











 nueva página del texto (beta)
nueva página del texto (beta) Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
The phyllosphere refers to the habitat encompassing the external surface and internal tissues of plant leaves, and it is the most extensive terrestrial habitat, covering nearly twice the Earth’s surface (Vorholt 2012). Bacteria are the most abundant microorganisms in the phyllosphere, occupying approximately 106 to 107 cells per cm2 of leaf tissue (Vorholt 2012). The bacteria living in the phyllosphere are epiphytic when they live on the surface and endophytic when they inhabit the internal tissues of the leaves (Vorholt 2012, Dastogeer et al. 2020). Both epiphytic and endophytic bacteria play an essential role in plant growth, development, and reproduction (Yadav et al. 2021). The endophytic microbiota of the phyllosphere consists of different taxa which are considered core microbiota when consistently present across multiple samples of the species, and other taxa that are variable and transient (Morella et al. 2020, Risely 2020). The contribution of core endophytic microbiota to nutrient acquisition and plant growth is assumed, as they can act as symbionts of the species. On the other hand, variable and transient (non-core) microbiota may be associated with differences in the host’s habitat. For instance, in Suaeda salsa (L.) Pall., the microbiota diversity in young leaves increases with phosphorus content in soil, while diversity decreases in mature leaves as soil pH increases (Zhou et al. 2023).
Within the microbiota, we can find nitrogen-fixing bacteria known as diazotrophs or diazotrophic bacteria, which have the metabolic capacity to reduce dinitrogen (N2) to ammonia (NH3) through the nitrogenase enzyme (Fe-Pérez et al. 2015). There are three nitrogenase isoforms, the most common being Mo-nitrogenase, which is encoded by a complex of genes including nifH, which is responsible for mediating ATP-dependent electron transfer during catalysis (Li et al. 2014, Kuypers et al. 2018). The nifH gene is widely distributed in diazotrophic bacteria (Sun et al. 2021). Therefore, it is the most used biomarker to study phylogeny, diversity, and potential diazotrophic activity (Gaby & Buckley 2014).
Symbiotic associations between diazotrophic bacteria and plants have traditionally been found in legume root nodules (Carvalho et al. 2014, Hernández-Rodríguez et al. 2017), where they fix nitrogen and transfer it to the host (Turner et al. 2013, Shin et al. 2016, Rosado et al. 2018). Diazotroph symbiotic associations are not limited to root nodules; in the last two decades, diazotrophic bacteria have been found in the phyllosphere playing an important role (Fürnkranz et al. 2008, Abadi et al. 2021). Estimates of nitrogen fixation rates in the Amazon suggest that nitrogen fixation in the phyllosphere is ten times higher than in the soil, with rates of 0.33 and 0.03 kg N ha⁻¹ yr⁻¹, respectively (Moreira et al. 2021). Nitrogen-fixing bacterial genera such as Bacillus, Pseudomonas, Pantoea, Microbacterium, Sphingomonas, Methylobacterium, Arthrobacter, and Azospirillum are common in the phyllosphere (Shabanamol et al. 2018, Bao et al. 2019, Liang et al. 2019, Abadi et al. 2021). Studies on diazotrophic communities have been conducted on annual crops such as Saccharum officinarum L. (Singh et al. 2022) and Zea mays L. (Zhang et al. 2022); in fruit trees such as Pyrus pyrifolia (Burm.f.) Nakai, Prunus armeniaca L., Prunus avium (L.) L., and Vitis vinifera L. (Liang et al. 2019), and shrubs such as Jatropha curcas L. (Madhaiyan et al. 2015). Diazotrophs have also been reported in temperate timber species as epiphytic microbiota of the phyllosphere in Quercus ilex L. (Rico et al. 2014), Pinus bungeana Zucc. ex Endl., and Sabina chinensis L. (Bao et al. 2020). Other reports of diazotrophs of the phyllosphere of timber temperate species, found them as endophytic microbiota in Populus trichocarpa Torr. & A.Gray ex Hook. (Doty et al. 2016), Pinus contorta Douglas ex Loudon (Padda et al. 2019), and Thuja plicata Donn ex D.Don (Bal et al. 2012). To our knowledge, no studies have examined the potential diazotroph taxa inhabiting the phyllosphere of tropical American timber species.
Cordia dodecandra DC. is a tree recognized in the Yucatan Peninsula (Mexico) for its use both as timber and a fruit tree (Pennington & Sarukhán 2005). The species in managed in smallholder properties as monoculture plantations and silvopastoral systems (Campos et al. 2015, López-Ramírez et al. 2023). The endophytic microbiota in the phyllosphere of C. dodecandra includes Methylobacterium, Actinomycetospora, Aureimonas, Allorizobium and Sphingomonas as dominant genera in forest and homegarden populations across two different regions in Yucatan (May-Mutul et al. 2022). Hence, they may be considered part of its core microbiota. Some members of Methylobacterium, Allorizobium and Sphingomonas are considered to have the metabolic capacity to fix nitrogen (Madhaiyan et al. 2015, Stone et al. 2018, Camargo-Neves & Araújo 2019, Zhu et al. 2022). In this study, we characterized the bacterial endophytic microbiota and assessed its potential diazotrophic communities in 21-year-old C. dodecandra growing in smallholder agroforestry systems. Exploratory comparisons of the diversity of endophytic microbiota in monoculture plantations and silvopastoral systems were also conducted, as carbon, nitrogen, and phosphorus levels are approximately two, three, and one hundred times higher, respectively, in the soil of silvopastoral systems (López-Ramírez et al. 2023).
Materials and methods
Study species and sites. Cordia dodecandra, locally named as ciricote, is a deciduous tree native to southern Mexico and Central America (Miranda & Hernández-X 1963), widely used for timber (Pennington & Sarukhán 2005). Due to deforestation and the decline of wild populations, the National Forestry Commission of Mexico has identified the species as a priority for conservation and management (López-Upton et al. 2011). The species is an important component of Mayan homegardens (Hurtado-Torres et al. 2020) and agroforestry systems at the region (Campos et al. 2015).
