











 nueva página del texto (beta)
nueva página del texto (beta) Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Grasslands cover about one-third of the ice-free land surface (Reynolds & Frame 2005), and despite their apparent simplicity, they support complex communities of plants and animals whose ecology is adapted to seasonal climatic fluctuations and intrinsic natural phenomena such as recurring fires (Axelrod 1985). Grasslands support livestock production in most of the world, but also, especially those in good condition, provide environmental services including biodiversity conservation, pollination, water retention, climate regulation, and recreation (Medina-Roldán et al. 2007, Bengtsson et al. 2019).
However, the ecological integrity of grasslands has been affected by human activities, with 46 % of their surface worldwide cleared to establish agroecosystems, while the remaining is used for the grazing by domestic animals (Peters et al. 2006, White et al. 2000). This grazing by domestic livestock has often exceeded the land`s carrying capacity (i.e., the grasslands have been “overgrazed”), resulting in severe consequences for biological conservation (Noss 1994, Reid et al. 2010). Such overgrazing has caused “…loss of biodiversity; lowering of population densities for a wide variety of taxa; disruption of ecosystem functions, including nutrient cycling and succession; change in community organization; and change in the physical characteristics of both terrestrial and aquatic habitats” (Fleischer 1994) and over 80 % of the original cover of North American grasslands has been modified (CEC 2015). Although widespread, overgrazing is a subtle form of habitat modification that develops in time, often exceeding the lifespan of the humans using the land, and therefore usually goes unnoticed (Mellink & Contreras 2014, Mellink & Riojas-López 2020). This lack of awareness, combined with socioeconomic pressures to continue ranching, often results in a failure to implement remedial management practices for grasslands conservation. In fact, grasslands are the biome with the smallest proportion of protected areas and consequently, face the highest conservation risk in the world (Hoekstra et al. 2005).
In Mexico, there are more than 1,000 species of native grasses (Poaceae), about 25 % of which are endemic to the country (Sánchez-Ken 2019). Mexican grasslands have been a center of diversification for the subfamily Chloridoideae, represented widely by the genera Eragrostis Wolf, Sporobolus R.Br., Chloris Sw, Muhlenbergia Schreb., and Bouteloua Lag. (Rzedowski 1975, 1991a,b, 2006, Peterson et al. 2007). Natural grasslands cover 9 % of the surface of the country (INEGI 2017), and two main types are recognized: those in semi-arid areas of the Mexican Plateau and those in cold regions above 4,000 m in elevation (Miranda & Hernández-X. 1963, Gómez-Pompa 1965, Rzedowski 1975, 2006).
The largest natural grasslands are found on the Mexican Plateau, ranging from northeastern Jalisco and neighboring areas of Guanajuato to northwestern Chihuahua and northeastern Sonora (Rzedowski 2006). These grasslands extend as a belt from central Mexico northwards across the Chihuahuan Desert, through the central United States, and into southern Canada (Shreve 1942, Rzedowski 1975, 2006, Brown 1982, Aguado-Santacruz & García-Moya 1998, Wick et al. 2015), making it the only continuous terrestrial ecoregion shared by all three countries. By 2015, however only 14 % of the original Chihuahuan Desert grasslands remained (CEC 2015). Like other parts of the world (Curtin & Western 2008), semiarid grasslands in Mexico are the least represented ecosystems in the system of natural protected areas (Challenger 1998).
In Mexico, the primary use of grasslands has been as rangeland for livestock raising (Miranda & Hernández-X. 1963, Challenger 1998, Rzedowski 2006, Challenger et al. 2009). This use has caused their transformation, and although details have not been sufficiently studied, there has been an obvious shift in rangeland composition and ground cover. This includes the loss of the species most palatable to livestock, along with an increase or appearance of less palatable or entirely undesirable species, such as toxic plants. Several areas exhibit severe erosion that has exposed the underlying duripan (Aguado-Santacruz & García-Moya 1998, Riojas-López & Mellink 2005, Medina-Roldán et al. 2007, 2008, Mellink & Riojas-López 2020). Also, grazing and browsing by livestock is a factor in the impaired condition of many of the world’s threatened species (Steinfeld et al. 2006). In Mexico animal populations have been severely impacted by overgrazing (Ceballos et al. 2010, Mellink & Riojas-López 2020), which has consequently negatively affected the provision of ecosystem services.
The southern part of the Mexican Plateau has not escaped the negative impacts of livestock grazing, which locally have ranged from strong to severe (Mellink & Riojas-López 2020). Given that grasslands receive low priority in conservation efforts, but are in urgent need of management focused on preserving their biodiversity and ecosystem services, research on their characteristics and processes at local scales is critically needed. In this article, we aim to gain a better understanding of the consequences of livestock grazing on the composition and ground cover of grasslands across a gradient from intensive grazing to no grazing, as well as at sites experiencing intermediate levels of grazing pressure, in the southernmost part of the North American grassland biome, within the southern section of the Mexican Plateau. The information we sought to obtain is intended to complement existing research and, together with it, contribute to the development of more effective grazing schemes for the region.
