | Gauge to 25 |
MgCl2: Cloruro de Magnesio, Taq Pol: ADN polimerasa, dNTPs: deoxinucleótidos trifosfato, Iniciador sentido para género Xanthomonas: RST65, iniciador antisentido para género Xanthomonas. RST 69, iniciador sentido para X. euvesicatoria Bs-XeF, iniciador antisentido para X. euvesicatoria: Bs-XvR, iniciador sentido para X. vesicatoria. Bs-XeF, iniciador antisentido para X. euvesicatoria: Bs-XeR, iniciador sentido para X. perforans: Bs-XpF, iniciador antisentido para X. perforans: Bs-XpR; iniciador sentido para X. gardneri. Bs-XgF, iniciador antisentido para X. gardneri: Bs-XgR.
Los amplicones se visualizaron mediante electroforesis en gel de agarosa a 1% usando una cámara de electroforesis (BioRad)y se sometieron a 60 V durante 50 min. Posteriormente, los geles se visualizaron en un transluminador (Benchtop) con luz UV.
Razas fisiológicas de Xanthomonas en chile. Las razas fisiológicas de Xanthomonas spp. se determinaron utilizando el material de chile cv. Early California Wonder (susceptible) y un conjunto de líneas isogénicas derivadas de Early California wonder (ECW), incluyendo a ECW-10R, ECW-20R, ECW-30R, PT235047 y ECWPLUS. Las líneas ECW-10R, ECW-20R, ECW-30R contienen los genes de resistencia 10R, 20R y 30R de Capsicum pubescens, respectivamente; mientras que la línea ECWPLUS tiene los genes 10R, 20R, 30R, 50R y 60R (Cuadro 3).
Las plantas se cultivaron en invernadero bajo condiciones de humedad relativa (60-70%) y temperatura (25 °C) controladas durante 60 días. Posteriormente, se llevó a cabo la infiltración, la cual se realizó con una solución bacteriana de 3 ´ 108 UFC mL-1 usando una jeringa de 5 mL sin aguja, teniendo cuidado que la infiltración abarcara gran parte del área de la hoja. Un grupo de plantas diferenciales inoculadas con agua destilada, sirvieron como control. Las plantas se evaluaron a las 48 h y a la semana después de inoculación para determinar las reacciones de hipersensibilidad (RH) y de susceptibilidad (S). En la prueba se consideró RH cuando se observaron manchas color café oscuro. En el caso exclusivo de la línea ECWPLUS, una RH se consideró como una mancha amarillenta seca; mientras que, se consideró una reacción S cuando en la hoja se observó flacidez y acuosidad. Los resultados se cotejaron con la información del Cuadro 3 (Stall et al., 2009).
Cuadro 3 Clasificación de razas de Xanthomonas spp. en chile (Capsicum annuum) por reacciones de susceptibilidad e hipersensibilidad.
| ECW | ECW10 | ECW20 | ECW30 | CP | ECW-12356 | RACE |
|---|---|---|---|---|---|---|
| S | HR | HR | HR | HR | HR | 0 |
| S | S | HR | HR | HR | HR | 1 |
| S | HR | HR | S | S | HR | 2 |
| S | S | HR | S | HR | HR | 3 |
| S | S | S | HR | HR | HR | 4 |
| S | HR | S | S | S | HR | 5 |
| S | S | S | S | HR | R | 6 |
| S | S | HR | HR | S | HR | 7 |
| S | S | HR | S | S | HR | 8 |
| S | S | S | HR | S | HR | 9 |
| S | S | S | S | S | R | 10 |
S= Susceptible.; HR= Reacción de hipersensibilidad.
