nueva página del texto (beta)
nueva página del texto (beta) Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink1. Introduction
As a result of the Early Cimmerian tectonic event, Iran is separated into two geologically independent zones; one in the northeast (comprising central Iran, northern and northeastern Iran) and the other in the southwest (Zagros area). These two distinct sedimentary-structural zones comprise the Jurassic strata of remarkably variable litho- and biofacies.
The late Triassic-Middle Jurassic deposits in the former zone are characterized by the Shemshak Formation bounded by two unconformities. The Shemshak Formation (Assereto, 1966) comprises a thick siliciclastic succession, widely distributed across central and northern Iran, the so-called Iran Plate (Seyed-Emami, 2003; Aghanabati, 2004; Seyed-Emami et al., 2008; Fürsich et al., 2009a). Generally, the Shemshak Formation rests unconformably on the Lower-Middle Triassic platform carbonates of the Elika Formation and in turn is followed by basinal carbonates of the Middle Jurassic Dalichai Formation. Much of the Shemshak Formation sedimentation in the Alborz Ranges took place on coastal plains in fluvial, lacustrine, or deltaic settings, paralic swamps, lakes, meandering as well as braided rivers, and shallow-deep shelfal environments (e.g., Seyed-Emami, 2003; Seyed-Emami et al., 2001, 2005, 2006, 2008; Fürsich et al., 2005, 2009a; Sajjadi and Hakimi Tehrani, 2009; Sajjadi et al., 2010). Due to an apparently humid climate, swamps were widespread, which is reflected by abundant and economically important coal deposits (Repin, 1987).
The age of the Shemshak Formation spans the Late Triassic-Middle Jurassic, deduced mostly from fossil plants (e.g., Barnard, 1968; Fakhr, 1977, Barnard and Miller, 1976; Schweitzer, 1977, 1978; Achilles et al., 1984; Schweitzer and Kirchner, 1995, 1996, 1998, 2003; Schweitzer et al., 1997, 2000, 2009; Badihagh and Uhl, 2019; Sadeghi and Hashemi, 2021), ammonites (e.g., Seyed-Emami et al., 2001, 2005, 2006), belemnites (e.g., Parent et al., 2013) and bivalves (Aghanabati, 2014).
Despite a wide areal extension of the Shemshak Formation on the Iran Plate, palynological investigations previously attempted are not widespread (Kimyai, 1975; Achilles et al., 1984; Bharadwaj and Kumar, 1988; Sajjadi and Hakimi Tehrani, 2009; Sajjadi et al., 2010; Ghasemi-Nejad et al., 2004, 2008). This scarcity is mostly due to the detrimental effect of heat/temperature, leading to the formation of coal seams within the rock unit. The objective of this research is to record the palynological characteristics of the Shemshak Formation located near Namin in Ardabil Province (western Alborz). This study aims to evaluate its stratigraphic, paleobotanical, paleoecological, and paleobiogeographical importance, while also comparing it with other Middle Jurassic data from Eurasia and Gondwana.
Palynostratigraphy is a well-recognized and effective biostratigraphic method (Traverse, 2007) that also serves to reconstruct plant communities, aiding in the interpretation of paleoenvironmental and paleoclimatic conditions (e.g., Abbink et al., 2004; Volkheimer et al., 2009). Investigating in situ palynomorphs within plant macrofossils creates a crucial connection between dispersed miospores and their corresponding botanical relationships (Filatoff, 1975; Balme, 1995).
2. Geological Setting and Stratigraphy
The Upper Triassic-Middle Jurassic Shemshak Formation or Group is one of the most widespread lithostratigraphic units of the Iran Plate. This Plate is part of the Cimmerian terranes, sandwiched between the Turan Plate in the north (part of Eurasia) and the Zagros fold belt in the south (part of Gondwana). The Iran Plate became detached from the northeastern margin of Gondwana during the Early Permian (Stampfli and Borel, 2002) and moved northwards during the Triassic, thus closing off the Paleotethys Ocean regionally. (e.g., Stampfli and Borel, 2002).
This orogenic movement has been termed the Early Cimmerian Orogeny. This event brought about a drastic change in the sedimentary regime as the orogeny built the Cimmerian mountains along the Paleotethys suture zone. Denudation of this mountain chain produced large amounts of sediment that collected in an extensive foreland basin situated to the south. The resulting rock unit is the Shemshak Formation or Group. Due to its vast thickness in eastern Alborz (up to 4000m), some have elevated the unit to Group rank (e.g., Aghanabati, 1998; Seyed-Emami, 2003; Fürsich et al., 2009a).
Consequently, the unit has been subdivided into various members/formations, and different lithostratigraphic schemes have been proposed (Nabavi and Seyed-Emami, 1977; Nabavi, 1980; Repin, 1987; Aghanabati, 1998). In the western part of the Alborz Ranges, the Shemshak Formation can be adopted. In this study, we utilize the formation usage.