Materials and methods
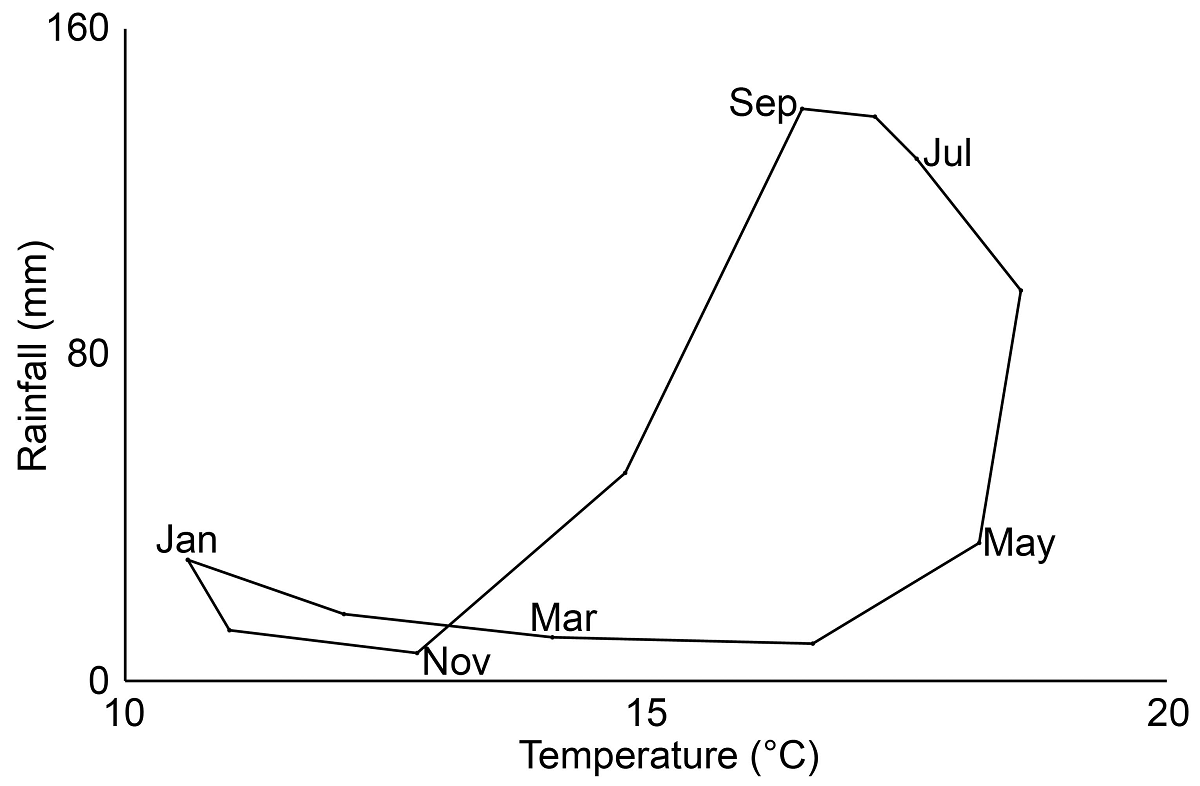
Study area. The study was carried out in the Llanos de Ojuelos, located in the southern part of the Central Plateau of Mexico, where the states of Jalisco, Zacatecas, Aguascalientes, San Luis Potosí and Guanajuato converge (center of the area at 21.9°, -101.7°). This semi-arid tableland, at an elevation of 1,900-2,600 m, with a geomorphology of low mountains and valleys (Nieto-Samaniego et al. 2005) has an average annual temperature, recorded at the Ojuelos de Jalisco, Jalisco, meteorological station (1969-2008), of 15 °C, with an average annual rainfall of 639 mm. Tank evaporation exceeds precipitation in every month of the year. The region experiences three distinct climatic seasons: cold dry, hot dry, and rainy (Mellink et al. 2016; Figure 1). The valley soils are of alluvial origin and contain an underlying layer of duripan cemented by alluvial silica, which is often exposed due to water and wind erosion, particularly when the vegetation is disturbed by grazing or plowing (Mellink & Riojas-López 2020).
The natural vegetation of the region is composed of grasslands, xerophilous shrublands, and dwarf oak stands (Quercus spp. L.) (Riojas-López & Mellink 2005). Grama (Bouteloua spp.), threeawns (Aristida spp. L.), and muhlies (Muhlenbergia spp.) are the most common components of grasslands, while catclaws (Mimosa spp. L.), silver dalea (Dalea bicolor Humb. & Bonpl. ex Willd.), leatherstem (Jatropha dioica Sessé), huisache (Vachellia spp. Wight & Arn.), arborescent nopales (Opuntia spp. (L.) Mill.), Peruvian peppertree (Schinus molle L.; peppertree from here on), and yucca (Yucca spp. L.) form the shrub and arborescent layers (Rzedowski & McVaugh 1966, Harker et al. 2008).
Livestock raising and agriculture are the main productive activities in the Llanos de Ojuelos, and they have transformed the region´s landscape since the arrival of Spanish conquerors 450 ~ 500 years ago (Riojas-López & Mellink 2005, 2023, Mellink & Riojas-López 2020). Over time, livestock in the region transitioned from cattle to sheep, and eventually to a mixture of sheep, goats, cattle, horses, and donkeys (Mellink & Riojas-López 2020). Currently, approximately 35.5 % of the surface of the municipalities of Ojuelos de Jalisco, Jal., and Pinos, Zac., is devoted to rain-fed farming, primarily of corn, beans and fruit-oriented nopal orchards, while sheep, goats, cows, and horses graze and browse throughout the region.
Study sites. We selected six study sites (Figures 2 and 3) that represented the full spectrum of local range conditions: from a research non-grazing enclosure area where grazing had been excluded for 36 years to a severely overgrazed range adjacent to it. Additionally, four other sites with varying histories of intermediate grazing pressure were included, two of which experienced reduction in grazing intensity several years ago after decades of severe overgrazing. Due to the availability of suitable study sites aligned with our objectives as well as logistic considerations, these locations were situated in two regions in the area: Vaquerías and Salitrillo de Chinampas (hereafter referred to as “Salitrillo”). These sites, arranged from west to east, were as follows:
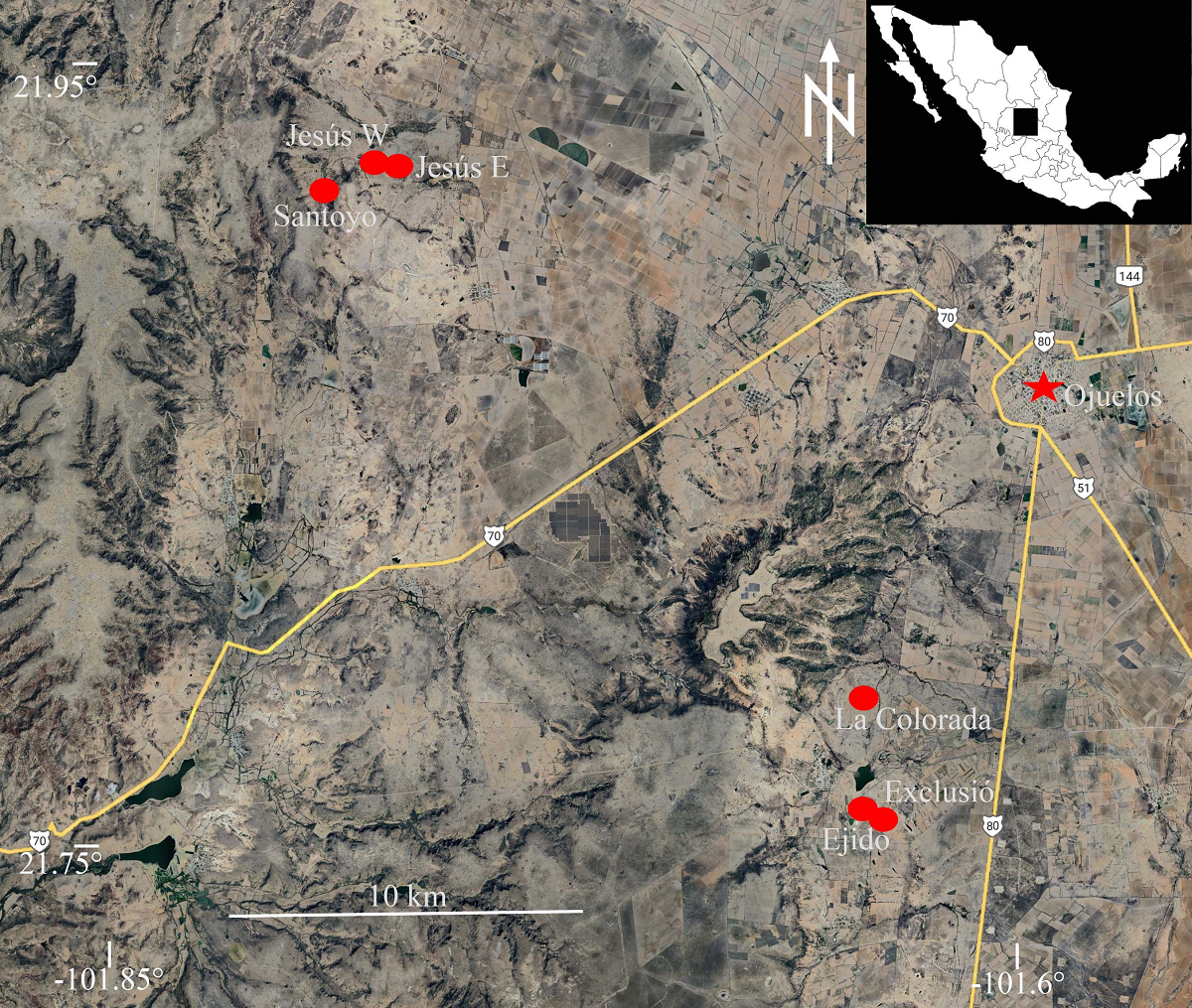

Figure 3 Study sites in the southern part of the Mexican Plateau: (A) Exclusión, (B) La Colorada, (C) Ejido, (D) Santoyo, (E) Jesús W, (F) Jesús E. Photographs taken 5-8 July 2016, after June rains had caused the greening of grasses.