Razas fisiológicas de Xanthomonas en jitomate. Para la identificación de razas fisiológicas de las cepas de Xanthomonas spp. en jitomate, se utilizaron las líneas diferenciales Hawaii7998, Hawaii7891, PI114490 y LA716 (Cuadro 4). Utilizando la misma metodología mencionada anteriormente para chile. Las plantas se evaluaron a las 48 h y a la semana después de inoculación se registraron las reacciones de hipersensibilidad (RH) y de susceptibilidad (S). La prueba se consideró como RH al observar manchas color café oscuro y secas, mientras que se consideró una reacción S cuando en la hoja se observó flacidez y acuosidad. Los resultados se cotejaron con la información del Cuadro 4 (Stall et al., 2009).
Cuadro 4 Clasificación de razas fisiológicas de Xanthomonas en jitomate por reacciones de susceptibilidad e hipersensibilidad.
| Strain | Municipality | Crop | HA | PCR | REA | Race P | Race T |
|---|---|---|---|---|---|---|---|
| 1 | Ahome | Jitomate | + | X. e | A | P0 | T3 |
| 2 | Culiacán | Jitomate | - | X. v | B | - | T3 |
| 3 | Elota | Chile | - | X. e | A | P8 | T1 |
| 4 | Guasave | Jitomate | + | X. e | A | - | T1 |
| 5 | Escuinapa | Jitomate | + | X. p | C | - | T1 |
| 6 | Navolato | Jitomate | + | X. e | A | - | T1 |
| 7 | Culiacán | Jitomate | + | X. e | A | - | T1 |
| 8 | Ahome | Jitomate | + | X. e | A | - | T1 |
| 9 | Guasave | Jitomate | + | X. e | A | - | T1 |
| 10 | Guasave | Jitomate | - | X. e | A | - | T1 |
| 11 | Escuinapa | Jitomate | + | X. e | A | - | T1 |
| 12 | Escuinapa | Jitomate | + | X. e | A | - | T1 |
| 13 | Culiacán | Jitomate | + | X. e | A | - | T1 |
| 14 | Ahome | Jitomate | + | X. e | A | - | T1 |
| 15 | Escuinapa | Jitomate | + | X. e | A | - | T1 |
| 16 | Navolato | Jitomate | - | X. e | A | - | T1 |
| 17 | Ahome | Jitomate | + | X. e | A | - | T1 |
| 18 | Escuinapa | Jitomate | + | X. e | A | - | T1 |
| 19 | Culiacán | Jitomate | - | X. v | B | - | T1 |
| 20 | Escuinapa | Jitomate | + | X. e | A | - | T1 |
| 21 | Guasave | Jitomate | + | X. e | A | - | T1 |
| 22 | Escuinapa | Chile | + | X. e | A | - | T1 |
| 23 | Ahome | Jitomate | + | X. e | A | - | T1 |
| 24 | Navolato | Jitomate | + | X. p | C | - | T1 |
| 25 | Elota | Jitomate | - | X. e | A | - | T1 |
| 26 | Escuinapa | Jitomate | + | X. e | A | - | T1 |
| 27 | Navolato | Chile | + | X. e | A | P0 | - |
| 28 | Escuinapa | Jitomate | + | X. e | A | P8 | T3 |
| 29 | Culiacán | Jitomate | + | X. e | A | - | T2 |
| 30 | Escuinapa | Jitomate | + | X. e | A | - | T1 |
| 31 | Guasave | Jitomate | - | X. e | A | - | T1 |
| 32 | Culiacán | Chile | + | X. p | C | P0 | - |
| 33 | Elota | Chile | + | X. p | C | P3 | T1 |
| 34 | Ahome | Chile | + | X. e | A | P6 | T1 |
| 35 | Guasave | Chile | + | X. p | C | P10 | T3 |
| 36 | Navolato | Jitomate | + | X. e | A | - | T3 |
| 37 | Culiacán | Jitomate | - | X. e | A | - | T3 |
| 38 | Guasave | Jitomate | + | X. e | A | P8 | T1 |
| 39 | Navolato | Jitomate | - | X. e | A | P8 | T1 |
| 40 | Elota | Jitomate | - | X. e | A | P8 | T1 |
| 41 | Culiacán | Chile | - | X. e | A | - | - |
| 42 | Ahome | Jitomate | - | X. e | A | - | T1 |
| 43 | Navolato | Jitomate | + | X. e | A | P8 | T3 |
| 44 | Escuinapa | Jitomate | - | X. e | A | - | T5 |
| 45 | Ahome | Jitomate | - | X. e | A | - | T5 |
| 46 | Elota | Jitomate | + | X. e | A | - | T5 |
| 47 | Culiacán | Jitomate | - | X. v | B | - | T5 |
Resultados y discusión
Los síntomas de mancha bacteriana se observaron en las hojas de chile y jitomate al momento de recolección del material vegetal en todos los sitios que se visitaron. La incidencia de la enfermedad fue de 5-10% y de 15-25% en campos comerciales con chile y jitomate, respectivamente.