This study was conducted at Rancho Juanes (Figure 1), where C. dodecandra plantations were established in 2002. The trees, sown simultaneously at three months of age and originating from the same nursery, were therefore 21 years old at the time of the study. The monoculture plantations are characterized by the dominance of C. dodecandra in the tree strata and some cacti growing under the trees, while in the silvopastoral system Cynodon plectostachyus (K.Schum.) Pilg. grass was sown for sheep fodder. The C. dodecandra trees present foliage from June to November in the monoculture plantations, while in the silvopastoral system the foliage presence is extended through all year excepting October (Campos et al. 2016). Pruning of C. dodecandra trees is performed twice a year in both monoculture plantation and silvopastoral system, while at the latter sprinkler irrigation, weed control and incorporation of sheep excreta are additional management practices (Campos et al. 2015).
Figure 1 Location of the study site in Rancho Juanes at Xmatkuil locality in the Yucatan state of Mexico.
According to NOM-021 (SEMARNAT 2000), the soils of the silvopastoral system and the monoculture plantation are quite similar, with high levels of nitrogen (> 0.25 %), calcium (< 10 cmol kg⁻¹), and organic matter (> 6.0 %). Nonetheless, the soil in the silvopastoral system shows higher concentrations of organic carbon, organic matter, carbonates, nitrogen, inorganic phosphorus, and calcium compared to that of the monoculture plantation. These differences are attributed to the deposition of sheep feces and the biomass left behind after grazing (López-Ramírez et al. 2023).
Sampling design. In July 2023, before the rainy season began, two composite samples were obtained for each tree: one from 10 young leaves and another from 10 mature leaves, both collected from the middle canopy, adding up to 12 initial samples. Leaves were sterilized with 70 % ethanol and transported on ice to the laboratory, where they were stored at -70 °C. To optimize the representation of healthy microbiota, the inclusion criteria for C. dodecandra trees were: 1) presence of young and mature leaves, 2) no evidence of herbivory on the leaves, and 3) no apparent fungal, viral or bacterial damage on the leaves. Only two individuals in the monoculture plantation and four in the silvopastoral system met these criteria.
Dasometric and foliar nutrient characterization of trees in the agroforestry systems. The variables height, DBH (diameter at breast height), and canopy cover, as described by Campos et al. (2015), were obtained as indirect measurements of tree growth (Table 1). Two weeks after initial collection nutrient analyses were performed. To estimate total nitrogen and carbon content in the foliar tissue of each tree, the Dumas method was conducted in 250 mg ground tissue with a 70-mesh-size, previously dried to constant weight at 70 °C (Simonne et al. 1994). The estimates were obtained by duplicates in the LECO CN 828 analyzer (LECO, St. Joseph, Michigan). The Nessler reactive method (Madero et al. 1998) was used to determine the foliar ammonium content with 1 g of tissue by triplicates. Due to equipment problems, nutrient content was not properly estimated on the 12 initial samples and a new composite sample of 20 leaves (young and mature) per tree were collected to analyze carbon, nitrogen (total and ammonium) as previously described.
Table 1 Dasometric measurements and foliar nutrient content of Cordia dodecandra trees growing in Rancho Juanes Yucatan at monoculture plantation and silvopastoral system
| Variable | Rancho Juanes | Monoculture plantation | Silvopastoral |
|---|---|---|---|
| Cover canopy (m) | 22.92 ± 9.93 | 20.90 ± 2.8 | 23.92 ± 10.78 |
| Height (m) | 5.91 ± 1.31 | 4.92 ± 0.2 | 6.40 ± 1.17 |
| DBH (cm) | 43.33 ± 22.15 | 24.15 ± 9.05 | 15.02 ± 1.36 |
| Foliar nitrogen (%) | 1.96 ± 0.21 | 1.72 ± 0.15 | 2.09 ± 0.06 |
| Foliar carbon (%) | 40.75 ± 1.27 | 41.05 ± 1.72 | 40.60 ± 0.81 |
| Foliar NH4+ (mg/Kg) | 664.70 ± 187.47 | 730.26 ± 182.43 | 631.92 ± 172.98 |
Endophytic bacterial communities in agroforestry systems. After five days of collection, leaves were sterilized with 96 % ethanol and 2 % commercial sodium hypochlorite and then rinsed five times with sterile distilled water (Patel & Archana 2017). Small sections (1 cm2) were cut from each leaf from the 12 initial samples and macerated with liquid nitrogen. DNA was isolated from 125 mg of the foliar tissue with ZymoBIOMICS™ DNA Miniprep Kit (Zymo Research, Irvine, California). The concentration and purity of DNA samples were evaluated using a Multiskan™ GO microplate spectrophotometer µDrop plate with SkanIt v. 7 software (Thermo Scientific™, Whaltham, Massachusetts). We verified the DNA integrity by TBE buffer-electrophoresis in 1.2 % agarose gels that were stained with ethidium bromide and visualized with UV light. The 12 initial samples and a negative control (DNAse-RNAse free water) were then amplified for the 16S rRNA gene using primers 16S-Fw (5'-GTAGACGCTATTCCCTAAGAT-3') and 16S-Rv (5'-CTCCCAACTATACAGTACACACCTCATA- 3’) (Stevanovic et al. 2011). A 25 μL reaction with 100 ng DNA, 12.5 μL GoTaq Green Master Mix 2X (Promega, Madison Wisconsin), and 10 μL of DNAse-RNAse free water was amplified in the C1000 Touch thermal cycler PCR system (Bio-Rad, Hercules, California). The PCR program consisted of an initial denaturation at 94 °C for 5 min followed by 34 cycles (94 °C for 30 s short denaturation, 58 °C primer annealing for 30 s, and 72 °C extension for 90 s) with a final extension at 72 °C for 5 min. Amplification was evaluated by electrophoresis as previously described. The library construction and DNA sequencing were performed in Novogene (Novogene, Shangai, China) on the Illumina NovaSeq 6000 platform using 515F (5-GTGYCAGCMGCCGCGGGTAA- 3’) and 907R (5'-CCGTCAATTCMTTRAGTTT- 3’) bacterial universal primers for the V4 toV5 region of 16S rRNA (Pichler et al. 2018, Sebastião et al. 2015, Zhou et al. 2024).