Vaquerías region.- La Colorada (21.78755º, -101.64288º, 2,293 m asl; Figure 3B): This rangeland, part of a privately owned ranch, has been used since 2012 for the raising of fine horses at low density (M. Contreras com. pers.). Before then, it was subject to heavy grazing by cattle. In addition to grasses, there were nopal plants, yucas, and huisaches.
Exclusión (21.75893º, -101.64252º; 2,311 m asl.; Figure 3A). This was a 1-ha plot established in 1980 on severely overgrazed rangeland, from which livestock was excluded to study the effects of grazing. This site was referred to as “La Presa” by Aguado-Santacruz & García-Moya (1998), Aguado-Santacruz et al. (2002), and Delgado-Balbuena et al. (2013). Herbaceous vegetation entirely or nearly entirely covered the ground. The plot was surrounded by heavily grazed rangeland. In 2022, the fence protecting the site was torn down after the communal land of Ejido Vaquerías was divided into personal allotments, allowing grazing again.
Ejido (21.75657º, -101.63946º; 2,322 m asl; Figure 3C). This plot was on the range that surrounded Exclusión, belonged communally to the Ejido Vaquerías, and there was no control of the number of animals grazing the range. Grazing by cattle, sheep, goats, donkeys, and some horses was intense. The site had dispersed clumps of low-grazed grasses, cespitose grasses, and abundant jimmyweed (Isocoma veneta [Kunth] Greene), in addition to some yuccas.
Salitrillo región.- Santoyo (21.92045º, -101.79305º, 2,144 m asl.; Figure 3D). This site was on a private ranch. It used to be grazed heavily by beef cattle until 2001, when it was redirected to the production of bullfighting cattle, which are raised at much lower densities. Grasses were widespread, but huisaches and nopales were remnants of previous abuse. The study site was 100 m from a xeroriparian habitat.
Jesús W (21.92511º, -101.77673º, 2,222 m asl.; Figure 3E). This rangeland was part of the Fracción de Salitrillo de Chinampas, a privately owned ranch that was used for grazing by beef cattle. Grasses were widespread, and some huisaches were present.
Jesús E (21.92369º, -101.77058º, 2,173 m asl.; Figure 3F). This rangeland was also part of the Fracción de Salitrillo de Chinampas, but unlike the previous site, it was not grazed in 2016, when we performed this study. Grasses were widespread, and there were some huisaches.
Field work. We carried out monthly field assessments of vegetation at each site from July to November 2016, after the onset of summer rains, when plant growth was stimulated and grass species could be identified more accurately. To estimate cover across different categories, we established three 41-m transects at each site, radiating from a central point. Transects were oriented 120º from each other (Figure 4), with the first transect´s orientation randomized. Metal stakes were used to mark the start and the end points of each transect to ensure precise repetition in subsequent assessments. Starting at 1.2 m on each transect, we recorded the cover category at ground level every 20 cm, counting to the end of the transect (i.e., 600 survey points per site; after Herrick & Jones 2002). Cover categories included grass species, non-grass herbs, mulch, rock, and bare ground. Non-grass herbs were not identified individually and were grouped together. Additionally, the height class of all herbaceous plants was recorded in three categories: 10 cm, 11-30 cm, and > 30 cm). Patches along a transect 20 cm without vegetation were classified as open patches. Grass species were verified at the herbarium of the Universidad Autónoma de Aguascalientes (UAG), and specimens were deposited at both the herbarium of UAG and at the herbarium IBUG, Universidad de Guadalajara.