Las pruebas realizadas mostraron que, de las 93 cepas aisladas, solo 47, presentaron las características morfológicas de colonias redondas, convexas, amarillas y mucoides en medio YDC; además, de que fueron Gram y oxidasa negativas. Estas características morfológicas y bioquímicas coincidieron con las reportadas por Schaad et al. (2001) para el caso de Xanthomonas spp. Cabe resaltar que, 32 de las 47 cepas, registraron amilasa negativa.
Igualmente, de las 93 cepas aisladas, solo 47 indujeron RH en plantas de tabaco a las 24 h después de la inoculación y con base en las pruebas bioquímicas, fisiológicas y de patogenicidad, se confirmó que las 47 cepas bacterianas pertenecieron al género Xanthomonas.
Fue posible realizar la identificación de tres especies de Xanthomonas spp. relacionadas con chile y jitomate utilizando los iniciadores específicos para cada especie (Cuadro 5). Todos los productos de PCR punto final se visualizaron en un gel de agarosa como el ejemplo que se muestra en la Figura 1.
El análisis molecular donde se utilizaron iniciadores específicos confirmó que, el 83% de las 47 cepas de Xanthomonas obtenidas de campos de jitomate y chile en Sinaloa (Cuadro 5), correspondió a X. euvesicatoria, el 10.6% a X. perforans y el 6.3% a X. vesicatoria. De manera similar, Lue et al. (2010) reportaron a estas tres especies de Xanthomonas asociadas a jitomate y chile en Taiwán, y tampoco detectaron a X. gardneri.
Cuadro 5 Especies y razas de Xanthomonas aisladas de chile y jitomate en el estado de Sinaloa durante noviembre de 2016 a Marzo de 2017.
| Strain | Municipality | Crop | HA | PCR | REA | Race P | Race T |
|---|---|---|---|---|---|---|---|
| 1 | Ahome | Jitomate | + | X. e | A | P0 | T3 |
| 2 | Culiacán | Jitomate | - | X. v | B | - | T3 |
| 3 | Elota | Chile | - | X. e | A | P8 | T1 |
| 4 | Guasave | Jitomate | + | X. e | A | - | T1 |
| 5 | Escuinapa | Jitomate | + | X. p | C | - | T1 |
| 6 | Navolato | Jitomate | + | X. e | A | - | T1 |
| 7 | Culiacán | Jitomate | + | X. e | A | - | T1 |
| 8 | Ahome | Jitomate | + | X. e | A | - | T1 |
| 9 | Guasave | Jitomate | + | X. e | A | - | T1 |
| 10 | Guasave | Jitomate | - | X. e | A | - | T1 |
| 11 | Escuinapa | Jitomate | + | X. e | A | - | T1 |
| 12 | Escuinapa | Jitomate | + | X. e | A | - | T1 |
| 13 | Culiacán | Jitomate | + | X. e | A | - | T1 |