The quality of raw sequences from the 12 initial samples was checked using FastQC (Andrews 2010). The pipeline reported by May-Mutul et al. (2022), was used in QIIME2 v 2023.2 (Bolyen et al. 2019), for demux, denoise, and assign the taxonomic group to ASVs. The latter was performed using the 138-99 weighted database of Silva (Kaehler et al. 2019). For further analyses, ASVs corresponding to host chloroplasts and mitochondria were filtered. An evaluation of the abundance of sequences for the 12 initial samples showed a large variation in sample size (65 to 5,490 sequences per sample). To increase the number of sequences per sample and reduce sample size variation, the ASVs from young and mature leaves were combined for each tree, resulting in six final merged samples. The final diversity analyses were performed in R v4.3.2 (R Core Team 2023), using phyloseq package v1.50.0 (McMurdie & Holmes 2013) as outlined in the pipeline (Supplemetary material S1). Due to the small number of trees grouped in the monoculture plantation (2 trees) and in the silvopastoral system (4 trees) the comparative analyses must be considered exploratory. The number of observed ASVs, Chao1, and Shannon indexes were estimated to characterize the alpha diversity. Means of the Shannon index from monoculture plantation and silvopastoral system were compared using Wilcoxon rank test on rarefied samples to even depths. To correct for differences on the observed sequences variation per sample (496 to 6,814), normalization of the six final merged samples was attained using cumulative sum scaling transformation (CSS). Principal Coordinate Analysis, ANOSIM, and PermANOVA (Zhang et al. 2022), were performed on the Bray-Curtis distances using both raw and normalized CSS data to test for differences on the composition and structure of endophytic communities. Relative abundance by order and genus was plotted for majoritarian taxa, defined as those representing more than 1 % of the total observed sequences. A differential abundance analysis ANCOMBC (Lin & Peddada. 2020) was performed on the relative abundance for the ASVs grouped by genus using both raw and normalized CSS data. In the result section only plots and heatmap from normalized CSS data were included.
Potential diazotroph-endophytic communities. The presence of potential diazotrophic bacteria was evaluated by endpoint PCR using the primers PolF (5'-TGCGAYCCSAARGCBGACTC 3’) and PolR (5'-ATSGCCAT-CATYTCRCCGGA) to amplify the region 115 (134) to 457 (476) of the nifH gene (Gaby & Buckley 2017). We used the DNA from root nodules of Leucaena leucocephala growing in Lerma, Campeche, as a positive control for amplification, as the presence of diazotrophs in its rhizosphere has been reported (Gibson et al. 1982) and the DNA from the 12 initial samples. The PCR reaction was similar to that described previously for 16S rRNA, although primers were at 10 pmol, and the program consisted of an initial denaturation at 95 °C for 3 min followed by 30 cycles (95 °C for 1 min short denaturation, 55 °C primer annealing for 1 min, and 72 °C extension for 1 min) with a final extension step at 72 °C for 7 min (Rusmana & Lestari 2015). Amplification was evaluated by electrophoresis as described before. The DNA of the 12 initial samples was sent to Novogene (Shangai, China) for library construction and sequencing of the nifH gene, using Pol-Fw and Pol-Rv primers, on the Illumina NovaSeq 6,000 platform. Successful amplification, library construction and sequencing in Novogene were only obtained in 3 out of the 12 initial samples. The quality of raw sequences from these 3 samples was checked using FastQC (Andrews 2010), and the pipeline in the supplementary material (Supplemetary material S2) was used in QIIME2 v. 2023.2 (Bolyen et al. 2019), for demux and denoise with dada2. The taxonomic assignment with nifH genes was obtained using the Marine Microbial Ecology database of University of California, Santa Cruz (Heller et al. 2014). Unassigned ASVs were filtered and excluded, and only those identified at the species level are reported here as potential diazotrophs, whit its relative abundance per sample (FigureS1).
To evaluate the relative abundance of the nifH gene, the initial 12 samples and the positive control of DNA from root nodules of L. leucocephala were amplified by qPCR. The 20 μL reactions consisted of 200 nM of each primer (PolFw and PolRv), 5 μL SSoAdvanced Universal SYBR® Green Supermix (Bio-Rad, Hercules, California), 50 ng of DNA, and DNAse- & RNAse-free water. The qPCR program consisted of an initial denaturation at 98 °C for 30 s followed by 40 cycles of short denaturation at 95 °C for 15 s an annealing at 62 °C for 30 s and extension at 72 °C for 40 s (Zhang et al. 2022) in the CFX96™ Touch Real-Time PCR Detection System (Bio-Rad, Hercules, California). The specificity of amplification at different temperatures was evaluated by melting curves and the relative abundance of the nifH gene was estimated by the fold change of amplification relative to the positive control. The threshold amplification for nifH gene was at cycle 14 for positive control and between cycles 23 and 25 for the 12 initial C. dodecandra samples. The curves started their stationary phase at cycle 32 (Table S1).