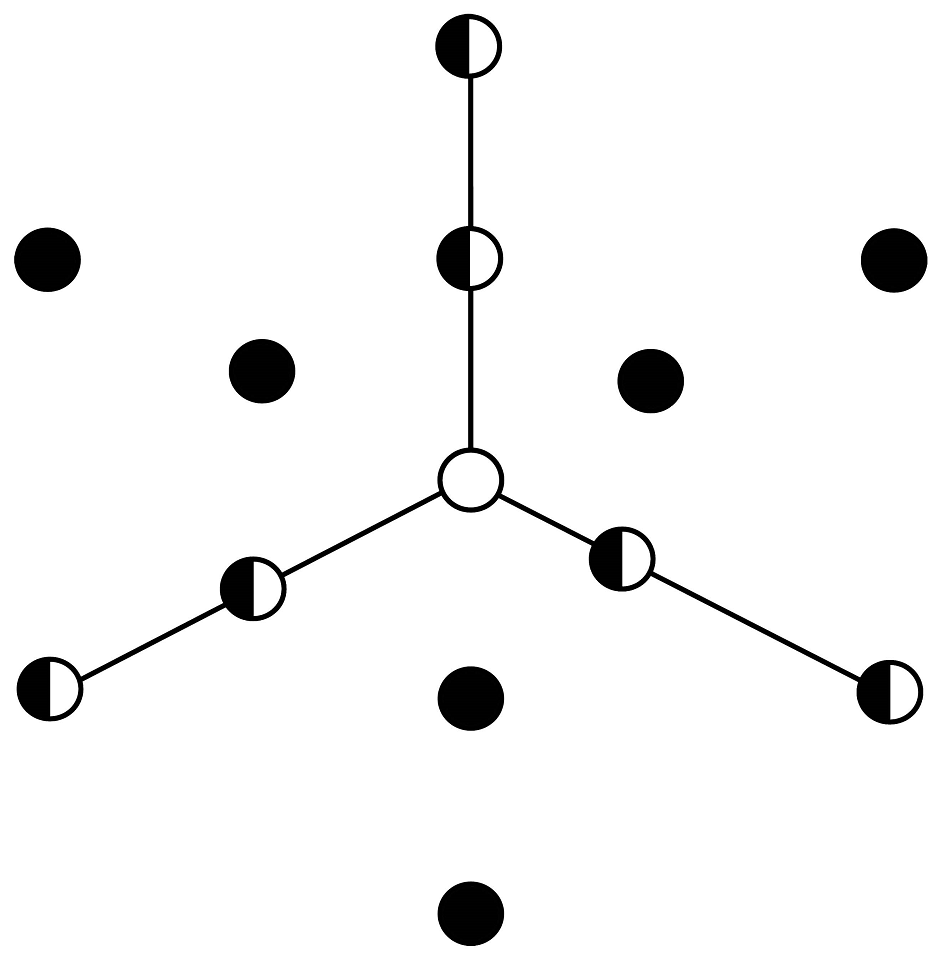
Figure 4 Survey scheme. Black dots indicate sites where soil resistance to penetration data was taken; the white dot, places from which a soil sample were taken, and black and white dots, where both were obtained.
Soil compaction, a known consequence of grazing, impacts plant root development, water infiltration, and overall vegetation growth (Cingolani et al. 2003, Chapin III et al. 2011). We assessed this attribute through soil resistance to penetration with a dynamic cone penetrometer (Herrick & Jones 2002) at 12 points within each study site (Figure 4). We recorded the number of hammer drops required to fully sink the cone tip into the ground. This measurement was taken in January 2017 to avoid the influence of soil moisture on penetration resistance.
Soils analyses were conducted to assess texture, organic matter, pH, electric conductivity, and macronutrients including nitric nitrogen, ammonia nitrogen, phosphorus, potassium, calcium, magnesium, and manganese. For analysis we collected a 1-kg soil composite sample, mixing seven subsamples, which were analyzed at the Laboratorio de Agrología of the Centro Universitario de Ciencias Biológicas y Agropecuarias, Universidad de Guadalajara, following NOM-021 standards.
Data analysis. Because grass species can be challenging to identify when they are not in bloom or seed, particularly amongst closely related species such as the blue grama (Bouteloua gracilis (Kunth) Lag. ex Griffiths) and Mexican plateau grama (B. scorpioides Lag.), we averaged the cover values for grasses in October and November, when they were more identifiable. In some instances, grasses encountered could be identified only at the genus level due to their growth stage. Subsequent surveys indicated that these grasses belonged to one of the species identified at other points and surveys. Therefore, to analyze inter-month variation, the hits recorded for threeawns, gramas and muhlies that were not identified to species were allocated to the corresponding identified species of these three genera based on proportion to the hits for the latter.
We compared the study sites and survey months based on numbers of hits for different cover categories and gap size using analysis of variance (ANOVA), followed by Tuckey post-hoc tests where appropriate. The high number of transect survey points precludes the need for data transformation. All tests were conducted with a significant level set at P ≤ 0.05, considering values of 0.054 or less as statistically significant. To evaluate grass species diversity, we calculated the combined cover values for October and November using the Shannon true diversity index (expH’, sensu Jost 2007).
Results
We recorded twenty-four species of grasses (Tables 1 and S1). Grama grasses, collectively, were significantly the most abundant herb species (P < 0.001), while there were no significant differences between the other plant cover categories. As the rainy season developed, grasses reflected it by growing taller, increasing ground cover, and developing the circannual pattern of vegetative growth, flowering, seeding, followed by drying, decay, and a reduction of plant height and cover at the end of the season (Figure 5). Rainfall also stimulated the appearance of six annual species (Table 1), although their combined cover never exceeded 2 %. The largest cover by these annual species was provided by exotic crabgrass (Digitaria ternata,[A.Rich.] Stapf; also known as black-seed finger grass), which accounted for 90 % of all cover by annuals in October and 79 % in November. The increase in perennial species after the rain was mostly due to grama grasses, notably sprucetop grama (Bouteloua chondrosioides(Kunth) Benth. ex S.Watson). Grass diversity values (expH’) were highest at Jesús E and lowest at Jesús W (Figure 6).