| 14 | Ahome | Jitomate | + | X. e | A | - | T1 |
| 15 | Escuinapa | Jitomate | + | X. e | A | - | T1 |
| 16 | Navolato | Jitomate | - | X. e | A | - | T1 |
| 17 | Ahome | Jitomate | + | X. e | A | - | T1 |
| 18 | Escuinapa | Jitomate | + | X. e | A | - | T1 |
| 19 | Culiacán | Jitomate | - | X. v | B | - | T1 |
| 20 | Escuinapa | Jitomate | + | X. e | A | - | T1 |
| 21 | Guasave | Jitomate | + | X. e | A | - | T1 |
| 22 | Escuinapa | Chile | + | X. e | A | - | T1 |
| 23 | Ahome | Jitomate | + | X. e | A | - | T1 |
| 24 | Navolato | Jitomate | + | X. p | C | - | T1 |
| 25 | Elota | Jitomate | - | X. e | A | - | T1 |
| 26 | Escuinapa | Jitomate | + | X. e | A | - | T1 |
| 27 | Navolato | Chile | + | X. e | A | P0 | - |
| 28 | Escuinapa | Jitomate | + | X. e | A | P8 | T3 |
| 29 | Culiacán | Jitomate | + | X. e | A | - | T2 |
| 30 | Escuinapa | Jitomate | + | X. e | A | - | T1 |
| 31 | Guasave | Jitomate | - | X. e | A | - | T1 |
| 32 | Culiacán | Chile | + | X. p | C | P0 | - |
| 33 | Elota | Chile | + | X. p | C | P3 | T1 |
| 34 | Ahome | Chile | + | X. e | A | P6 | T1 |
| 35 | Guasave | Chile | + | X. p | C | P10 | T3 |
| 36 | Navolato | Jitomate | + | X. e | A | - | T3 |
| 37 | Culiacán | Jitomate | - | X. e | A | - | T3 |
| 38 | Guasave | Jitomate | + | X. e | A | P8 | T1 |
| 39 | Navolato | Jitomate | - | X. e | A | P8 | T1 |
| 40 | Elota | Jitomate | - | X. e | A | P8 | T1 |
| 41 | Culiacán | Chile | - | X. e | A | - | - |
| 42 | Ahome | Jitomate | - | X. e | A | - | T1 |
| 43 | Navolato | Jitomate | + | X. e | A | P8 | T3 |
| 44 | Escuinapa | Jitomate | - | X. e | A | - | T5 |
| 45 | Ahome | Jitomate | - | X. e | A | - | T5 |
| 46 | Elota | Jitomate | + | X. e | A | - | T5 |
| 47 | Culiacán | Jitomate | - | X. v | B | - | T5 |
HA= Hidrólisis de Almidón; Raza P= Raza en Chile; Raza T= Raza en jitomate; X. e.= Xanthomonas euvesicatoria, X. v.= Xanthomonas vesicatoria, X. p.= Xanthomonas perforans.
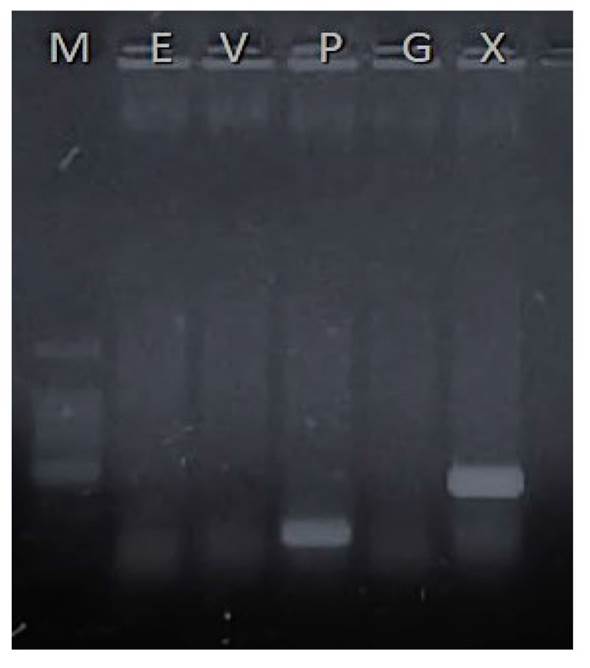
Figura 1 Gel de agarosa para detección de especies de Xanthomonas spp. En el primer carril (M) marcador de 100 pb, segundo, iniciadores para X. euvesicatoria (E), tercero, X. vesicatoria (V), cuarto carril con banda específica para X. perforans (P), el quinto X. gardneri (G) y en sexto carril con iniciadores universales para Xanthomonas spp. (X).
