










Services on Demand
Journal
Article
 text in
text in  English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista mexicana de fitopatología
On-line version ISSN 2007-8080Print version ISSN 0185-3309
Rev. mex. fitopatol vol.35 n.2 Texcoco May. 2017
https://doi.org/10.18781/r.mex.fit.1608-2
Phytopathological notes
Comparison of enzymes and phenolic compounds in three citrus species infected with Candidatus Liberibacter asiaticus
1Universidad Autónoma Agraria Antonio Narro, Departamento de Parasitología Agrícola, Buenavista, Saltillo, Coahuila, México. C.P. 25315.
2Campo Experimental INIFAP, Carretera Montemorelos-China Km. 31, Colonia Ex Hacienda Las Anacuas General Terán C.P. 67413, General Terán Nuevo León.
3Centro de Investigación y Estudios Avanzados Unidad Irapuato, Instituto Politécnico Nacional, km 9.6 Libramiento Norte carretera. Irapuato-León, Irapuato, Guanajuato, CP 36821, México.
4Universidad Autónoma Agraria Antonio Narro, Departamento de Horticultura, Buenavista, Saltillo, Coahuila, México. C.P. 25315.
5CONACYT-Universidad Autónoma Agraria Antonio Narro, Buenavista, Saltillo, Coahuila, México. C.P. 25315.
6Universidad Autónoma Agraria Antonio Narro, Departamento de Fitomejoramiento, Buenavista, Saltillo, Coahuila, México. C.P. 25315.
The objective was to determine differences in antioxidant enzyme activity and total phenol concentration in Mexican lime plants (Citrus aurantifolia (Christm.) Swingle), Persian lime (Citrus latifolia Tanaka); and Valencia sweet orange (Citrus sinensis (L.) Osbeck). Infected under natural field conditions with Candidatus Liberibacter asiaticus (CaLas), in Bustamante, Tamaulipas, Mexico. The presence of CaLas was determined by qPCR. We determined the enzymatic activity of phenylalanine ammonia lyase (PAL), which is activated by pathogen attack; the peroxidase involved in wall lignification in response to infection; the α-amylase which degrades starch and accumulates in vascular tissue; and phenolic compounds involved in defense functions. A contrasts analysis was performed. Total protein concentration showed significant differences between species (P<0.0001). With a mean of 6.1 and 6.37 mg of protein per g of fresh tissue for infected Mexican lime trees, and sweet orange negative CaLas. The peroxidase activity presented a significant difference for Persian lime (P=0.0341), with a mean of 1.96 U.mg of protein-1. In sweet orange, higher α-amylase activity was observed in CaLas infected trees (1.19 U.mg protein-1). The concentration of PAL and total phenols did not show significant differences between species. In the present study it was observed that CaLas influences the enzymatic activity of Citrus species.
Key words: Huanglongbing; Phenylalanine ammonia lyase; peroxidase; alpha amylase; total proteins; total phenols
El objetivo fue determinar diferencias en la actividad enzimática antioxidante y la concentración de fenoles totales en plantas de limón mexicano (Citrus aurantifolia (Christm.) Swingle), limón persa (Citrus latifolia Tanaka); y naranja dulce Valencia (Citrus sinensis (L.) Osbeck), infectadas bajo condiciones naturales de campo con Candidatus Liberibacter asiaticus (CaLas), en Bustamante, Tamaulipas, México. Se determinó la presencia de CaLas por qPCR; la actividad enzimática de fenilalanina amonio liasa (PAL), que se activa ante el ataque de patógenos; peroxidasa que interviene en la lignificación de paredes como respuesta a la infección; α-amilasa que degrada el almidón y se acumula en el tejido vascular, y compuestos fenólicos que cumplen funciones de defensa. Se realizó un análisis de contrastes. La concentración de proteínas totales mostró diferencias significativas entre especies (P<0.0001). Con una media de 6.1 y 6.37 mg de proteínas por g de tejido fresco para árboles infectados de limón mexicano; y naranja dulce negativa a CaLas. La actividad peroxidasa presentó diferencia significativa para limón persa (P=0.0341), con una media de 1.96 U.mg de proteína-1. En naranja dulce se observó mayor actividad de α amilasa en los árboles infectados por CaLas (1.19 U.mg de proteína-1). La concentración de PAL y fenoles totales no mostraron diferencias significativas entre especies. En el presente estudio se observó que CaLas influye en la actividad enzimática de cítricos.