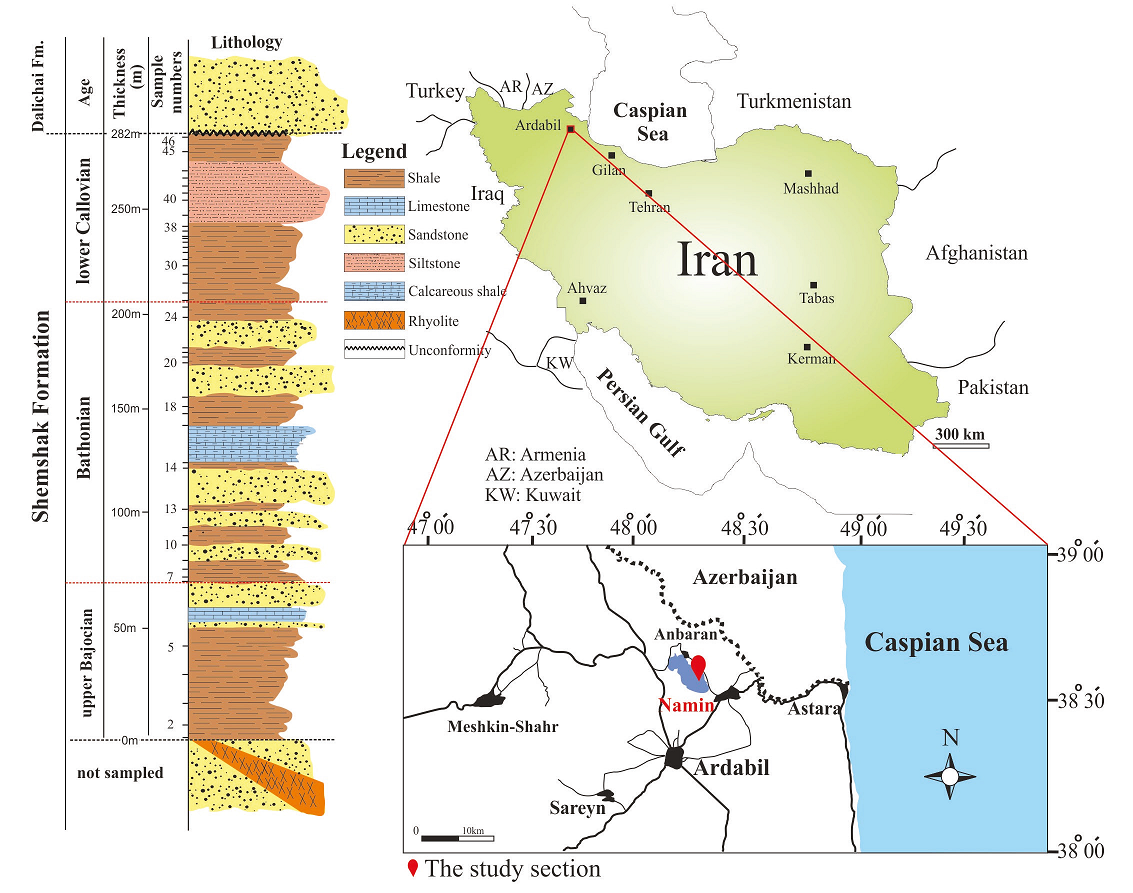
In most areas, the Shemshak Formation lies unconformably on the Elika Formation, but locally it may overlie older beds (e.g., Permian or even older rocks). In turn, it is overlain by the Dalichai-Lar formations from which it is separated by the Mid-Cimmerian tectonic event (Fürsich et al., 2009b). The Shemshak Formation is considered to be diachronous at both lower and upper boundaries (Assereto, 1966); the rock unit is collectively attributed to the Rhaetian-Middle Jurassic (Aghanabati, 1998). The upper limit of the Shemshak Formation apparently possibly extends to the Callovian in western Alborz Ranges as the succeeding Dalichai Formation commences from the Callovian (Aghanabati, 1998). The section investigated is situated northeast of Ardabil, 5 km northwest of Namin, Ardabil Province; western Alborz Ranges (Figure 1), the coordinates of the base of the section are N38°35′26″ and E48°20′33″. There, the rock unit here mainly consists of 282m of alternation of sandstones, siltstones, shales, limestones and calcareous shales. The lower boundary is covered whereas the upper limit is unconformable with the Middle Jurassic Dalichai Formation. There is a rhyolite vein in the lower part of the section studied (Figure 1).
3. Material and methods
Forty-six samples are collected from the shale and siltstone successions of the Namin stratigraphic section (Figure 1). Standard palynological procedures (e.g., Phipps and Playford, 1984, Wood, 1996) were applied for extraction and concentration of the palynomorphs. Samples were firstly disaggregated into pea-sized pieces (1-2 mm in diameter) and treated with 30% HCl to eliminate carbonates. The residue was then washed to neutrality and the remaining inorganic matter dissolved in HF (ca. 40%); fluoride precipitants formed during this step were removed using hot 30% HCl followed by washing the residue to neutrality. The organic residue was sieved via a 20μm mesh.