En estudios previos en México, se reportó a X. vesicatoria como la especie predominante en campos con jitomate y chile en los estados de Sinaloa (Bouzar et al., 1996; Carrillo-Fasio et al., 2001) y Sonora (Bouzar et al., 1996). Esto contrastó con nuestro estudio en el cual, X. euvesicatoria fue la especie más frecuente en campos de Sinaloa; mientras que, X. perforans y X. vesicatoria fueron las especies menos frecuentes (Cuadro 5). Esta variación puede explicarse por el continuo movimiento de semilla contaminada entre países (Abbasi et al., 2015; Jibrin et al., 2022).
Para el caso de las ocho cepas obtenidas de plantas de chile en campos de Sinaloa, el 62.5% correspondió a X. euvesicatoria. Esto coincidió con estudios realizados en Australia (Roach et al., 2018), Tailandia (Sitthitanasin et al., 2020) y Bulgaria (Bogatzevska et al., 2021), donde X. euvesicatoria fue la especie predominante en el cultivo de chile.
En este estudio, X. euvesicatoria fue la especie más frecuente (87.1%) en las 39 cepas analizadas de campos de jitomate en Sinaloa. Similarmente, esta especie se ha reportado en campos con jitomate en Taiwán (Lue et al., 2010), Canadá (Abbasi et al., 2015), Nigeria (Jibrin et al., 2018), Bulgaria (Bogatzevska et al., 2021) y medio oeste de los EE. UU. (Rotondo et al., 2022).
Xanthomonas perforans fue la segunda especie más común en nuestro estudio con un 10.6% de frecuencia en campos con jitomate y chile en Sinaloa. Esta especie se detectó previamente en campos con jitomate en Taiwán (Lue et al., 2010), Canadá (Abbasi et al., 2015), Australia (Roach et al., 2018), Tailandia (Sitthitanasin et al., 2020) y medio oeste de los EE. UU. (Rotondo et al., 2022),
Xanthomonas vesicatoria fue la especie menos frecuente (6.4%) de las cepas aisladas de campos con jitomate y chile en Sinaloa, coincidiendo ampliamente con el estudio de Lue et al. (2010), quienes encontraron a esta especie en menor frecuencia y menos distribuida en campos de jitomate en Taiwán en comparación con X. euvesicatoria y X. perforans. Por el contrario, Bogatzevska et al. (2021) señalaron que X. vesicatoria fue la especie más frecuente y ampliamente distribuida en el mismo cultivo, en Bulgaria.
De las cepas de X. euvesicatoria que se aislaron de campos con chile en Sinaloa, se identificaron las razas P0, P3, P6, P8 y P10. De manera similar, estas razas también se han detectado en campos con chile en Bulgaria (Bogatzevska et al., 2021). Mientras que, de las cepas aisladas de jitomate, se encontraron que seis fueron capaces de atacar a plantas de chile siendo las razas P0 y P8.
Comparando con los resultados previos obtenidos por Carrillo-Fasio et al. (2001), en el estudio de razas de Xanthomonas spp. en Sinaloa, se observó que después de más de 15 años; las razas P1 y P2 no se encontraron en este estudio debido a probablemente a que no están presentes en las zonas de producción muestreadas o que la fecha de muestreo no coincidió con la presencia del patógeno. Entretanto, las razas P6 y P10 se detectaron por primera vez; mientras que, las razas P0 y P3 continúan en la población de Xanthomonas spp. Las razas que predominaron en Sinaloa fueron P0, P6 y P8, a diferencia del año 2001, en el cual la raza predominante fue la P3.