Palabras clave: Huanglongbing; Fenilalanina amonio liasa; peroxidasa; α-amilasa; proteínas totales; fenoles totales
Candidatus Liberibacter asiaticus (CaLas), affects the citrus-producing areas in Mexico (DGSV-SENASICA and Mora-Aguilera, 2012); it reduces by 18.6 % the volume of juice and by 17.3 % the weight of fruits in Persian lime in Yucatan, Mexico, causing losses in production of 2.4 t.ha-1 (Flores-Sánchez, et al., 2015). The infection induced by HLB alters the export of photoassymilates caused by the retention of starch, which generates the expression of symptoms in the tree (Kim et al., 2009; Koh et al., 2012).
CaLas causes biochemical and structural changes in citrus plants to prevent the spread of the bacteria, by activating response proteins (Albrecht and Bowman, 2008). In response to the infection of citrus plants with HLB, carbohydrate metabolism involving α-amylase is altered (Albrecht and Bowman, 2008; Etxeberria et al., 2009).
Phenylalanine ammonia lyase (PAL) intervenes in response to the attack of the pathogens (Almario et al., 1994). In citrics, the flavedo has the ability to respond to attacks by pathogens, by the increase of the PAL transcript levels, as well as of its activity (Ballester et al., 2006). The enzyme peroxidase (POD) protects plants from the damage caused by the free radicals or ROS generated by different types of stress and also participates in the lignification of the cell wall and in the degeneration of indolacetic acid (Robinson, 1991). Phenols have proven to play an important part in the defense of plants to different biotic and abiotic factors (Lu et al., 2015). They can be modified by effectors secreted by the bacteria Ca. Liberibacter inside the host, and that alter genes related to defense; CaLas codifies salicylate hydroxylase as a mechanism to evade the plants defenses (Aritua et al., 2013). Mora-Aguilera et al., 2014, indicate that HLB control must consider the vulnerability of the citrus species and the load of the inoculant. Understanding the enzymatic behavior of trees infected with CaLas in different species could help develop alternatives for the protection or defense of the plant. This study was developed with the objective of determining differences in the antioxidant enzyme activity and the concentration of phenols in three citrus species in Bustamante, Tamaulipas, Mexico.
Samples were taken in Felipe Angeles, Bustamante, Tamaulipas, Mexican lime (Citrus aurantifolia (Christm.) Swingle), Persian lime (Citrus latifolia Tanaka) and sweet orange trees (Citrus sinensis (L.) Osbeck) aged 10 to 12 years were sampled under rainfed conditions, in the phenological stage of fruit set. Samples were taken from two trees with symptoms per species, and one without symptoms. The material was gathered in each one of the cardinal points of the tree (north, south, east, and west), three repetitions of five leaves were taken from each point and kept in liquid nitrogen.
For DNA extraction, the technique quoted by Almeyda-León et al. (2001) was modified for its use: 100 mg of tissue were ground in liquid nitrogen, 1 mL was added of 2 ME/CTAB extraction solution, preheated at 65 °C. It was mixed in a vortex for 30 sec and incubated at 65 °C for 45 min. Next, 500 μL of isoamyl alcohol chloroform was added 24:1, the mixture was stirred inversely and centrifuged at 12,000 rpm for 10 min. The aqueous phase was extracted and an equal volume of isoamyl alcohol chloroform was added 24:1; the centrifuge cycle was repeated. The top layer was taken and 0.6 volume of isopropanol was added to precipitate the DNA at -20 °C for 24 h. It was centrifuged at 12,000 rpm for 20 min, the pellet was washed with ethanol at 70 %, centrifuged at 12,000 rpm for 15 min, left to dry and resuspended in 50 μl injectable water (Pisa). The presence of bacteria was determined according to the protocol by SENASICA-SAGARPA, (2010) used in the National Plant Epidemiology and Health Station (SENASICA-ENECUSaV), in the state of Querétaro, Mexico.