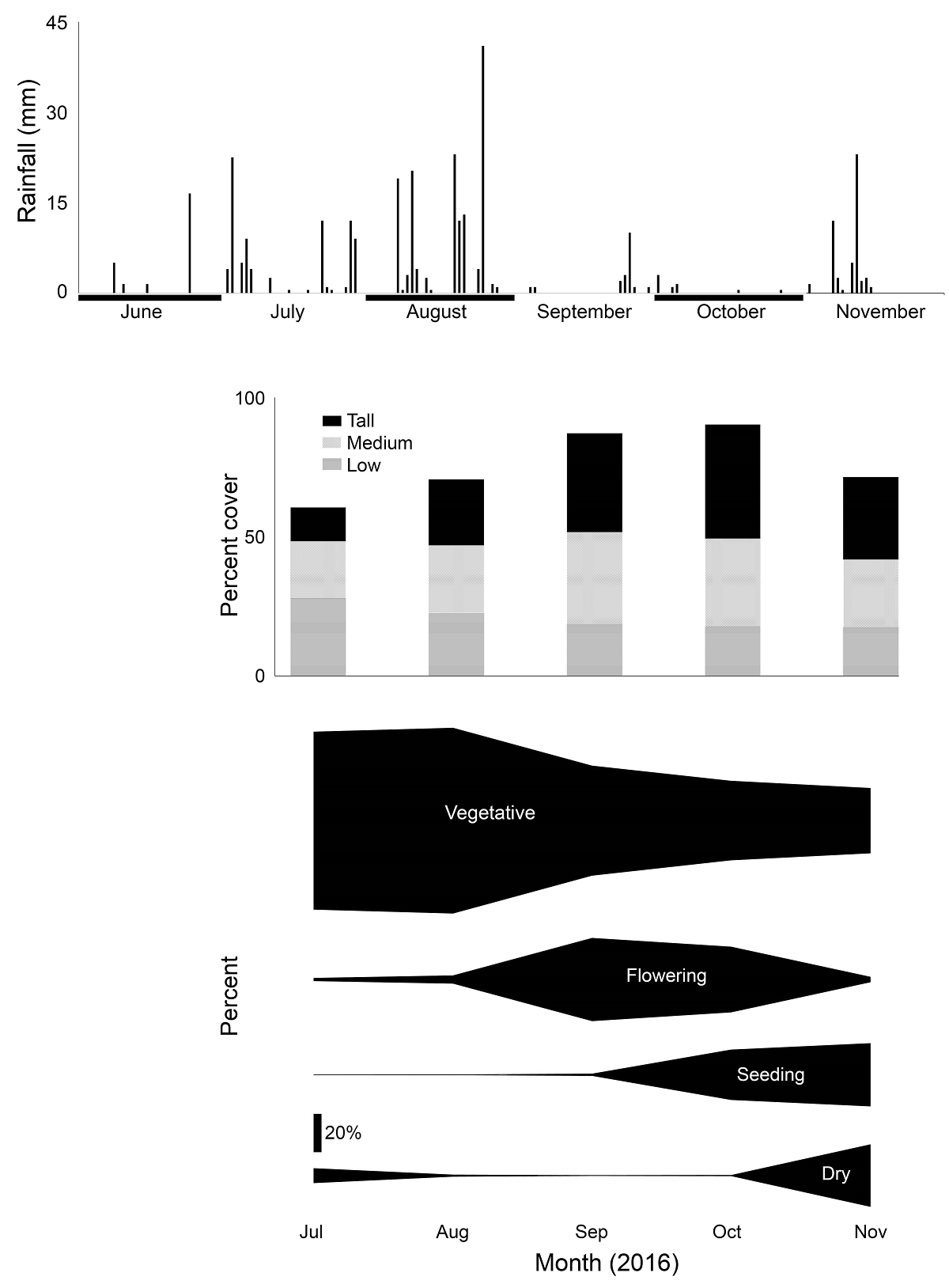
Figure 5 2016 summer rainfall, plant cover, and plant phenological stage in six grasslands in the southern part of the Mexican Plateau.
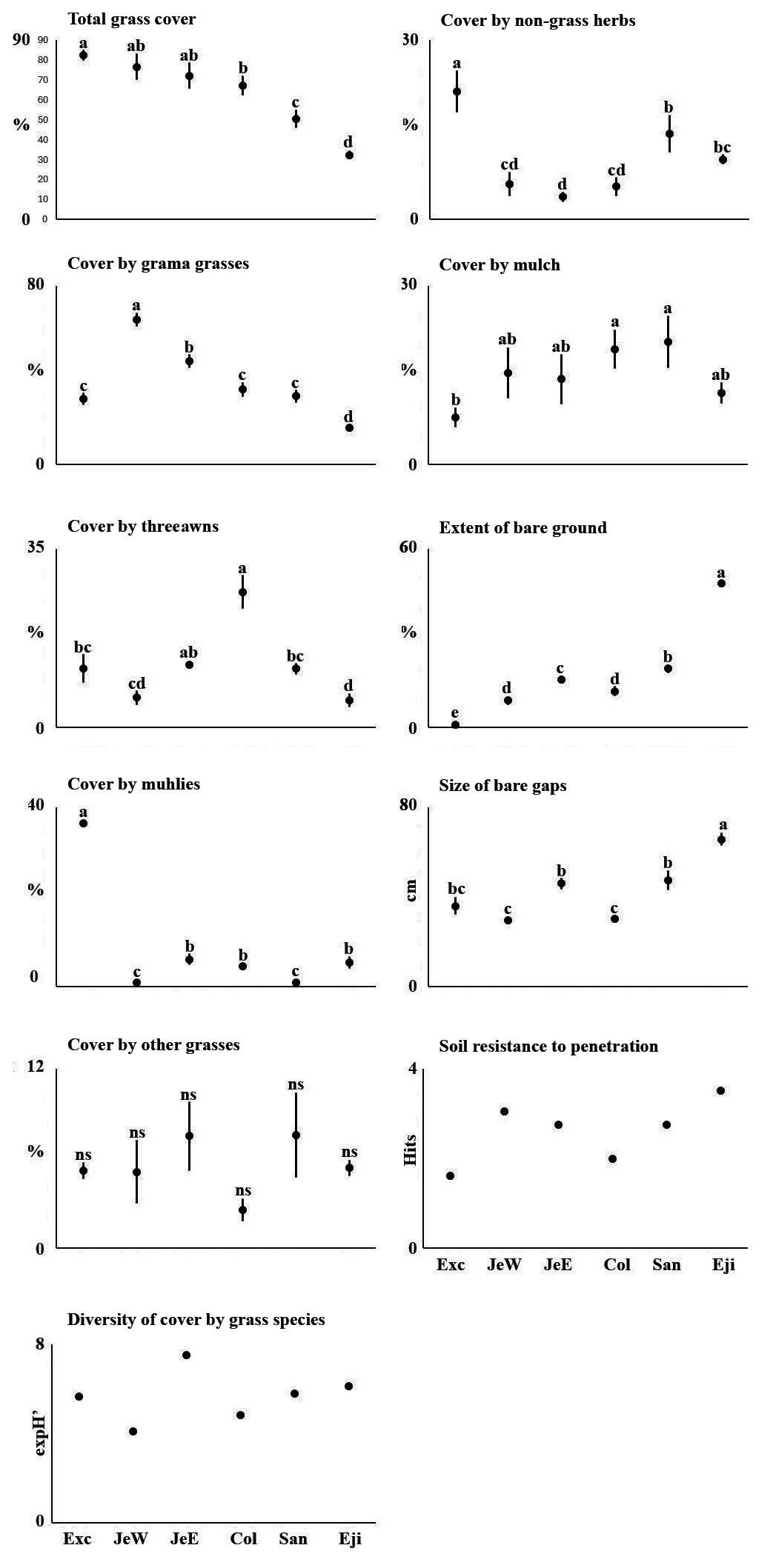
Figure 6 Cover by plants, bare soil and soil resistance to penetration in six rangeland plots with different grazing history in the southern part of the Mexican Plateau.