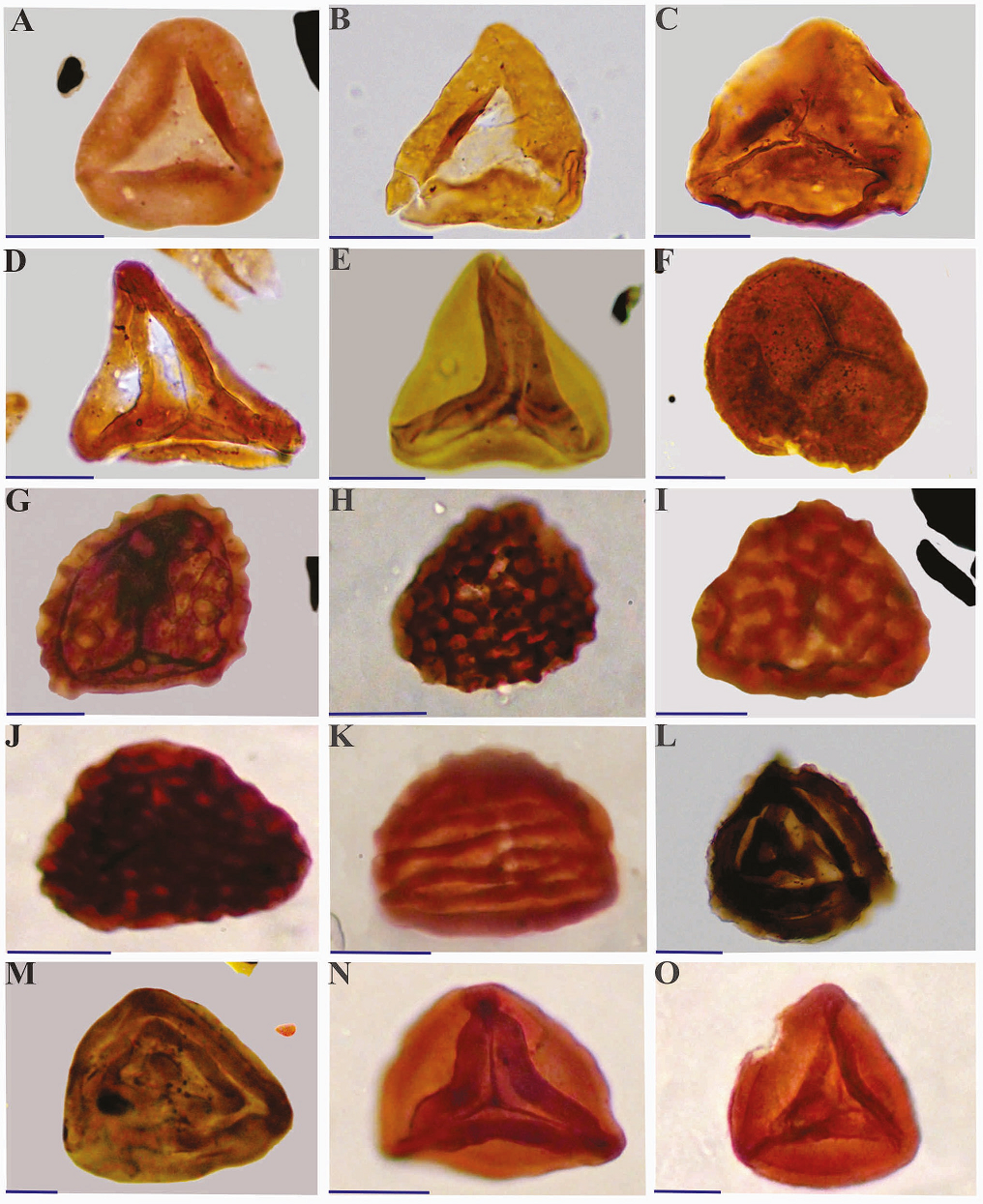
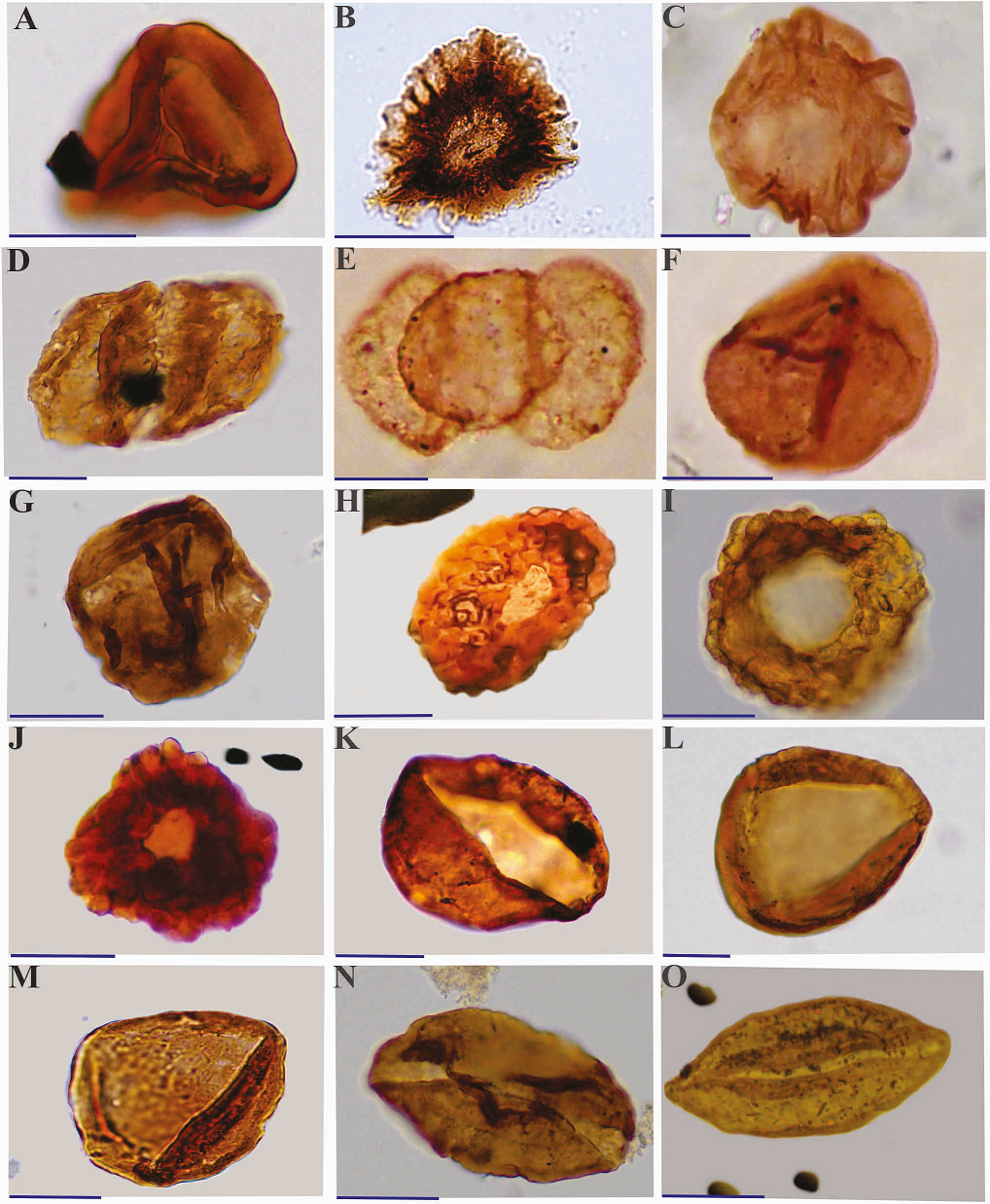
Heavy minerals and other remaining inorganic particles are removed by the use of a heavy-liquid solution (ZnCl2) at appropriate specific gravity. The remaining residue was then sieved through a 20μm nylon sieve before mounting on slides. At least three permanent strew slides per sample were prepared using Entellan as the mounting medium. An Olympus BH-2 microscope of the School of Geology, College of Science, University of Tehran, equipped with a mechanical stage and automatic Leitz camera facility, was employed for light microscopy and 35mm photography. Representative taxa are illustrated in Plates I-III. All rock samples, residues, and strew slides used in this investigation are permanently housed in the University of Tehran Palynology Collection (UTSH 1-46) at the School of Geology, College of Science, University of Tehran, Tehran, Iran.