The raw extract for the quantification of total proteins, phenylalanine ammonia lyase, peroxidase, and α-amiylase, was taken from Díaz et al. (2010), and modified: 1 g of grinded sample was placed in liquid nitrogen, in a sodium phosphate buffer solution 100 mM, pH 7. The samples were centrifuged at 12,000 rpm for 20 minutes at a temperature of 4 °C; later, the supernatant was collected and stored at -20 °C. For the extraction of phenols, methodology B, used by Kähkönen et al. (1999) was used here with some modifications; 250 mg of the grinded sample were placed in a 2 mL eppendorf tube and 1 mL of methanol at 80 % was added; it was shaken in a vortex for one minute and centrifuged at 10,000 rpm for 15 minutes. The supernatant was collected in a 1.5 mL amber eppendorf tube. It was re-extracted with 500 μL of absolute methanol, repeating the process and stored at -20 °C until use.
Protein concentration was determined according to the technique by Bradford (1976), which consisted in mixing 1 mL of Bradford reagent with 100 μL of raw extract. The total protein absorbance values were determined at a λ=595 nm in a thermo spectronic Biomate 3. The protein content was expressed in mg of protein per mg of fresh tissue. The pattern curve was obtained with the albumen of cow serum, as per García and Vázquez (1998).
Phenylalanine ammonia lyase (PAL) activity was determined according to the technique described by Rodríguez-Pedroso et al., (2006). The 0.0174 M-1cm-1 molar extinction coefficient was used as a reference to calculate enzyme activity (Trotel-Aziz et al., 2008).
Peroxidase activity was determined using the methodology used by Ruttimann et al., (1992). Enzyme activity was expressed as millimoles of phenol red oxidized per gram of fresh tissue per minute (Yedidia et al., 1999).
For the quantification of α amylase, we used the SIGMA-ALDRICH protocol, (2015). The standard curve was produced with various concentrations of maltose. The α amylase activity was defined as µmol.min-1 of maltose released per µg of protein (Menéndez et al., 2006).
The content of phenols was determined with the Folin-Ciocalteu method (Singleton et al., 1999). A mixture was prepared using 200 μL of the methanolic extract, 100 μL Folin-Ciocalteu reagent, and 200 μL of sodium carbonate at 20 %. It was shaken and left to rest for 30 min. After this time, total phenol absorbance was measured at one λ=760 nm. A standard curve was produced with various concentrations of gallic acid.
The SAS 9.0 statistical package was used to perform an analysis of contrasts between the trees with positive and negative diagnoses for CaLas. The variables analyzed were: total proteins, PAL, POD, α-amylase, and total phenols.
The sampled trees with symptoms displayed a general yellowness of the canopy and they appeared dehydrated; leaves presented asymmetric, less visible spots. Symptomatic samples were positive for CaLas, and asymptomatic samples were negative. For the detection of CaLam all samples were negative.
The analysis of contrasts between positive and negative trees displayed significant differences between species (P<0.0001). Protein concentration for Mexican lime displayed an average of 6.1 mg of proteins per g of fresh tissue for infected trees, whereas for negative ones, it was 5.46 mg of proteins per g of fresh tissue with a significance of P=0.0238. In sweet orange, there is a significant difference of P=0.0002 with an average of 5.27 and 6.37 mg of proteins per g of fresh tissue in positive and negative trees, respectively (Table 1; Figure 1). Determining total proteins in the plan helps find changes caused by different types of biotic and abiotic stress (Casado, 2004), as shown in this study when finding differences in the concentration of proteins between positive and negative trees for CaLas.
Table 1. Analysis of contrasts between trees with negative and positive diagnoses for Ca. Liberibacter asiaticus in the state of Tamaulipas.