Table 1 Plan components, especially grasses identified on six range sites in the southern part of the Mexican Altiplano, 2016. Values under each locality are percent ground cover by the species. The sum of bare ground, mulch and the different species can add up to >100, as species can overlap in their cover.
| Component | Locality | |||||
|---|---|---|---|---|---|---|
| Exclusión | Jesús W | Jesús E | La Colorada | Santoyo | Ejido | |
| Bare ground | 1 | 9 | 16 | 12 | 20 | 48 |
| Mulch | 8 | 15 | 14 | 19 | 21 | 12 |
| Grasses | ||||||
| Sixweeks threawn (Aristida adscensionis) | 0 | < 1 | 0 | 0 | 0 | 0 |
| Poverty threeawn (Aristida divaricata) | 12 | 6 | 12 | 26 | 12 | 5 |
| Generic threeawn (Aristida sp.) | 0 | 0 | 0 | < 1 | 0 | 0 |
| Spider grass (Aristida ternipes) | < 1 | 0 | < 1 | < 1 | 0 | < 1 |
| Cane bluestem (Botriochloa barbinodis) | 3 | 0 | 0 | 0 | 0 | 0 |
| Sprucetop grama (Bouteloua chondrosioides) | 0 | 13 | 4 | 0 | 1 | 0 |
| Sideoats grama (Bouteloua curtipendula) | 0 | 1 | 0 | 0 | 0 | 0 |
| Buffalo grass (Bouteloua dactyloides) | 0 | 3 | 1 | 0 | 2 | < 1 |
| Blue grama (Bouteloua gracilis) | 3 | 22 | 20 | 7 | 7 | < 1 |
| Hairy grama (Bouteloua hirsuta) | < 1 | 1 | 0 | 3 | 0 | 5 |
| Mexican plateau grama (Bouteloua scorpioides) | 3 | 0 | 3 | 9 | 0 | 3 |
| Mat grama (Bouteloua simplex) | 0 | 0 | 0 | 0 | 0 | 0 |
| Generic grama (Bouteloua sp.) | 24 | 26 | 19 | 15 | 21 | 7 |
| Feather fingergrass (Chloris virgata) | 0 | 0 | 0 | 0 | 0 | 0 |
| Crabgrass (Digitaria ternata) | 0 | < 1 | 1 | < 1 | 2 | < 1 |
| Plains lovegrass (Eragrostis intermedia) | < 1 | < 1 | 1 | < 1 | < 1 | 1 |
| Mexican lovegrass (Eragrostis mexicana) | 0 | 0 | < 1 | 0 | 0 | 0 |
| Kunth's small grass (Microchloa kunthii) | < 1 | 5 | 2 | 1 | 4 | 2 |
| Wolftail grass (Muhlenbergia phleoides) | 5 | 1 | 6 | 3 | 1 | 5 |
| Creeping muhly (Muhlenbergia repens) | 0 | 0 | 0 | 0 | 0 | 0 |
| Purple muhly (Muhlenbergia rigida) | 31 | < 1 | 0 | 1 | < 1 | 0 |
| Prickly needlegrass (Nassella mucronata) | < 1 | 0 | 0 | 0 | 0 | 0 |
| Generic panicgrass (Panicum sp.) | 0 | 0 | < 1 | 0 | 1 | < 1 |
| Mexican paspalum (Paspalum convexum) | 0 | 0 | < 1 | 0 | < 1 | 0 |
| Pringle hairgrass (Peyritschia pringlei) | 1 | 0 | < 1 | 0 | 0 | 0 |
| Marsh bristlegrass (Setaria parviflora) | 0 | < 1 | 1 | 0 | 0 | < 1 |
| Unidentified grass | < 1 | < 1 | 1 | 1 | < 1 | 1 |
| Non grasses | ||||||
| Herbs | 21 | 6 | 4 | 6 | 14 | 10 |
| Huisache (Vachellia sp.) | 0 | 3 | 0 | 0 | 9 | 0 |
In October and November, when grass species were most identifiable, of all survey points in which grass was intercepted, 54 % were of grama, 21 % of threeawns and 14 % of muhlies (Figure 7). Individually, at this time, blue grama (21 % of all grass) was the most abundant species, followed by the poverty threeawn (Aristida divaricata Humb. & Bonpl. ex Willd; 20 %), sprucetop grama (15 %), Mexican plateau grama (15 %), and purple muhly (Muhlenbergia rigida (Kunth) Kunth, 8 %; Table 1). Panic grass could not be identified at species level as no validated equivalents were in the herbarium.
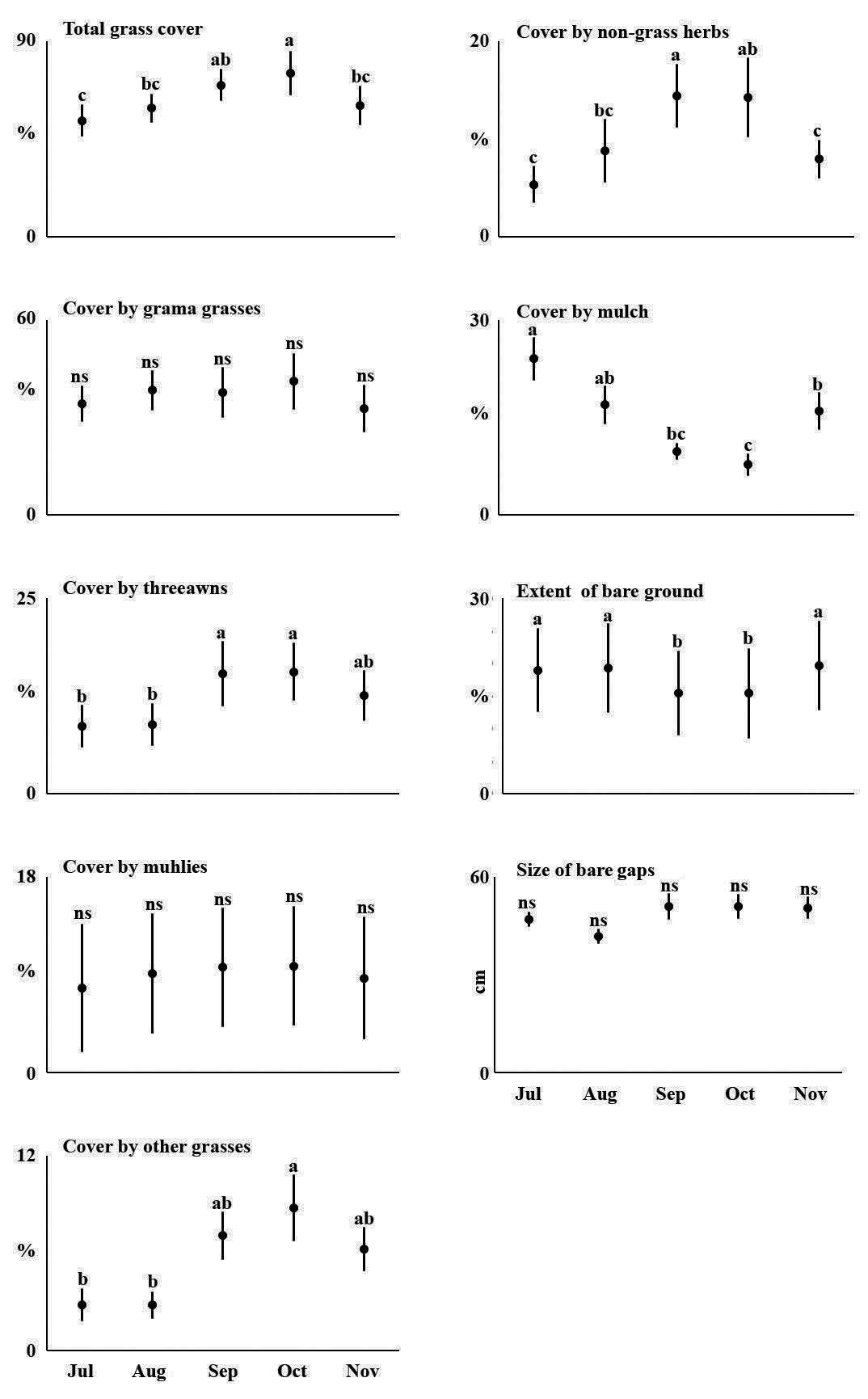
Figure 7 Cover by plants, bare soil and soil resistance to penetration from July to November in six rangeland plots with different grazing history in the southern part of the Mexican Plateau.