4. Characteristics and stratigraphic significance of the Shemshak palynofloras
Summarized below is a qualitative complexion of the palynofloras assemblages retrieved from the Shemshak Formation followed by taxonomical comparisons that may be drawn with approximately coeval palynological assemblages from Iran and elsewhere. An appraisal of the age of the rock unit based on palynological data is also attempted.
4.1. GENERAL FEATURES
Apart from virtually no dinoflagellates cysts intervals at the uppermost part of the section studied, the majority of the subject samples are palyniferous containing reasonably diverse and reasonably preserved palynofloras of terrestrial and marine derivation. Most of the samples are dominated by miospores and marine palynomorphs (dinoflagellate cysts), rare acritarchs, foraminiferal test linings, and fungal spores in good states of preservation. The assemblages comprise 44 spore species (30 genera; 41.06%), 19 pollen species (eight genera; 12.56%), and 20 dinoflagellate cysts (11 genera; 46.25%).
A general summary of the qualitative and quantitative features of the palynofloral assemblages retrieved from the upper part of the Shemshak Formation follows. The palynological contents are productive and reasonably preserved thus allowing documenting and appraising the stratigraphic significance of the recovered miospores and dinoflagellate cysts and, in turn, providing clues on the Middle Jurassic paleogeography and paleoecology of northwestern Iran.
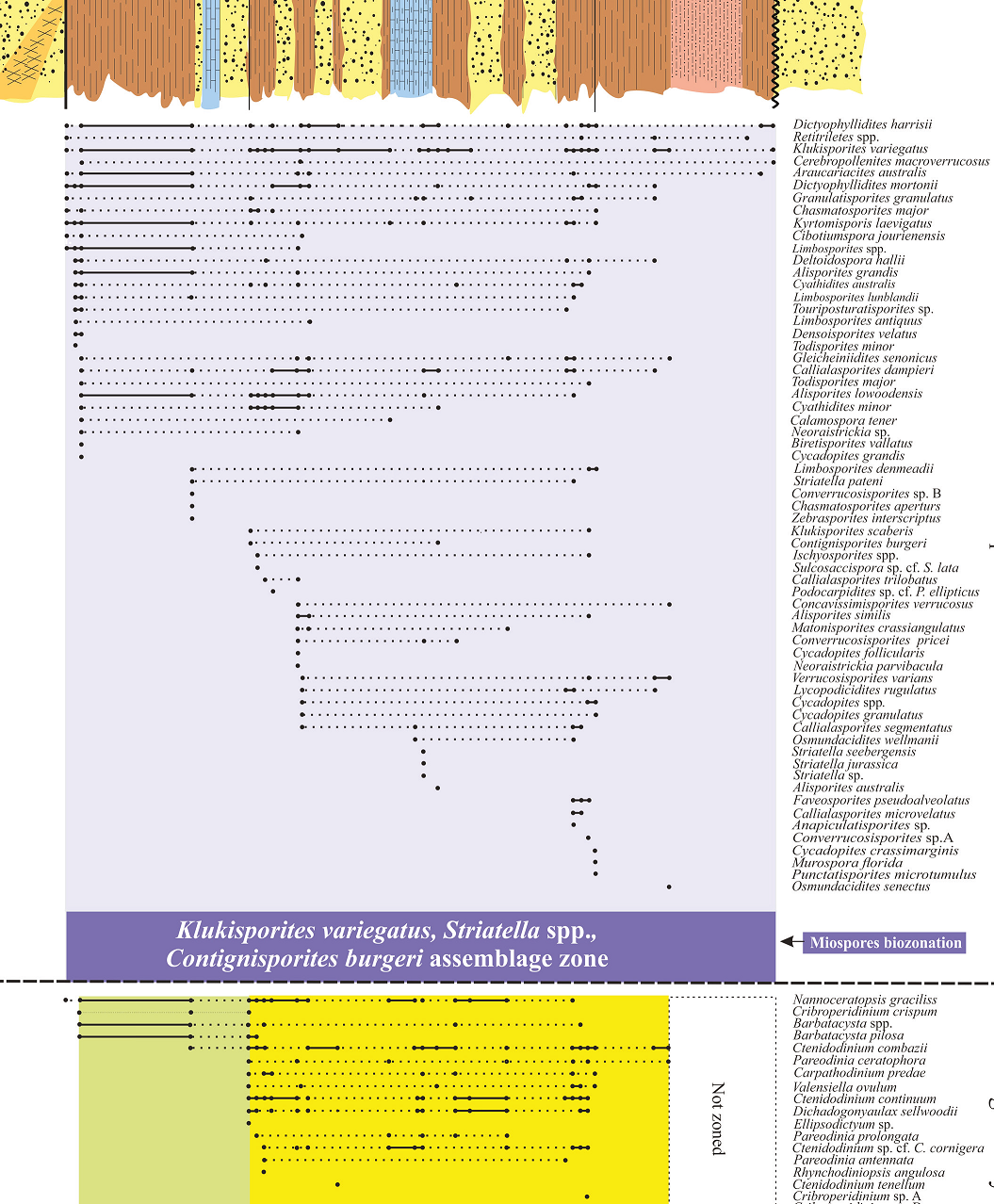
The stratigraphic distribution of certain species enables the introduction of one informal spore-pollen assemblage, and two formal dinoflagellate cyst zones and a subzone (Figure 2). These are based mainly on the first and last appearance datum (FOOs and LOOs) of certain species, respectively. These biozones have potential value for both inter-basinal correlation and relative age determination by reference to known Iranian, Eurasian, and Gondwanan Jurassic palynostratigraphic schemes. Of the taxa encountered in the biozones, only those with known stratigraphic value and/or persistence through all or a certain part of the section are illustrated in Plates I-III.