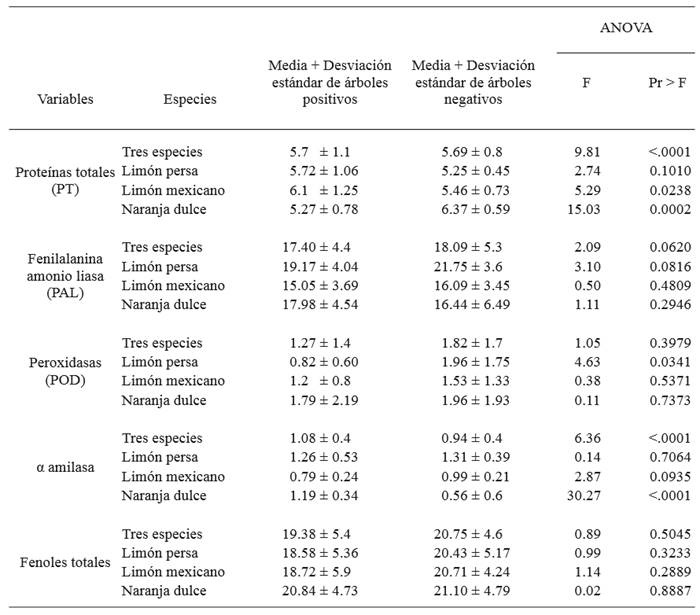
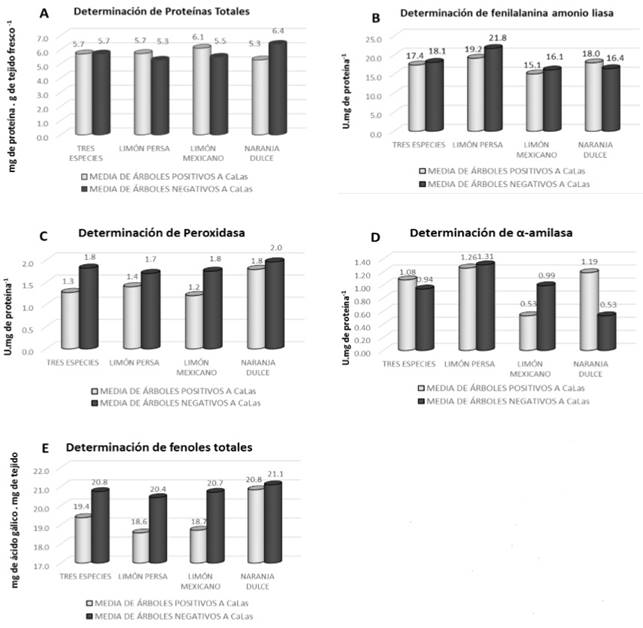
Figure 1. Comparison of averages of total proteins (1A), phenylalanine ammonium lyase (PAL) (1B), peroxidase (1C) α-amylase (1D), and total phenols (1E), for the sources of variations of location, species, and branch orientation evalu ated in trees, both negative and positive for Ca. Liberibacter asiaticus.
Determining PAL did not display significant differences between species, although a higher concentration of PAL was observed in Persian lime and Mexican line (21.75, 16.09 U.mg of protein-1) for negative trees. In contrast, sweet orange trees infected by CaLas showed a higher concentration (17.98 U.mg of protein-1) (Table 1; Figure 1). The results of this study coincided with Chenyang et al. (2001), who mention that the enzyme activity of PAL is modified by biotic and abiotic factors in the plant, since despite having no significant differences, a higher PAL concentration was observed in sweet oranges affected by HLB. The results in Persian and Mexican lime coincided with Martinelli et al. (2016), who point out that the regulation of PAL dropped due to the infection of CaLas in both species, one considered moderately tolerant, Volkameriana lime (Citrus × volkameriana), and the other, Navel orange, highly vulnerable.
The levels of peroxidase activity presented a significant difference for Persian lime (P=0.0341). A higher concentration of the enzyme was observed in trees not infected with CaLas with averages of 1.96 U.mg of protein-1 for Persian lime and sweet orange, and for Mexican lime, 1.53 U.mg of protein -1 (Table 1; Figure 1). In this investigation, we oberved a higher enzyme peroxidase activity in negative citrus trees; however, symptoms of central and secondary veins degeneration in leaves leaves infected with HLB, produced by the induced lignification by this enzyme, as mentioned Robinson (1991).
In the enzyme quantification of α-amylase, significant differences were presented between species and between positive and negative sweet orange trees (P<0.0001). Negative Persian lime and Mexican lime trees displayed a higher concentration of α-amylase (1.31, 0.99 U.mg of protein-1), respectively; in positive trees, the highest concentration was found in Persian lime (1.26 U.mg of protein-1) (Table 1; Figure 1). In the study by Martinelli et al. (2016), α-amylase was expressed in Volkameriana lime (Citrus × volkameriana), indicated as moderately tolerant, and β-amylase in Navel orange, a highly vulnerable material. In this study, the stress on the tree caused the α-amylase activity to increase in Persian lime, followed by sweet orange.
The levels of total phenols were displayed in a minimum range of 18.5 mg of gallic acid per g of fresh weight registered for the samples of Persian lime infected with CaLas. Sweet orange presented the maximum value (21.10 mg of gallic acid per g of fresh weight) in negative trees. In the analysis of contrasts, the determination of total phenols did not show significant differences between species (P=0.5045) (Table 1; Figure 1). Coinciding with Yedidia et al. (1999), who point out that when suppressing the peroxidase activity, the production of phenolic compounds is also reduced, the results do not display a significant difference in peroxidase activity or in phenol compounds.