Grama grasses were the most common component in five of the six sites, but each site exhibited individual differences (Figure 6; P ≤ 0.01; Table S3). Excluding the highly overgrazed Ejido site, grama was the major component in both Jesús E and W, while purple muhly was the dominant species in Exclusión (P ≤ 0.01) where it had a cover severalfold that in other sites. Three awns were significantly more abundant in La Colorada (P ≤ 0.01). In the different sites, between 4 and 23 % of the ground was covered by non-grass herbs. These were significantly more abundant in Exclusión (P ≤ 0.01), where major non-grass herbs were candy leaf (Stevia sp. Cav.), red-crescent marigold (Tagetes lunulata Ortega), pine-needle milkweed (Asclepias linaria Cav.), and yerba de pasmo (Baccharis pteronioides DC.). At the heavily grazed Ejido site, jimmyweed was the dominant plant, but other species like silver ponysfoot (Dichondra argentea Humb. & Bonpl. ex Willd.), onionweed (Asphodelus fistulosus L.) were also present. Santoyo, which was intermediate in its cover by non-grass herbs, had some jimmyweed, pine-needle milkweed, dogweed (Dyssodia papposa (Vent.) Hitchc.), Irish lace (Tagetes filifolia Lag.), and Fort Huachuca vervain (Verbena gracilis Desf.).
Between 1 and 49 % of the ground at the study sites was bare, going from the significantly less bare Exclusión, with softer soil, to the significantly barest Ejido (P ≤ 0.01), with the hardest soil. The other four sites had between 9 and 19 % bare ground and intermediate soil hardness values. Ejido had significantly larger bare gaps (66 ± 2.8 cm, P ≤ 0.01) than the other study sites (Figure 6).
The major components of the plant community were perennial grasses, and temporal changes in plant cover were slight (Figure 7). These were limited to threeawns (P ≤ 0.01), non-identifiable grasses (P = 0.01), and non-grass herbs (P ≤ 0.01). The latter two groups included annual species. Although slight, these changes were reflected in the extent of bare ground (P ≤ 0.01).
The ground cover by some species was correlated either positively or negatively. The abundance of sprucetop grama was correlated positively with those of blue grama (r2 = 0.86) and Kunth's small grass (Microchloa kunthii Desv.) (r2 = 0.84), and negatively with that of Mexican plateau grama (r2 = -0.73). Additionally, Mexican plateau grama was negatively correlated with Kunth's small grass (r2 = -0.96) and huisache (r2 = -0.83); Kunth's small grass was also correlated negatively with wolftail grass (Muhlenbergia phleoides (Kunth) P.M.Peterson) (r2 = -0.71) and positively with huisache (r 2 = 0.72); wolftail grass was correlated negatively with huisache (r2 = -0.71); and purple muhly was correlated positively with non-grass herbs (r2 = 0.86).
All sites had loamy soils, and differences in soil characteristics between them were slight (Table S2). Clay, percentage of organic matter (OM), pH, percentage of available water (AW), and calcium (Ca) exhibited some differences between study plots. Clay content, OM, Ca, and AW content of the soil were higher at the Salitrillo region study sites, while there were differences between the Vaquerías region sites. The soils of all sites had close to neutral pH, with the three Salitrillo region sites and Exclusión being slightly acidic (pH = 6.3 ≈ 6.6), while the two other Vaquerías region sites being slightly alkaline (pH = 7.1 and 7.23).
Resistance to penetration ranged between 1.4 and 3.1 hammer drops at the different sites (Figure 6) and was correlated positively with total bare ground (correlation coefficient r2 = 0.8) and with the sum of the length of open patches (r2 = 0.67), and negatively with total cover by plants (r2 = -0.72), cover by poverty threeawn (r2 = -0.75), and cover by purple muhly (r2 = -73). Resistance to penetration was not correlated with the percentages of different soil texture classes, or with soil organic matter content (r2 > 0.15 in all cases).
Discussion
All native grass species recorded in this study are typical of rangelands in the southern part of the Mexican Plateau, where grama grasses, notably the blue and Mexican plateau gramas, are the most conspicuous group of grasses. Two of the species recorded in this study were exotic species. Crabgrass is an invasive species from the tropical and subtropical regions of Africa and Asia that had not been reported in the U.S. in 1950 (Hitchcock 1950) and appears to have been introduced more recently to Mexico (Beetle 1987). The unidentified panic grass was very likely kleingrass (Panicum coloratum L.), an African species that was introduced in 1978 as part of “range improvement” experiments to a research station 4 km distant from our study sites in the Vaquerías region (Negrete et al. 1981).
Dryland grasses respond to climate in a circannual phenological pattern and, not surprisingly, the main factors of grassland structure and composition in this area have been found to be rainfall, mostly through its effect on annual herbs, and grazing, with soil characteristics being of secondary importance (Aguado-Santacruz & García-Moya 1998, Aguado-Santacruz et al. 2002). Although this is well known by rangers and cattle herders in drylands worldwide, perhaps because of its obviousness it is absent from the scientific literature, at least for northern Mexico. Our data exhibited a clear phenological pattern that supersedes the effects of differences between ranges (Figure 5). This phenological progression caused increases in overall grass cover and its height as a result of rainfall, but such increases in cover were slight, reflecting that the majority of grass cover in the region consists of perennial species.
The major drivers of the composition of grassland communities in semiarid grasslands are grazing history and weather (Aguado-Santacruz et al. 2002, Mellink & Riojas-López 2020). Aguado-Santacruz et al. (2002) indicated that climate is the primary driver, while we argued that grazing history is more important (Mellink & Riojas-López 2020). This difference in opinion is related to the geographical and climatic scales used: the prior authors based their conclusions on 11 years of vegetation monitoring, while ours is a historical reconstruction of 500 years. The results presented in this contribution derive from one growing season; hence they primarily reflect grazing history, although there appears to be some influence of soil characteristics.