En el cultivo de jitomate en campos de Sinaloa, se detectaron las razas TI, T2, T3 y T5 de Xanthomonas. Con base en datos de distribución y frecuencia de las cepas, al norte del estado de Sinaloa (Ahome y Guasave) se encuentran las razas T1 y T3; en el centro (Culiacán, Navolato y Elota) las razas T1, T2 y T3; al sur (Escuinapa) las razas T1, T2, T3 y T5, siendo las razas T1 y T3, las predominantes en todo el estado (Figura 2). En el caso de chile, las razas de Xanthomonas spp. se distribuyeron de la siguiente manera: al norte (Ahome y Guasave) se encontraron las razas P0, P6, P8 y P10; en el centro (Culiacán, Navolato y Elota) las razas P0, P3, P6 y P8; al sur (Escuinapa) las razas P0, P6 y P8; siendo las razas P0, P6 y P8 las predominantes en todo el estado (Figura 2).
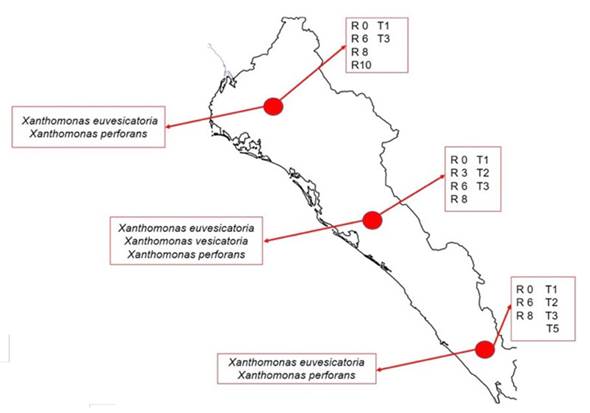
Figura 2 Distribución de especies y razas de Xanthomonas aisladas de jitomate y chile en campos distribuidos en el estado de Sinaloa, México durante el ciclo 2016-2017.
Además de la identificación de Xanthomonas spp. mediante iniciadores específicos, existen algunos estudios que han analizado secuencias de ADN y realizado análisis filogenéticos multilocus para la distinción de las diversas especies de Xanthomonas que infectan a jitomate y chile en diversos países (Abbasi et al., 2015; Schwartz et al., 2015; Timilsina et al., 2015; Roach et al., 2018). Por lo que, se sugiere realizar posteriores estudios filogenéticos con secuencias de ADN de Xanthomonas spp. para obtener mayor información sobre su origen y diversidad genética en el estado de Sinaloa y en otros estados productores de jitomate y chile en México.
Conclusiones
La información generada en este estudio confirmó que en campos con jitomate en el estado de Sinaloa se encuentran las especies X. euvesicatoria, X. perforans y X. vesicatoria. Mientras que, en campos con chile se detectó a X. euvesicatoria y X. perforans. Además, se registró la presencia de las razas T1, T2, T3 y T5 para jitomate; así como, las razas P0, P3, P6, P8 y P10 para chile. Este es el primer reporte de la presencia de X. perforans en México. Por lo que se requiere de estudios posteriores para depositar secuencias representativas de cada especie de Xanthomonas en el GenBank, conocer la sensibilidad de estas cepas a bactericidas; además de, generar datos sobre epidemiología y manejo de estas bacterias fitopatógenas en México. Asimismo, es importante resaltar que no se detectó la presencia de X. gardneri.