Conclusions
The study showed a higher concentration of proteins in Mexican lime with CaLas, and sweet orange in the negative ones. There was a higher PAL enzyme activity in sweet orange tested positive for CaLas than in negative trees. The activity of the enzyme peroxidase was not perceivable in this stage of the disease, as was the case for the concentration of total phenolic compounds. The activity of α-amylase was higher in Persian lime.
Acknowledgements
To the UAAAN and the CONACYT scholarship program for the funds granted. To the Tamaulipas State Plant Health Committee for the logistical support provided.
REFERENCES
Albrecht U. and Bowman K. D. 2008. Gene expression in Citrus sinensis (L.) Osbeck following infection with the bacterial pathogen Candidatus Liberibacter asiaticus causing Huanglongbing in Florida. Plant Science. 175: 291-306. http://dx.doi.org/10.1016/j.plantsci.2008.05.001 [ Links ]
Almario, M. F., Ocampo C. y Montes de G. V. 1994. Establecimiento de un método para la determinación de la actividad enzimática de L Fenilalanina amonio liasa (PAL) en Cafeto. Revista de la Academia Colombiana de Ciencias. 19 (72): 137-140. http://www.accefyn.org.co/revista/Vol_19/72/137-140.pdf [ Links ]
Almeyda-León. I. H., Rocha-Peña. M. A., Piña-Razo. J. and Martínez-Soriano. J. P. 2001. The use of polymerase chain reaction and molecular hybridization for detection of phytoplasmas in different plant species in Mexico. Revista Mexicana de Fitopatología 19:1-9. http://www.redalyc.org/pdf/612/61219101.pdf [ Links ]
Arfaoui A., El Hadrami A., Mabrouk Y., et al. 2007. Treatment of chickpea with Rhizobium isolates enhances the expresión of phenylpropanoid defense-related genes in response to infection by Fusarium oxysporum sp. Ciceris. Plant Physiology and Biochemistry 45 (6-7): 470-479. http://dx.doi.org/10.1016/j.plaphy.2007.04.004 [ Links ]
Aritua V, Achor D, Gmitter F. G., Albrigo G., Wang N. 2013. Transcriptional and microscopic analyses of citrus stem and root responses to Candidatus Liberibacter asiaticus infection. PLoS ONE 8: doi: 10.1371/journal.pone.0073742 [ Links ]
Ballester A. R., Lafuente M. T., González-Candelas L. 2006. Spatial study of antioxidant enzymes, peroxidase and phenylalanine ammonia-lyase in the citrus fruit- Penicillium digitatum interaction. Postharvest Biology and Technology 39: 115-124. http://dx.doi.org/10.1016/j.postharvbio.2005.10.002 [ Links ]
Bernal L., Martínez B. E. 2006. Una nueva visión de la degradación del almidón. Revista del Centro de Investigación. Universidad La Salle. 7(25): 77-90. http://www.redalyc.org/articulo.oa?id=34202506 [ Links ]
Bradford, M. M. 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Analytical Biochemistry. 72(1/2): 248-254. doi:10.1016/0003-2697(76)90527-3 [ Links ]
Casado V. J. 2004. Aproximación cinética, molecular y proteómica al studio de la podredumbre apical en frutos de tomate (Lycopersicon esculentum M.): implicación de polifenol oxidase (PPO) y enzimas antioxidants. Tesis de Doctorado. Universidad de Alicante. P-113 http://hdl.handle.net/10045/9919 [ Links ]
Chenyang H., Hsiang T., Wolyn D. 2001. Activation of defense responses to Fusarium infection in Asparagus densiflorus. European Journal of Plant Pathology 107:473-483. http://link.springer.com/article/10.1023/A:1011218304331 [ Links ]
Christopoulos M. V. and Tsantili E. 2015. Participation of phenylalanine ammonia-lyase (PAL) in increased phenolic compounds in fresh col stressed walnut. (Juglans regia L.) Kernels. Postharvest Biology and Technology 104: 17-25 http://dx.doi.org/10.1016/j.postharvbio.2015.03.003 [ Links ]