According to García Moya & Villa (1977) and Aguado-Santacruz & García-Moya (1998), blue grama should be considered the climax species of regional grasslands, and its cover should therefore be used to assess range condition in accordance with the standard procedure in range management (Dyksterhuis 1949, Huss & Aguirre 1979). Based on this, the study sites of Jesús W (28 % of relative cover by blue grama) and Jesús E (25 %) would be classified as fair, while Santoyo (21 %), La Colorada (13 %), Exclusión (11 %), and Ejido (3 %) would be classified as poor. However, Exclusión, with its exuberant herbaceous plant community including muhlies and non-grass herbs, does not fit the description of a range in poor condition given its nearly complete ground cover by grasses and non-grass herbs, and softer soil compared to the other sites.
At the other end of the plant cover spectrum is Ejido, which had significantly more bare ground (49 %), larger gap size, and harder soil. Its depleted plant community was composed mostly of hairy grama (Bouteloua hirsuta Lag.), wolftail grass, threeawn, and Mexican plateau grama. These species have been identified as subordinated grass species that replace blue grama under severe grazing (Aguado-Santacruz & García-Moya 1998, Aguado-Santacruz et al. 2002, Medina-Roldán et al. 2007, Delgado-Balbuena et al. 2013). Consistent with this, our data exhibited a negative relationship between blue and Mexican plateau gramas. Jimmyweed, the visually dominant low shrub at Ejido, is a major indicator of overgrazing in the region, and its dominance reflects the severe deterioration of the grassland at this site. Its presence at the other sites could perhaps be a ghost of past overgrazing, that under current use will surely take a long time to disappear. The plant community at Ejido suggests that hairy grama and wolftail grass indicate even more severe overgrazing conditions, than threeawn and Mexican plateau grama.
The other four study sites had range conditions between those of Exclusión and Ejido. Of these, Santoyo, located near a permanent stream, had a lower content of sand and higher nitrogen content, possibly explaining its larger cover by non-grass herbs, second only to Exclusión. From a range condition viewpoint, La Colorada could be classified as in worse condition than Jesús W and Jesús E due to its higher ground cover of poverty threeawn, despite that grazing pressure has eased in the last 20 years and is currently lower than at the latter two sites. Conversely, the dominance of poverty threeawn at la Colorada might be caused by the siltier and drier soil (Beetle 1983), and perhaps by its lower calcium content.
Available water capacity was higher at the three Salitrillo study plots. In Jesús E and Jesús W, this may be related to the higher cover of blue grama (Medina-Roldán et al. 2007), and at Santoyo to its proximity to a perennial stream. However, this explanation is complicated by the fact that the Salitrillo study plots have higher clay content, which was positively correlated with water availability across our sites (r2 = 0.95).
A higher diversity in ecological communities is generally considered positive from an ecological standpoint (Gaston & Spicer 2004), but from a traditional range management perspective, high diversity in rangelands is not necessarily associated with the most desirable range condition. Instead, high dominance (hence low diversity), of good forage species is preferable. In rangelands subject to use by domestic herbivores, species that are more resistant to grazing tend to dominate, leading to lower diversity. In our study, the dominance of purple muhly in Exclusión, the most mature ecological community, had also the same effect: a low diversity value.
Soil hardness is one of the range attributes commonly affected by heavy grazing pressure, reflecting the history of range use. In our study, the hardest soil was at the most grazed site (Ejido) while the softest soil was at Exclusion. If grasslands with more poverty threeawn and less blue grama are more impacted by grazing, as indicated in the literature, La Colorada should have had a soil surface more resistant to penetration than Jesús E and Jesús W; but the opposite was true (Figure 6). This apparent contradiction questions the “more threeawn - less blue grama” assumption and requires further study. The softer soils at La Colorada could reflect the approximately 20 years of reduced grazing pressure.
The selection of blue grama as the climax indicator species for the region’s grasslands was based on a temporal scale limited to the past couple of centuries. Until our survey, there was no evidence suggesting that any grass species other than blue grama should be considered so. Even in 2008-2009, after nearly 30 years without grazing, blue grama remained the main component of Exclusión (Delgado-Balbuena et al. 2013). However, 7-8 years later, purple muhly dominated the grass community in same site. We have contended that this may be a closer representation of the grasslands´ composition prior to cattle introduction (Mellink & Riojas-López 2020).
We acknowledge that we cannot be completely certain whether purple muhly was the dominant pre-cattle species, but this is so far this is so far our best approximation of what the mature stage of the disclimax succession in the southern Mexican Plateau grasslands would have been in the absence of large herbivores (sensu Martin 1975). If purple muhly was a dominant species in many of the region’s grasslands, being an excellent (Beetle et al. 1991) to good forage species when young (Herrera Arrieta & Pámanes García 2010), it was likely a decreaser (sensu Dyksterhuis 1949, Huss & Aguirre 1979), avidly consumed, allowing the more grazing tolerant blue grama to increase. The recent destruction of Exclusion precludes further investigation of pre-cattle grassland conditions. Even if other non-grazing enclosures were established and enough time allowed to elapse, the current development of a global climatic change will interfere with the re-establishment of pre-domestic livestock conditions.
Our data supports the view that dense prairies, rather than grasslands, dominated the region in the XVI Century, consisting of non-grass herbs as well as grass, likely with purple muhly rather than grama grasses as the dominant species. This view aligns with historical accounts that “grasses touched the bellies of the horses” at that time (Crosby 1972). Through initial grazing, purple muhlies and other highly palatable species were gradually replaced by grama grasses. Given their adaptation to grazing, grama grasses persisted much longer in the systems, as was the case in the Salitrillo sites, until grazing pressure became extreme at the XX Century, with the consequences observed at Ejido.
Most of the region´s grasslands are overgrazed, many heavily so, but although different ranges have different grazing histories, lack of stocking records and their change through time precludes analysis of the relationship between grazing intensity and range condition. The Exclusion non-grazing enclosure provided crucial information on this relationship until its destruction in 2022. This loss is irreparable.
Finally, the need for grassland/prairie restoration is undeniable, but managers and conservationists face the question on which scenario to use as a target: the grama grasslands of 200 years ago? or the prairie of 500 years ago? Regardless of the scenario aimed at, it must be pursued through regenerative grazing schemes, as it would be impossible and socially insensitive and unacceptable, to cease ranching in the region: A balance must be struck between conservation objectives and livestock production.
Supplementary material
Supplemental data for this article can be accessed here: https://doi.org/10.17129/botsci.3586.
Supplemental material 1 and 2.