4.2. PALYNOSTRATIGRAPHY
The Namin palynofloras comprise 44 spore species (30 genera), and 19 pollen species (eight genera). The vertical ranges of certain miospore species with known stratigraphic significance and/ or persistence throughout the section investigated authorize the introduction of one informal miospore-based biozone as Klukisporites variegatus -Striatella spp.- Contignisporites burgeri assemblage zone (Figure 2). The lower and upper stratigraphic boundaries of this biozone extend from the base to the top of the studied section (282 m). Only miospores taxa with known stratigraphic importance and/or persistence are illustrated in Plates I and II. The associated miospores include:
Spores: Anapiculatisporites sp., Biretisporites vallatus, Calamospora tener, Cerebropollenites macroverrucosus, Chasmatosporites apertus, C. major, Cibotiumspora jurienensis, Concavissimisporites verrucosus, Converrucosisporites pricei, Converrucosisporites sp. A, Converrucosisporites sp. B, Cyathidites australis, C. minor, Deltoidospora hallii, Densoisporites velatus, Dictyophyllidites harrisii, D. mortonii, Foveosporites pseudoalveolatus, Gleicheniidites senonicus, Granulatisporites granulatus, Ischyosporites spp., Klukisporites variegatus, K. scaberis, Kyrtomisporis laevigatus, Limbosporites antiquus, L. denmeadii, L. lunblandii, Limbosporites spp., Lycopodiumsporites rugulatus, Matonisporites crassiangulatus, Murospora florida, Neoraistrickia parvibacula, Neorestrickia sp., Osmundacidites senectus, O. wellmanii, Punctatisporites microtumulus, Retitriletes spp., Striatella jurassica, S. patenii, S. seebergensis, Striatella sp., Toripustulatisporites sp., Todisporites major, T. minor, Verrucosisporites varians, Zebrasporites interscriptus.
Pollen: Alisporites australis, A. grandis, A. lowoodensis, A. similis, Araucariacites australis, Callialasporites dampieri, C. microvelatus, C. segmentatus, C. trilobatus, Cycadopites crassimarginis, C. follicularis, C. grandis, Cycadopites spp., Podocarpidites sp. cf. P. ellipticus, Sulcosaccispora sp. cf. S. lata.
The Namin biozonation scheme is compared (Figure 3) with ±coeval palynozones introduced from Gondwana (e.g., Balme, 1957, 1964; Filatoff, 1975; McKellar, 1998; Mantle and Riding, 2012; Ibrahim et al., 2001; Tripathi, 2004; Quattrocchio et al., 2001; Stukins et al., 2013) and Eurasia (e.g., Stefanowicz, 2008; Rostovtseva, 2011; Ashraf, 1977; Ashraf et al., 1999, 2010; Arjang, 1975; Achilles et al., 1984; Bharadwaj and Kumar, 1986; Sajjadi et al., 2007; Dehbozorgi, 2014; Hashemi Yazdi et al., 2014; Sajjadi and Dermanaki Farahani, 2017; Hashemi Yazdi et al. 2018; Badihagh et al., 2019).
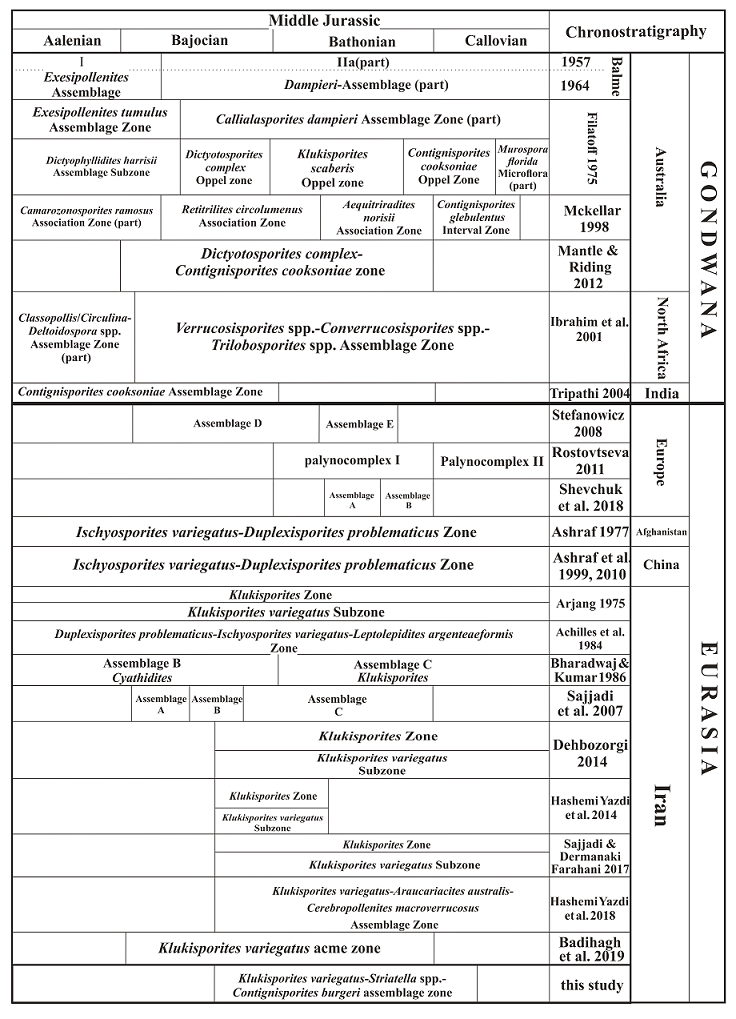
Figure 3 Miospore-based correlation of the Middle Jurassic Shemshak Formation with those established in the Gondwanan and Eurasian coeval strata.