Díaz M., Pérez Y., Cazaña Y., Prieto M., Wencomo H. y Lugo Y. 2010. Determinación de antioxidantes enzimáticos en variedades e híbridos de Morus alba. Pastos y Forrajes 3 (3) http://scielo.sld.cu/pdf/pyf/v33n3/pyf06310.pdf [ Links ]
DGSV-Senasica y Mora-Aguilera G. 2012. Ficha técnica HLB Huanglongbing. 31 p. http://senasica.gob.mx/?id=4612 [ Links ]
Etxeberria E., Gonzalez P., Achor D. and Albrigo G. 2009. Anatomical distribution of abnormally high levels of starch in HLB-affected valencia orange trees. Physiological and Molecular Plant Pathology. 74: 75-83 DOI:10.1016/j.pmpp.2009.09.004 [ Links ]
Flores-Sánchez J. L, Mora-Aguilera G., Loeza-Kuk E, López-Arroyo. J. I., Domínguez-Monge S, Acevedo-Sánchez. G. y Robles-García P. 2015. Pérdidas en producción inducidas por Candidatus Liberibacter asiaticus en limón persa en Yucatán, México. Revista Mexicana de Fitopatología 33: 195-210. http://www.redalyc.org/articulo.oa?id=61242145005 [ Links ]
Folimonova S. Y., Robertson C. J., Gansey S. M, Gowda S. and Dawson O. 2009. Examination of the responses of different genotype of citrus to Huanglongbing (Citrus Greening) under different conditions. Phytopathology. 99 (12):1346-1354. http://apsjournals.apsnet.org/doi/pdfplus/10.1094/PHYTO-99-12-1346 [ Links ]
García A. H. y Vázquez D. R. 1998. Cuantificación de proteínas: una revisión. Bitácora. Biotecnología 3: 77-88. www.smbb.com.mx/revista/Revista_1998_2/bitacora.pdf [ Links ]
Kähkönen, M.P., Hopia A. I., Heikki J. V., Jussi P. R., Pihlaja K., Kujala T. S., and Heinonene M. 1999. Antioxidant activity of plant extracts containing phenolic compounds. Journal of Agricultural and Food Chemistry 47:3954-3962 http://pubs.acs.org/doi/abs/10.1021/jf990146l [ Links ]
Kim J.S., Sagaram U. S., Burns J.K., Li J.L., Wang N. 2009 Response of sweet orange (Citrus sinensis) to ‘Candidatus Liberibacter asiaticus’ infection: microscopy and microarray analyses. Phytopathology 99:50-57 http://dx.doi.org/10.1094/PHYTO-99-1-0050 [ Links ]
Koh E-J., Zhou L., Williams D. S., Park J., Ding N., Duam J-P. and Kang B-H. 2012. Callose deposition in the phloem plasmodesmata and inhibition of phloem transport in citrus leaves infected with “Candidatus Liberibacter asiaticus”. Protoplasma. 249: 687-697. DOI: 10.1007/s00709-011-0312-3 [ Links ]
Lozoya S. H., Rivera H. R. y Colinas-León. M. T. 2007. Fenoles, peroxidasa y fenilalanina amonio-lyasa: su relación con la resistencia genética de clones de papa (Solanum tuberosum L.) contra el tizón tardío (Phytophthora infestans Mont. De Bary). Agrociencia 41: 479-489 https://dialnet.unirioja.es/servlet/articulo?codigo=2284649 [ Links ]
Lu L., Wang J., Zhu R., Lu H., Zheng X. and Yu T. 2015. Transcript profiling analysis of Rhodosporidium paludigenum mediated signalling pathways and defense responses in mandarin orange. Food Chemistry. 172: 603-612. http://dx.doi.org/10.1016/j.foodchem.2014.09.097 [ Links ]
Martinelli F., Reagan R. L., Dolan D., Fileccia V. and Dandekar A. M. 2016. Proteomic analysis highlights the role of detoxification pathways in increased tolerance to Huanglongbing disease. BMC Plant Biology 16:167 DOI 10.1186/s12870-016-0858-5 [ Links ]
Menéndez A. O., Evangelista L. S., Arenas O. M., Bermúdez T. K., Del Villar M. A., Jiménez A. A. 2006. Cambios en la actividad de α-amilasa, pectinmetilesterasa y poligalacturonasa durante la maduración del maracuyá amarillo (Passiflora edulis Var. Flavicarpa Degener). Interciencia. 31(10): 728-733. ISSN 0378-1844 [ Links ]