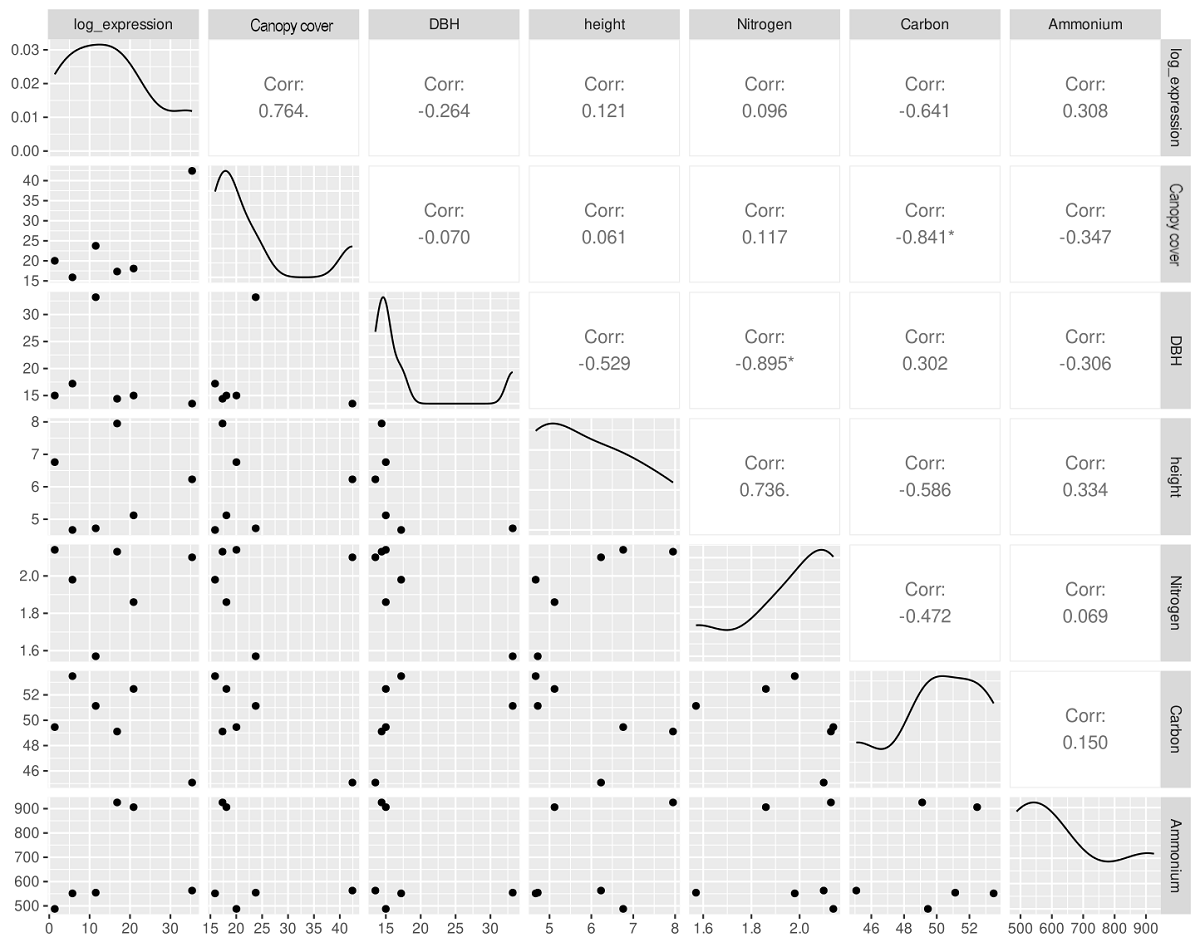
To test for an association between the relative abundance or the nifH gene and host tree growth, a series of Pearson correlation analysis (Holm-corrected P) were performed using dasometric measurements -DBH, height, and canopy cover, and the nutrient contents: carbon, nitrogen and ammonium (Table 1) against fold change values. The fold change values per tree was obtained by adding the values obtained for the young leaves and the mature leaves of the 12 initial samples for each of the six trees (Table S1).
Results
Dasometric and foliar nutrient characterization of trees in the agroforestry systems. The 21-year-old trees assessed in this study had on average a height of 5.91 m ± 1.31 m, DBH of 18.07 cm ± 7.51 cm, and a canopy cover of 22.92 m2 ± 9.93 m2. A large variation in the dasometric variables was observed among the six trees, with coefficients of variation of 22 for height, and 42 % and 43 % for DBH and canopy cover, respectively (Table 1). Variation among trees exceeded 50 % in DBH for trees from monoculture plantations and in canopy cover for trees from the silvopastoral system. Nutrient content in the foliar tissue was less variable among trees with an average of 1.96 ± 0.21 % and 40.75 ± 1.27 % for nitrogen and carbon, respectively, although variation in the ammonium was larger with an average of 664.70 ± 187.47 g/kg resulting in a coefficient of variation of 28.20 % for all trees, and 27.37 and 28.59 % in monoculture plantation and silvopastoral system, respectively (Table 1).
Endophytic bacterial communities in agroforestry systems. We obtained 2,474,697 paired reads of 250 pb in length that were down to 15,632 sequences after filtering host’s chloroplasts and mitochondria. The number of sequences ranged from 496 to 6,814 per final merged samples (six trees). The rarefaction of the sequences to an even depth of 496 per sample, were plotted in curves that began to flatten at depths of 250 for Shannon index, but did not for the observed ASVs, therefore we did not perform comparisons for observed ASVs and Chao index (Figure 2A). The endophytic microbiota of C. dodecandra phyllosphere included 420 ASVs. The comparison of Shannon diversity index values between monoculture plantation and silvopastoral system was no significant (P = 1) and ranged from 2.84 to 3.72 (Figure 2B). However taxonomic assignment was higher than 90 % at the family level and less than 65 % for genus and species taxa. The majoritarian orders (with a relative abundance higher than 1 % and present in all samples) were i) Rhizobiales and Sphingomonadales of the class Alphaproteobacteria; and ii) Kineosporiales, Frankiales, Micrococcales, and Pseudonocardiales of the class Actinobacteria (Figure 3A). The majoritarian endophytic bacteria included 18 classified at the genus level, six grouped at the family level, and two grouped at class level (Figure 3B). Only Methylobacterium-Methylorubrum, Aureimonas, Sphingomonas, Microbacterium, Quadrisphaera, Pseudokyneococcus, Curtobacterium, and Actinomycetospora were present in all samples (Figure 3B). The endophytic communities of the phyllosphere from silvopastoral system were grouped closer according to the principal coordinate analysis (Figure 4). However, no significant differences were observed in the Bray-Curtis distances between endophytic communities of monoculture plantation and silvopastoral according to ANOSIM-permANOVA (P adj = 0.4). No differences in the abundance at the genus level were observed either between agroforestry systems, and the heatmap plotting the results from ANCOMBC suggests that each tree has different genera of endophytic bacteria enriched in their phyllosphere (Figure 5).