Dinoflagellate cysts are excellent index fossils for the Middle-Late Jurassic because many have a wide geographical distribution and relatively short duration (Riding and Thomas, 1992; Poulsen and Riding, 2003; Riding, 2020; Figure 4). Diverse and profuse dinoflagellate cyst assemblages, in good state of preservation, occur in the Namin stratigraphic section. The assemblages comprise 20 dinoflagellate cysts species (11 genera). Two dinoflagellate cyst zones and a subzone are distinguished in the studied strata (Figure 2) based on the first (FOO) and last (LOO) appearances of the dinoflagellate cysts index species:
Cribroperidinium crispum Total Range Biozone of Woollam and Riding (1983; Bajocian). Cribroperidinium crispum Total Range Biozone spans 71m of the studied interval and is defined by the first observed (FOO) and last observed (LOO) of Cribroperidinium crispum (al. Acanthaulax crispa). The lower and upper stratigraphic limits of the biozone occur 4m and 75m respectively, above the base of the section. Suggested literature ages for this biozone are late Bajocian (Woollam and Riding, 1983) and early-late Bajocian (Riding and Thomas, 1992; Figure 2). Described originally from England (Woollam and Riding, 1983; p. 11) the biozone has subsequently been reported from continental northwest Europe (Poulsen and Riding, 2003) as well as parts of Iran (Ghasemi-Nejad et al., 2012; Mafi et al., 2014; Dehbozorgi, 2014; Hashemi Yazdi, 2015; Sajjadi et al., 2018; Dehbozorgi et al., 2018; Dehbozorgi and Senemari, 2020; Mohammad-khani and Zarei, 2018; Figure 4). Other species recorded from this biozone include: Barbatacycsta spp., B. pilosa, Ctenididinium combazii, and Nannoceratopsis gracilis.
Dichadogonyaulax sellwoodii Interval Biozone of Woollam and Riding (1983; Bathonian-early Callovian).
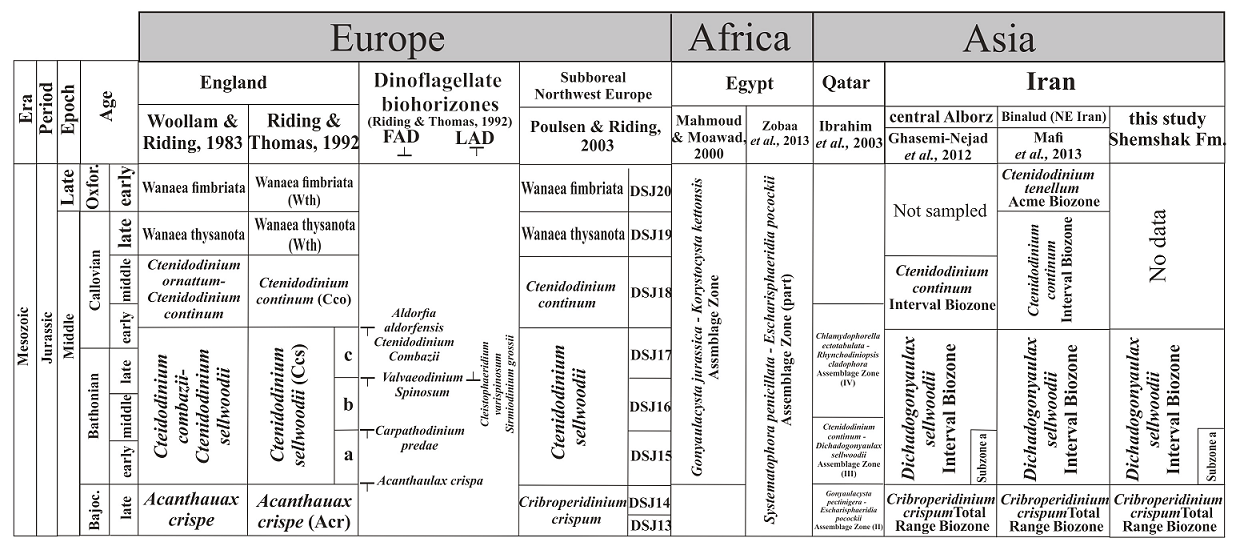
Figure 4 Dinoflagellate-based palynostratigraphic correlation scheme proposed herein for the Middle Jurassic Shemshak Formation with those established in coeval strata.
Succeeding the C. crispum Total Range Biozone, the Dichadogonyaulax (al. Ctenidodinium) sellwoodii Interval Biozone ranges from the LOO of C. crispum (at 75m above the base) to the LOO of Ctenidodinium combazii (241m above the base); thus spanning 166m of the section (Figure 2). The suggested literature age for this biozone is Bathonian- early Callovian, e.g., Riding and Thomas (1992), and Poulsen and Riding (2003).