Mora-Aguilera, G., Robles-García, P., López-Arroyo, J. I., Flores-Sánchez, J., Acevedo-Sánchez G., Domínguez-Monge S., Gutiérrez-Espinosa A., Loeza-Kuk E., González-Gómez R. 2014. Situación actual y perspectivas del manejo del HLB de los cítricos. Revista Mexicana de Fitopatología. 32(2): 108-119. http://www.redalyc.org/articulo.oa?id=61243856003 [ Links ]
Paz-Lago. D., Cabrera G., Ramírez M., Pombo R. y Gutiérrez A. 1999. Influencia de derivados de quitina en la interacción tomate-Fusarium oxysporum f.sp. licopersici a nivel de bioensayo. Cultivos Tropicales 20:59-66. http://www.sidalc.net/cgi-bin/wxis.exe/?IsisScript=ctrop.xis&method=post&formato=2&cantidad=1&expresion=mfn=000790 [ Links ]
Robinson, D. S. 1991. Peroxidases and their significance in fruits and vegetables. In: Fox P. F. (ed). Food Enzimology. Vol. 1. Elsevier, London. pp: 399-426. [ Links ]
Rodríguez-Pedroso A. T., Ramírez-Arrebato M. A., Cárdenas-Traviéso R. M., Falcón-Rodríguez A., Bautista-Baños S. 2006. Efecto de la quitosana en la inducción de la actividad de enzimas relacionadas con la defensa y protección de plántulas de arroz (Oryza Sativa L.) contra Pyricularia grisea sacc. Revista Mexicana de Fitopatología, 24, 1; 1-7. https://www.researchgate.net/profile/Silvia_Bautista-Banos/publication/237037513_Efecto_de_la_Quitosana_en_la_Induccion_de_la_Actividad_de_Enzimas_Relacionadas_con_la_Defensa_y_Proteccion_de_Plantulas_de_Arroz_Oryza_sativa_L_contra_Pyricularia_grisea_Sacc/links/0deec51b2975c15955000000.pdf [ Links ]
Rüttimann, C., E. Schwember, L. Salas, D. Cullen, and R. Vicuna. 1992. Ligninolytic enzymes of the white rot basidiomycetes Phlebia breviospora and Ceriporiopsis subvermispora. Biotechnology and Applied Biochemistry 16:64-76. [ Links ]
Senasica-Sagarpa. 2010. Protocolo de diagnóstico de Candidatus Liberibacter spp mediante la Técnica Reacción en Cadena de la Polimerasa (PCR) en Tiempo Real. Dirección General de Sanidad Vegetal. [ Links ]
SIGMA-ALDRICH. 2015 http://www.sigmaaldrich.com/technicaldocuments/protocols/biology/enzymaticassayofaamylase.printerview.html [ Links ]
Singleton, V. L., Orthofer, R., Lamuela-Raventos R. M. 1999. Analysis of total phenols and other oxidation substrates and antioxidants by means of folin-ciocalteau reagent. Methods in Enzymology 299: 152-178 http://dx.doi.org/10.1016/S0076-6879(99)99017-1 [ Links ]
Trotel-Aziz. P., Couderchet M., Biagianti S. and Aziz A. 2008. Characterization of new bacterial biocontrol agents Acinetobacter, Bacillus, Pantoea and Pseudomonas spp. mediating grapevine resistance against Botrytis cinérea. Enviromental and Experimental Botany. 64: 21-32. http://dx.doi.org/10.1016/j.envexpbot.2007.12.009 [ Links ]
Wang C.Y. 1995. Effect of temperature preconditioning on catalase, peroxidase, and superoxide dismutase in chilling zucchini squash. Postharvest Biology and Technology 1: 33-45. doi:10.1016/0925-5214(94)00020-S [ Links ]
Zhong Y., Cheng C. Z., Jiang N. H., Jiang B., Zhang Y. Y., Wu B., Hu M. L., Zeng J. W., Yan H. X., Yi G. J., Zhong G. Y. 2015. Comparative transcriptome and iTraq proteome analyses of citrus root responses to Candidatus Liberibacter asiaticus infection. PLoS ONE. 10(6):e0126973. http://dx.doi.org/10.1371/journal.pone.0126973 [ Links ]
Received: September 04, 2016; Accepted: January 13, 2017
 Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