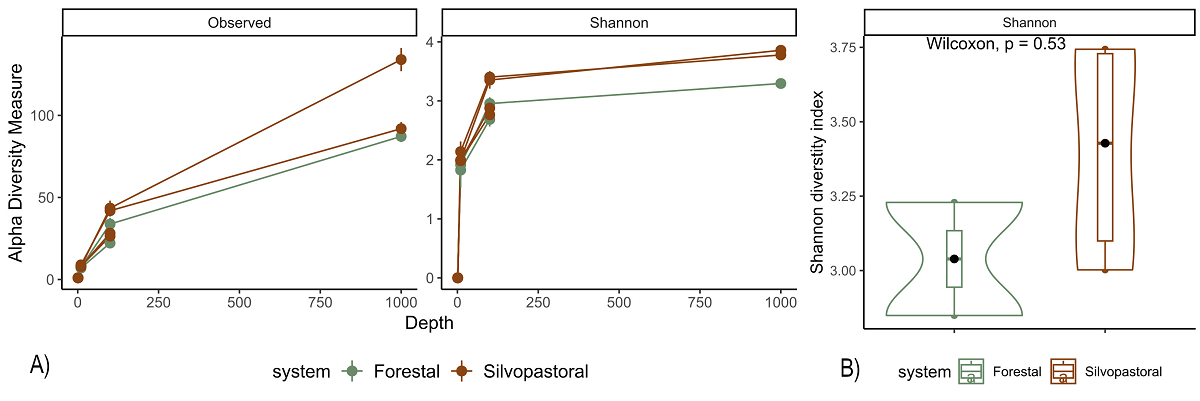
Figure 2 Rarefaction curves of alpha diversity indices for the endophytic microbiota in the phyllosphere of Cordia dodecandra, based on observed ASVs and the Shannon diversity index. A Wilcoxon test was used to compare Shannon diversity between trees growing in monoculture plantations (green) and silvopastoral systems (brown) in Yucatán
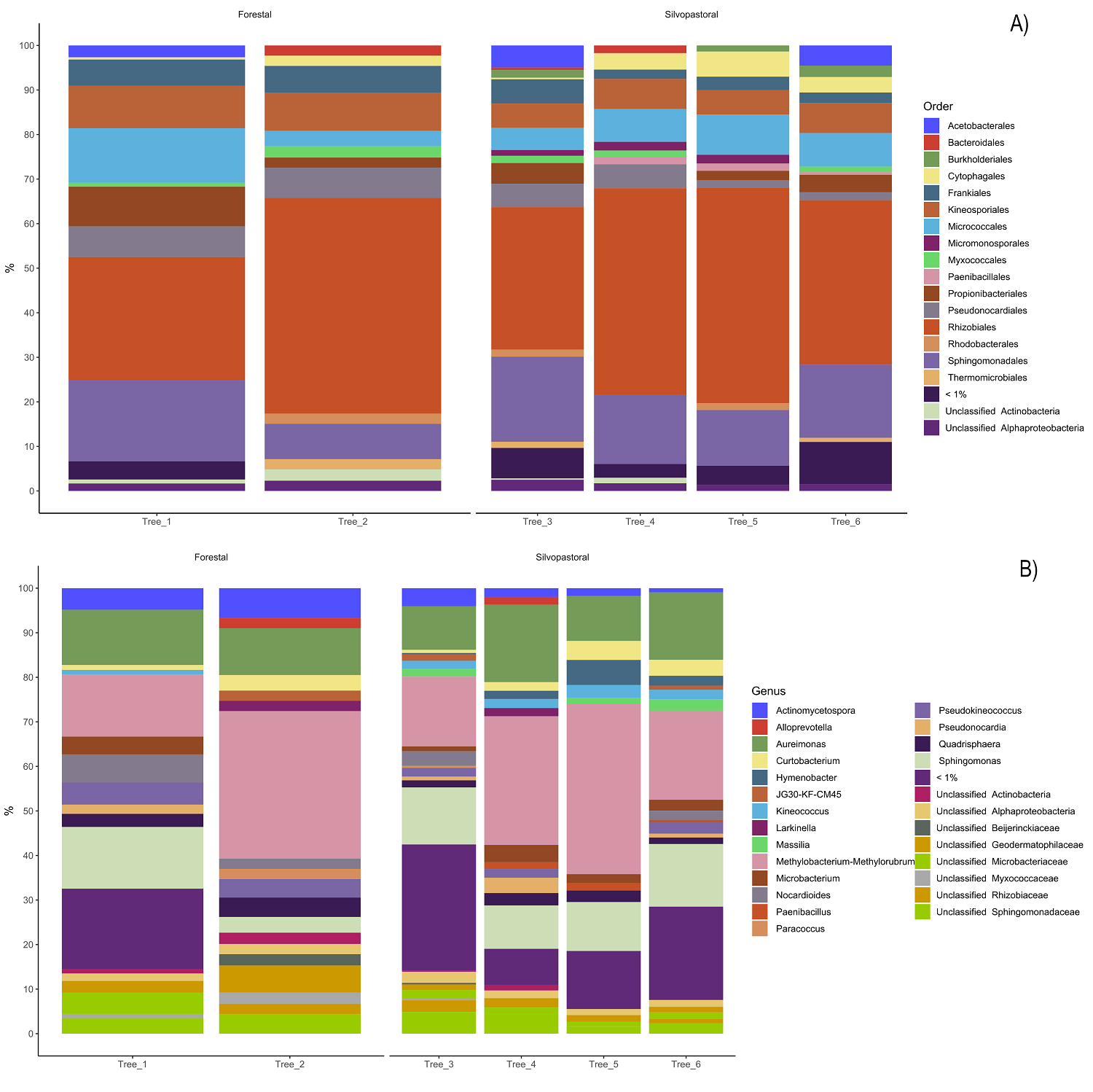
Figure 3 Relative abundance of the majoritarian (relative abundance < 1 %) taxa classified at the A) order and B) genus level of the 420 ASVs of endophytic microbiota in the phyllosphere of Cordia dodecandra trees growing in monoculture plantation and silvopastoral systems in Yucatan.