Literature cited
Abbasi PA, Khabbaz SE, Weselowski B and Zhang L. 2015. Occurrence of copper-resistant strains and a shift in Xanthomonas spp. causing tomato bacterial spot in Ontario. Canadian Journal of Microbiology 61: 1-9. https://doi.org/10.1139/cjm-2015-0228 [ Links ]
Adhikari P, Adhikari TB, Louws FJ and Panthee DR. 2020. Advances and challenges in bacterial spot resistance raceing in tomato (Solanum lycopersicum L.). International Journal of Molecular Sciences 21: 1734. https://doi.org/10.3390/ijms21051734 [ Links ]
Bogatzevska N, Vancheva-Ebben T, Vasileva K, Kizheva Y and Moncheva P. 2021. An overview of the diversity of pathogens causing bacterial spot on tomato and chili pepper in Bulgaria. Bulgarian Journal of Agricultural Science 27(1): 137-146. https://agrojournal.org/27/01-19.pdf [ Links ]
Bouzar H, Jones JB, Stall RE, Hodge NC, Minsavage GV, Benedict AA and Alvarez AM. 1994. Physiological, chemical, serological, and pathogenic analyses of worldwide collection of Xanthomonas campestris pv. vesicatoria strains. Phytopathology 31: 753-755. https://doi.org/10.1094/Phyto-84-663 [ Links ]
Bouzar H, Jones JB, Somodi GC, Stall RE, Daouzli N, Lambe RC, Felix-Gastelum R and Trinidad-Correa R. 1996. Diversity of Xanthomonas campestris pv. vesicatoria in tomato and chili pepper fields of Mexico. Canadian Journal of Plant Pathology 18: 75-77. http://dx.doi.org/10.1080/07060669609500659 [ Links ]
Carrillo-Fasio JA, Sánchez-Bautista L, García-Estrada RS, Allende-Molar R y Márquez-Zequera I. 2001. Razas de Xanthomonas campestris pv. vesicatoria (Doidge) Dye. presentes en el estado de Sinaloa, México. Revista Mexicana de Fitopatología 19: 248-250. https://www.redalyc.org/pdf/612/61219219.pdf [ Links ]
Hartman GL and Yang CH. 1990. Occurrence of three races of Xanthomonas campestris pv. vesicatoria on chili pepper and tomato in Taiwan. Plant Disease 74: 252. [ Links ]
Jibrin MO, Potnis N, Timilsina S, Minsavage GV, Vallad GE, Roberts PD, Jones GB and Goss EM. 2018. Genomic inference of recombination-mediated evolution in Xanthomonas euvesicatoria and X. perforans. Applied and Environmental Microbiology 84: e136-e118. https://doi.org/10.1128/AEM.00136-18 [ Links ]
Jibrin MO, Timilsina S, Minsavage GV, Vallad GE, Roberts PD, Goss EM and Jones JB. 2022. Bacterial spot of tomato and chili pepper in Africa: diversity, emergence of T5 race, and management. Frontiers in Microbiology 13:835647. https://doi.org/10.3389/fmicb.2022.835647 [ Links ]
Jones JB, Lacy GH, Bouzar H, Stall RE and Schaad NW. 2004. Reclassification of Xanthomonads associated with bacterial spot disease of tomato and chili pepper. Systematic and Applied Microbiology 27: 755-762. https://doi.org/10.1078/0723202042369884 [ Links ]
Király Z, Kelman Z, Solymosy L and Voros J. 1974. Methods in Plant Pathology. With special reference to raceing for disease resistance. Elsevier Scientific Publishing Company. New York. pp. 153-166. [ Links ]
Koike ST, Gladders P and Paulus AO. 2007. Vegetable Diseases: A colour Handbook. Manson Publishing, London, UK. 448 p. [ Links ]
Kousik CS and Ritchie DF. 1995. Isolation of chili pepper races 4 and 5 of Xanthomonas campestris pv. vesicatoria from diseased chili peppers in Southeastern U.S. fields. Plant Disease 79: 540. [ Links ]
Lue YS, Deng WL, Wu YF, Cheng AS, Hsu ST and Tzeng KC. 2010. Characterization of Xanthomonas associated with bacterial spot of tomato and chili pepper in Taiwan. Plant Pathology Bulletin 19: 181-190. [ Links ]