Described originally from England (Woollam and Riding, 1983; p.11) the biozone has been reported subsequently from sub-boreal northwest Europe (e.g., Poulsen and Riding, 2003), as well as, Egypt (Ibrahim et al., 2001) and parts of Iran (Ghasemi-Nejad et al., 2012; Mafi et al., 2014; Dehbozorgi, 2014; Hashemi Yazdi, 2015; Sajjadi et al., 2018; Dehbozorgi et al., 2018; Dehbozorgi and Senemari, 2020; Mohammad-khani and Zarei, 2018; Figure 4). Dinoflagellate cysts in the D. sellwoodii Biozone are more common and diverse than in the preceding C. crispum Biozone. Except for C. crispum, other species of the older biozone range into the younger biozone. In addition to D. sellwoodii other species first appearing in the younger biozone include: Carpathodinium predae, Cribroperidinium sp. A, Cribroperidinium sp. B, Ctenidodinium sp. cf. C. cornigera, C. continuum, C. tenellum, Ellipsodictyum sp., Pareodinia halosa, P. prolongata, P. ceratophora, P. antennata, Rhynchodiniopsis angulosa and Valensiella ovulum (Figure 2).
2.1. Subzone “a” of Riding and Thomas (1992; early-middle Bathonian).
A subzone recognized within the lower part of the D. sellwoodii Interval Biozone is here designated as Subzone”a” (Figure 2). The respective LOOs of Cribroperidinium crispum and Carpathodinium predae define this subzone. This is compatible with the DSJ15 subzone as part of the northwestern European Ctenidodinium sellwoodii Zone (Poulsen and Riding, 2003), and are both considered of the early-middle Bathonian age (Riding and Thomas, 1992; Figure 4).
5. Age of the palynofloras
Spores and pollen grains provide the sole direct means of determining the age of the Shemshak Formation in the studied section. Dating can be achieved by taking into account the known vertical restriction of certain species in the Middle Jurassic.
Two forms are the most abundant palynomorphs in all the samples studied including K. variegatus and Callialasporites dampieri (Figure 5). In Iran, the former commence from the Middle Jurassic (e.g., Arjang, 1975; Achilles et al., 1984). The relative abundance of K. variegatus and araucarian pollen, notably Callialasporites dampieri, yet the absence of Corollina spp. and Callialasporites turbatus in the Namin section suggests an age no older than the Middle Jurassic (e.g., Helby et al., 1987). An upper age limit is afforded by the absence of Retitriletes watherooensis, Trilobosporites, Pilosisporites, Cicatricosisporites spp. and Ruffordiaspora spp. suggesting that the section is no younger than Late Jurassic (Kimmeridgian; e.g., Helby et al., 1987; McKellar, 1998; Batten and Koppelhus, 1996). Accordingly, the age of Namin section has been confined to the Middle Jurassic.
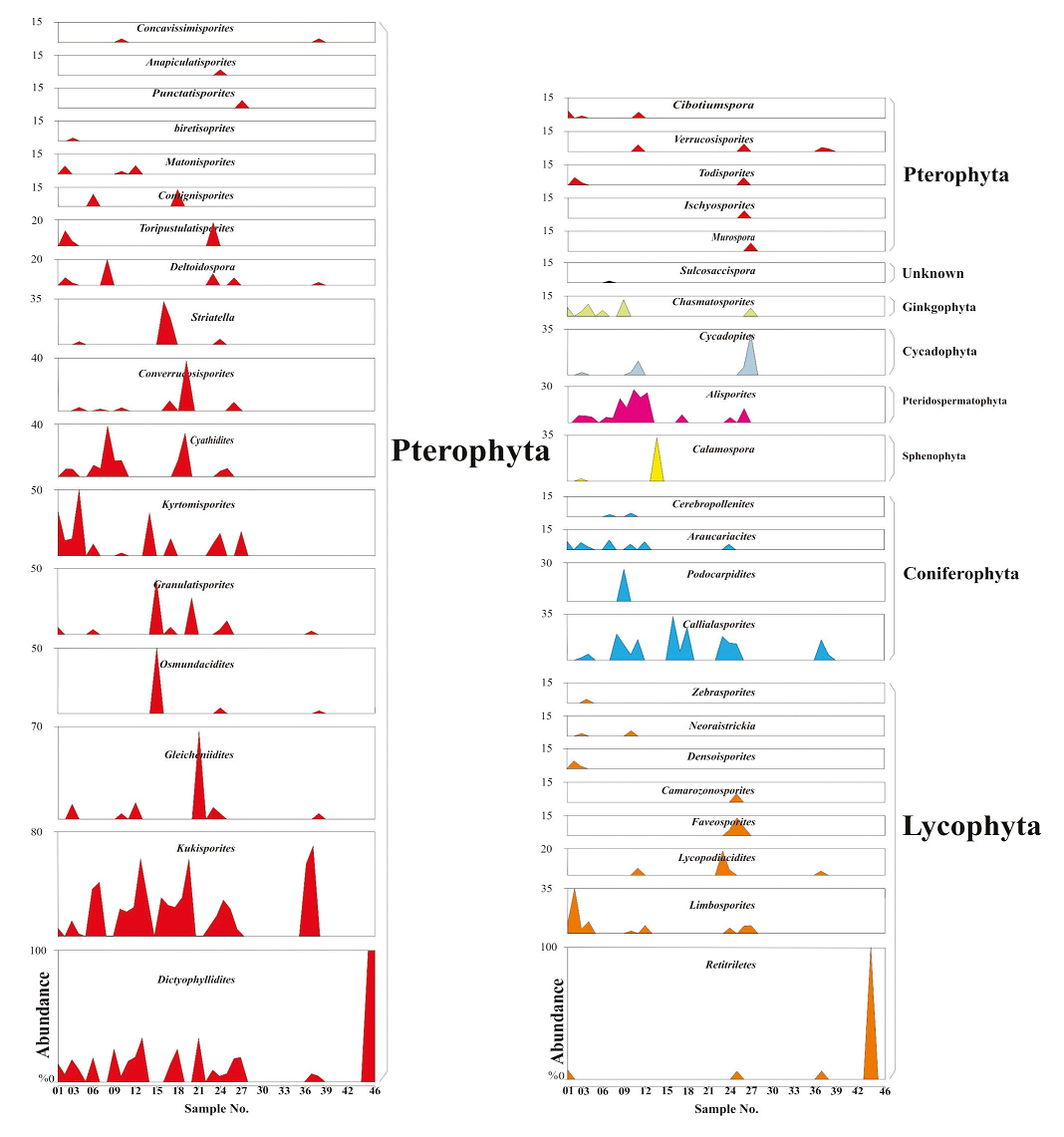
Figure 5 Abundance (percentage) of miospores of the Shemshak Formation at the Namin stratigraphic section.