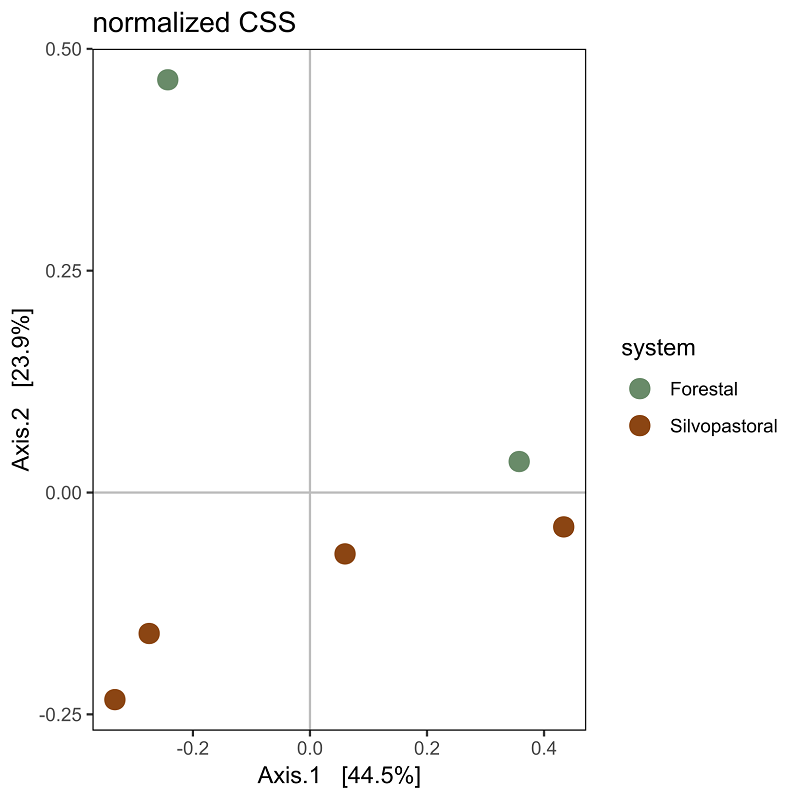
Figure 4 Principal coordinate analysis (PCoA) of the endophytic microbiota in the phyllosphere of Cordia dodecandra trees growing in monoculture plantation (green) and silvopastoral systems (brown) in Yucatan.
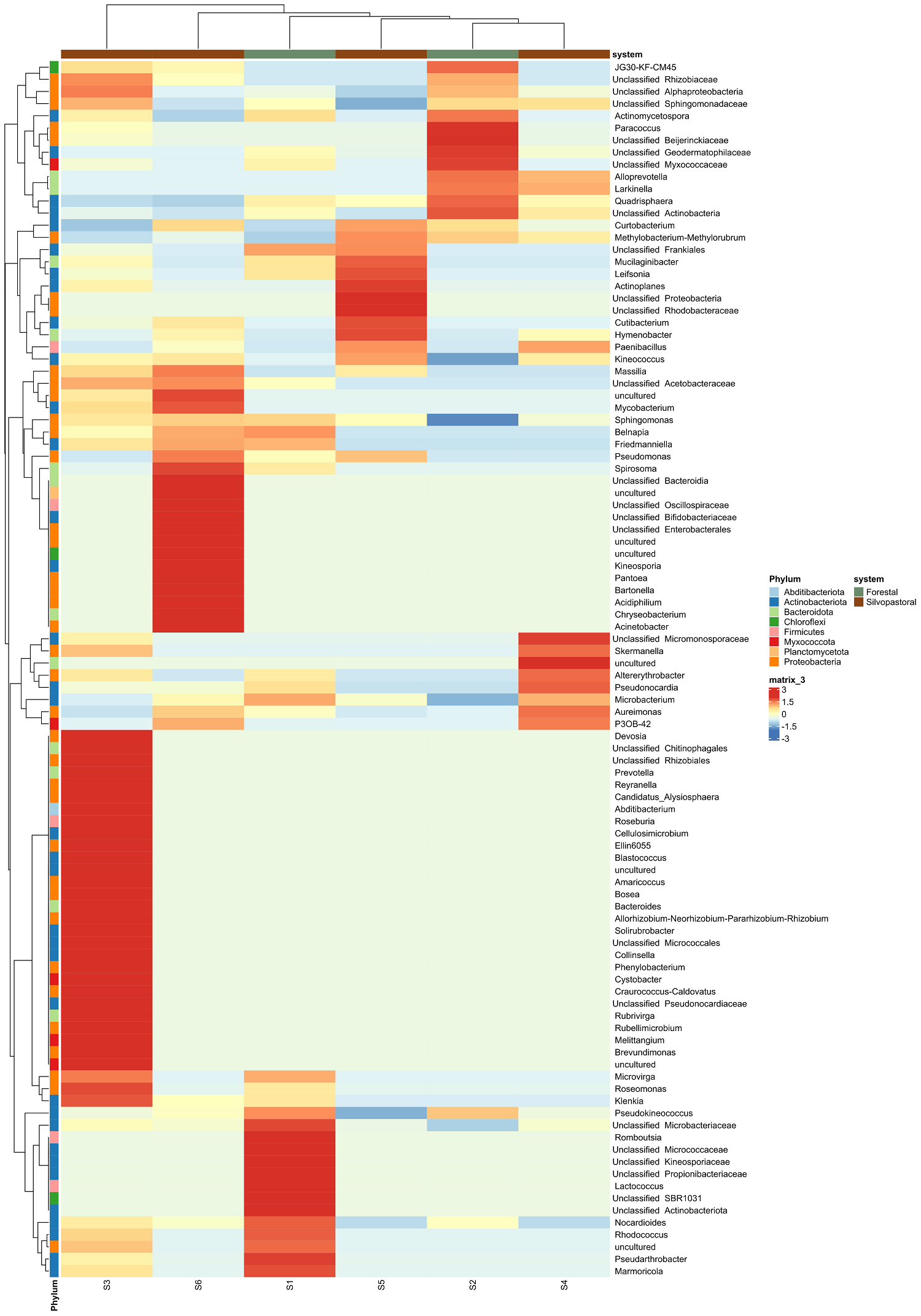
Figure 5 Heatmap showing the relative abundance of endophyte bacteria of the C. dodecandra phyllosphere at the genus level based on differential abundance analysis. No significant differences among the relative abundance of the genera were found between monoculture plantations and silvopastoral systems, according to ANCOM-BC results (see text for details).
Potential diazotroph-endophytic communities. A total of 495,747 paired 360 bp reads of the nifH gene were obtained for the three samples that were successfully amplified and sequenced. After exclusion of the unclassified ASVs, 26 remained, and sample size of the three samples ranged from 1,898 to 10,307 sequences. The ASVs were assigned to six potential diazotrophs species (Table 2). The most abundant order among potential diazotrophs was Nostocales with abundances ranging from 60 to 90 %. At the species level, Hassalia byssoidea and Nostoc commune were the most abundant potential diazotrophs with relative abundances of 60 and 10 %, respectively (Figure S1).