Obradovic A, Mavridis A, Rudolph K, Janse JD, Arsenijevic M, Jones JB, Minsavage GV and Wang JF. 2004. Characterization and PCR-based typing of Xanthomonas campestris pv. vesicatoria from chili peppers and tomatoes in Serbia. European Journal of Plant Pathology 110: 285-292. https://doi.org/10.1023/B:EJPP.0000019797.27952.1d [ Links ]
O’Garro LW. 1998. Bacterial spot of tomato and chili pepper on four East Caribbean islands: Races, their abundance, distribution, aggressiveness, and prospects for control. Plant Disease 82: 864-870. [ Links ]
Potnis N, Timilsina S, Strayer A, Shantharaj D, Barak JD, Paret ML, Vallad GE, and Jones JB. 2015. Bacterial spot of tomato and chili pepper: diverse Xanthomonas species with a wide variety of virulence factors posing a worldwide challenge. Molecular Plant Pathology 16(9): 907-920. https://doi.org/10.1111/mpp.12244 [ Links ]
Quezado-Duval AM e Camargo LEA. 2004. Raças de Xanthomonas spp. associadas à mancha-bacteriana em tomate para processamento industrial no Brasil. Horticultura Brasileira 22(1):80-86. [ Links ]
Roach R, Mann R, Gambley CG, Shivas RG and Rodoni B. 2018. Identification of Xanthomonas species associated with bacterial leaf spot of tomato, capsicum and chilli crops in eastern Australia. European Journal of Plant Patholology 150: 595-608. https://doi.org/10.1007/s10658-017-1303-9 [ Links ]
Rotondo F, Bernal E, Ma X, Lewis Ivey ML, Sahin F, Francis DM and Miller SA. 2022. Shifts in Xanthomonas spp. causing bacterial spot in processing tomato in the Midwest of the United States. Canadian Journal of Plant Pathology. https://doi.org/10.1080/07060661.2022.2047788 [ Links ]
Schaad NW, Jones JB and Chun W. 2001. Laboratory guide for identification of plant pathogenic bacteria. American Phytopathological Society. St. Paul. Minnesota, USA. [ Links ]
Schwartz AR, Potnis N, Timilsina S, Wilson M, Patané J, Martins JJr, Minsavage GV, Dahlbeck D, Akhunova A, Almeida N, Vallad GE, Barak JD, White FF, Miller SA, Ritchie D, Goss E, Bart RS, Setubal JC, Jones JB and Staskawicz BJ. 2015. Phylogenomics of Xanthomonas field strains infecting chili pepper and tomato reveals diversity in effector repertoires and identifies determinants of host specificity. Frontiers in Microbiology 6:535. https://doi.org/10.3389/fmicb.2015.00535 [ Links ]
Sharma S and Bhattarai K. 2019. Progress in developing bacterial spot resistance in tomato. Agronomy 9: 26. https://doi:10.3390/agronomy9010026 [ Links ]
Stall RE, Jones JB and Minsavage GV. 2009. Durability of resistance in tomato and chili pepper to xanthomonads causing bacterial spot. Annual Review of Phytopathology 47: 265-284. https://doi.org/10.1146/annurev-phyto-080508-081752 [ Links ]
Sitthitanasin S, Korakngam C, Kanhayart T, Kositcharoenkul N, Patarapuwadol S, Kositratana W and Watcharachaiyakup J. 2020. Characterization of Xanthomonas causing of bacterial leaf spot of tomato and chili pepper in Thailand. Thai Agricultural Research Journal 38(1): 80-89. https://doi.org/10.14456/thaidoa-agres.2020.7 [ Links ]
Timilsina S, Jibrin MO, Potnis N, Minsavage GV, Kebede M, Schwartz A, Bart R, Staskawicz B, Boyer C, Vallad GE, Pruvost O, Jones JB and Goss EM. 2015. Multilocus sequence analysis of xanthomonads causing bacterial spot of tomato and chili pepper plants reveals strains generated by recombination among species and recent global spread of Xanthomonas gardneri. Appl Environmental Microbiology 81:1520-1529. https://doi.org/10.1128/AEM.03000-14 [ Links ]
Recibido: 29 de Octubre de 2022; Aprobado: 12 de Mayo de 2023
 This is an open-access article distributed under the terms of the
Creative Commons Attribution License
This is an open-access article distributed under the terms of the
Creative Commons Attribution License