Table 2 Classification for the diazotroph-endophytic microbiota in the phyllosphere of Cordia dodecandra in Yucatan using nifH database from The Marine Microbial Ecology database and ID and authors of the Genbank accession.
| Species | Taxonomic clasification | Genbank ID | Authors |
|---|---|---|---|
| Alcaligenes faecalis | D_Bacteria. p_Pseudomonadota. c_Betaproteobacteria. o_ Burkholderiales. f_Alcaligeniaceae | CCF77362.1 | Shahid 2013 |
| Ensifer americanum | D_Bacteria. p_ Proteobacteria. c_Alphaproteobacteria. o_ Rhizobiales. f_Rhizobiaceae | AIX97566.1 | Cordero et al. 2016 |
| Methanosaeta concilii | D_Archaea p_ Stenosarchaea. c_ Methanomicrobia. o_ Methanotrichales. f_Methanotrichaceae | AEB67348.1 | Barber et al. 2011 |
| Chroococcidiopsis thermalis | D_Archaea p_Cyanobacteriota. c_ Cyanophyceae. o_ Chroococcidiopsidales. f_ Chroococcidiopsidaceae | AAQ99146.1 | Boison et al. 2004 |
| Hassallia byssoidea | D_Archaea p_Cyanobacteriota. c_ Cyanophyceae. o_ Nostocales.f_Tolypothrichaceae | KIF38087.1 | Singh et al. 2015 |
| Nostoc commune | D_Archaea p_Cyanobacteriota. c_ Cyanophyceae. o_ Nostocales. f_ Nostocaceae | ABF81842.1 | Yeager et al. 2007 |
All 12 initial samples from young and mature leaves for each one of the trees were successfully amplified using qPCR. Values of the fold change were similar between monoculture plantation and silvopastoral system (t = 0.15, P = 0.88). After adding nifH gene fold change values for each tree, the correlations with values obtained to characterize each tree was determined, and only canopy cover had a marginally significant and positive correlation (r = 0.76, P = 0.06) with the nifH relative abundance. No significant correlations were observed with DBH and height (r = 0.26, P = 1 and r = 0.12, P = 1, respectively) or with carbon, total nitrogen and ammonium (r = -0.64, P = 0.17, r = 0.10, P = 0.89 and r = 0.31, P = 0.52, respectively) (Figure 6).
Discussion
Dasometric and foliar nutrient characterization of trees in the agroforestry systems. In this study, C. dodecandra displayed high variability in dasometric measurements and foliar ammonium content, but relatively low variability in foliar carbon and nitrogen content. Although the trees examined originated from the same nursery and were established simultaneously, the recorded variability may have genetic and environmental biases. Campos et al. (2015) report that the trees from Rancho Juanes agroforestry systems came from four different nurseries. They also found that differences in management, mainly irrigation, weeding and pruning are associated with variations in the DBH, height and canopy cover. They observed significant differences in these variables between the monoculture plantations and the silvopastoral system; however, our study failed to find this effect, largely because of the small sample size of trees included. The foliar nitrogen and carbon measured in our study are intermediate compared to those reported for trees in homegardens and forest populations (Estrada-Medina et al. 2023), suggesting that the species has phenotypic plasticity to reallocate these nutrients to its foliage from the soil or other sources. The ammonium (NH4+) found on the C. dodecandra leaves was variable with three out of the six trees having almost two times more than the others (Table 1). This variability may reflect differences in the potential absorption of atmospheric N2, nitrogen absorption and fixation or protein metabolism (Shin et al. 2016). Further studies to determine the nitrogen source of ammonium within the foliage are needed, particularly as potential diazotroph endophytes were found in this study as we will discuss in the next sections.
Endophytic bacterial communities in agroforestry systems. The endophytic communities of the phyllosphere of C. dodecandra are relatively rich with up to 420 ASVs and Shannon diversity indexes around 3. The endophytic communities did not differ between the agroforestry systems. The small sample size (six trees) and the large variation in the number of 16 S rRNA reads obtained per sample diminished the power of the test implemented in our study. The composition of endophytic communities varied among trees, with different genera enriched in each tree sample (Figure 5). Variation in the endophytic communities of the phyllosphere may be associated with the position of leaves within the canopy and the height relative to the base of the tree (Laforest-Lapointe et al. 2016, Herrmann et al. 2021). Additionally, specific endophyte species may help individual trees cope with distinct microenvironmental stresses. Soil physicochemical characteristics, such as phosphorus availability, redox potential, and sand content, also varied considerably in both the monoculture plantation and the silvopastoral system (López-Ramírez et al. 2023), with potential impacts on tree growth and development. Determining how canopy and soil conditions associated with each tree affect the diversity of endophytic microbiota in the phyllosphere of C. dodecandra requires further experimental investigation.
Some of the most abundant bacterial orders, such as Rhizobiales, Sphingomonadales, and Pseudonocardiales, have previously been described in the rhizosphere of C. dodecandra (López-Ramírez et al. 2023), as well as among phyllosphere endophytes in different populations (May-Mutul et al. 2022). Among the eight genera present in all samples with relatively large abundance (< 1 %), six were also recorded as the majoritarian genera in homegarden and forest populations from two different regions in Yucatan (May-Mutul et al. 2022). We propose that Methylobacterium-Methylorubrum, Aureimonas, Sphingomonas, Quadrisphaera, Curtobacterium, and Actinomycetospora may constitute the core endophytic microbiome of C. dodecandra phyllosphere. In addition to those genera, Microbacterium, Quadrisphaera, and Pseudokineococcus may play important roles in the agroforestry systems of Rancho Juanes. Several of these genera have been identified as endophytes in the phyllosphere of other plant species: Quadrisphaera in rice (Oryza sativa L.) (Muangham et al. 2019) and peanut (Arachis hypogaea L.) (Li et al. 2023); Aureimonas in Jatropha curcas (Madhaiyan et al. 2013) and Colobanthus quitensis (Kunth) Bartl. (Perazzolli et al. 2022), and Pseudokineococcus in Morus spp. (Chen et al. 2022). Methylobacterium and Sphingomonas have been reported in the phyllosphere of Saccharum spp. (Hamonts et al. 2018), Glycine max (L.) Merr., Trifolium repens L., and Arabidopsis thaliana (L.) Heynh. (Thapa & Prasanna 2018), while Curtobacterium has been found in Zea mays and Glycine max (Dunleavy 1989). The widespread presence of these genera across various angiosperms suggests that a core group of endophytic microbiota may be adapted to the phyllosphere, potentially establishing long-term symbiotic relationships with their hosts. We propose that Actinomycetospora, Methylobacterium, Sphingomonas, and Curtobacterium may contribute to plant growth in both monoculture plantations and silvopastoral systems. For example, Actinomycetospora has been shown to increase the dry weight of Lemna aequinoctialis Welw. by 1.3 to 1.4 times (Saimee et al. 2024). Additionally, Methylobacterium, Sphingomonas, and Curtobacterium have been reported to promote plant growth and suppress pathogens (Chase et al. 2016, Garrido et al. 2016). Therefore, it is plausible that these dominant genera play a key role in the growth and health of C. dodecandra, and their functions merit further investigation.
Potential diazotroph-endophytic communities. We identified six species that have the diazotroph potential inhabiting as endophytes of C. dodecandra phyllosphere in only three out of twelve samples of the agroforestry systems. The species did not belong to the majoritarian genera of the endophytic microbiota, and were not successfully amplified and sequenced from all trees. This suggests that the potential diazotrophic endophtyes may be rare species; however, the qPCR successful amplification allows us to establish that all individual trees have endophytic microbiota with a potential diazotroph function. The six potential diazotrophs identified in C. dodecandra phyllosphere by their sequence of the nifH gene, have been reported as microbiota of the phyllosphere, except for Alcaligenes faecalis. Alcaligenes faecalis, originally isolated from Oriza sativa with strain A15 was considered a diazotroph (You & Zhou 1989). Alcaligenes faecalis has been found as a diazotroph in rhizosphere soils on Sesbania aculeata Poir. and Atriplex lentiformis (Torr.) S.Watson (Babar et al. 2021) with the ability to consume nitrite and nitrate (Chen et al. 2021) and on the leaves of Ixora chinensis Lam., Ervatamia divaricata (L.) Burkill, Hibiscus rosa-sinensis L., and Amaranthus cruentus L. (Undugoda et al. 2016). The second potential diazotroph identified was Ensifer americanum, which has been found as a diazotroph in roots of Phaseolus filiformis Benth. (Rocha et al. 2020), Anthyllis henoniana Coss. Ex Batt. (Yan et al. 2016), Glycine sp., Psoralea corylifolia L., Sesbania cannabina (Retz.) Pers., Medicago lupulina L. (Wang et al. 2013), and Acacia spp. (Lloret et al. 2007, Sakrouhi et al. 2016). The third potential diazotroph identified was Methanosaeta concilii (syn. Methanothrix concilii), a free-living bacterium (Auman et al. 2001) that is frequently found in soils (Bae et al. 2018). The fourth potential diazotroph identified was Chroococcidiopsis thermalis a facultative diazotroph (Boison et al. 2004, Lan et al. 2021) widely distributed in various xeric freshwater and marine habitats, forming cyanolichens for nitrogen fixation (Villanueva et al. 2018). The fifth potential diazotroph was Hassallia byssoidea, a diazotrophic heterocyst with antibiotic properties (Rossi et al. 2012), sometimes associated with streptophytic algae (Singh et al. 2015, Mukhin et al. 2018) such as Chaetosphaeridium globosum (Knack et al. 2015). The sixth potential diazotroph was Nostoc commune, a heterocystous cyanobacteria (Peng & Bruns 2019) that has been found in soil biocrusts (Lan et al. 2021) and in basidiocarps (Mukhin et al. 2018); also considered drought resistant (Hata et al. 2022). In addition to N2 fixation, Nostoc commune can establish surface consortia that immobilize and retain inorganic nitrogen when it is in excess (Mukhin et al. 2018).
The presence of the potential diazotroph microbiota in C. dodecandra phyllosphere may play other functional roles. It has been proposed that diazotroph communities may also influence leaf growth by promoting the synthesis of phytohormones (Dobbelaere et al. 2003) such as auxins, cytokinins, gibberellins, and N-acyl homoserine lactones, as experimentally observed in O. sativa (Defez et al. 2017). Tree and herb growth is enhanced when endophytic diazotrophs are present in Salix sitchensis Sanson ex Bong. and Populus trichocarpa (Knoth et al. 2013); in Saccharum officinarum (Antunes et al. 2022); in Trifolium repens and Lolium multiflorum Lam. (Hahn et al. 2014). In C. dodecandra a marginally significant positive correlation was found between canopy cover and nifH gene abundance, suggesting that endophytic diazotrophs in the phyllosphere may promote canopy growth in C. dodecandra, although it was not correlated with nitrogen and ammonium content in the foliage tissue. Previous studies suggest that there is a linear relationship between N2 fixation rates and relative nifH gene abundance (Reed et al. 2010); future studies on nitrogen fixation or nifH gene expression in C. dodecandra are needed to know whether nitrogenase is active in the phyllosphere.
The trees of C. dodecandra growing in agroforestry systems harbor a potential diazotrophic community that was variable among individual trees. The relatively large abundance and prevalence among samples of seven genera: Methylobacterium-Methylorubrum, Aureimonas, Actinomycetospora, Sphingomonas, Microbacterium, Quadrisphaera, and Pseudokineococcus place them as the potential core endophyte community of the phyllosphere of C. dodecandra. Six potential diazotroph species form part of the endophytic community of the phyllosphere of C. dodecandra, Alcaligenes faecalis, Ensifer americanum, Methanosaeta concilii, Chroococcidiopsis thermalis, Hassallia byssoidea, and Nostoc commune. Future experimental work is needed to deepen our understanding of the specific functions of these dominant taxa and to investigate nitrogen fixation by diazotrophic endophytic microbiota in the phyllosphere of this and other tropical timber species.
Supplementary material
Supplemental material for this article can be accessed here: https://doi.org/10.17129/botsci.3684