The first appearance of Contignisporites burgeri in the lower mid part of the section suggests that the strata at that level are no older than Bathonian (Filatoff and Price, 1988, fig. 1). The last appearance datum (LADE) of Striatella patenii (middle Bathonian; Filatoff and Price, 1988), S. scanica (Callovian; Filatoff and Price, 1988; Batten and Koppelhus, 1996), S. seebergensis (early Bathonian; Filatoff and Price, 1988; Batten and Koppelhus, 1996), and S. jurassica (Callovian-? Portlandian; Filatoff and Price, 1988; Batten and Koppelhus, 1996) suggest late Bajocian-Bathonian age for the most part of the Namin section. Furthermore, the co-occurrence of multiple Striatella species, along with Contignisporites burgeri (Filatoff and Price, 1988, fig. 1) supports the age determination.
Importantly the occurrence of several key dinoflagellate cyst index taxa has also greatly helped to restrict the age of the Shemshak Formation palynological assemblages to the late Bajocian-early Callovian. Two identified biozones, namely the Cribroperidinium crispum Total Range Zone (late Bajocian), Dichadogonyaulax sellwoodii Interval Biozone (Bathonian-early Callovian), and Subzone “a” (early-middle Bathonian) contribute to this enhancement. Notably, the presence of the eponymous dinoflagellate cyst further aids in constraining the age of the Shemshak Formation palynological assemblages to the late Bajocian-early Callovian.
6. Paleobiogeographic and paleoecological implications
Pangea’s rifting and fragmentation, which began in the Early Triassic, culminated in the supercontinent’s breakup into the northern Laurasia and southern Gondwana landmasses during the Middle Triassic. In the Permian, the Iranian microcontinent was separated from Gondwana and migrated northward, finally colliding with Eurasia during the Triassic period (Berberian and King, 1981; Alavi et al., 1997; Stampfli and Borel, 2002; Wilmsen et al., 2009a). The Late Triassic collision of the Cimmeride with Eurasia (Corsin and Stampfli, 1977; Stampfli et al., 1991; Muttoni et al., 2001; Moix et al., 2008) caused the beginning of the Cimmerian orogeny and development of peripheral foreland basins that accommodated units such as the Shemshak and Dalichai formations at the southern margin of Eurasia (Fürsich et al., 2009b).
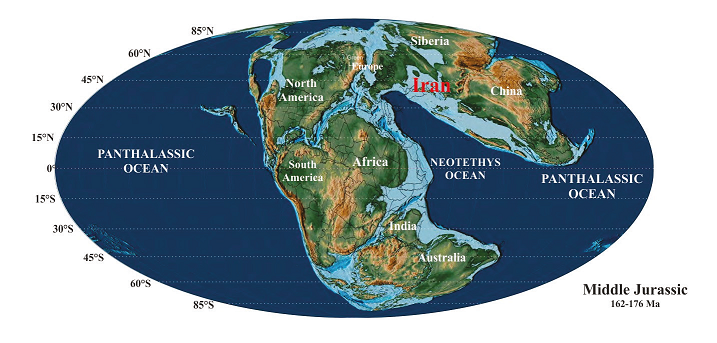
The Iran plate was part of the Eurasian continent during the Jurassic (Wilmsen et al., 2009b; Robert et al., 2014). Studies of Jurassic fossil plant composition and distribution throughout Eurasia reveal two paleoclimatic plant provinces: Siberian in the north and Euro-Sinian in southern Eurasia (Vakhrameev, 1987). The existing paleogeographic maps (Thierry, 2000; Robert et al., 2014) show that in the Middle Jurassic, the north and center of the Iranian plate was located in the northeastern margin of the Neotethys Ocean (Figure 6), in latitudes 20° to 30°N (Seyed-Emami et al., 2008; Wilmsen et al., 2009b).
The worldwide distribution of the Middle Jurassic Araucariaceae-fern-dominated palynofloras confirms the extensive connections within Laurasia and Gondwana, which were clustered around the Tethys Ocean (Mantle and Ridding, 2012). Despite the overall similarities of the Middle Jurassic terrestrial palynoflora of Gondwana and Eurasia, there were some forms largely endemic to the southern hemisphere (Gondwana) that was not found in the Namin section, these include:
Aequitriradites norrisii, Anapiculatisporites dawsonensis, A. pristidentatus, Annulispora densata, Antulsporites saevus, Apiculatisporis taroomensis, Camarozonosporites ramosus, Coronatispora perforata, Convolutispora Prisca, Contignisporites cooksoniae, Dejerseysporites biannuliverrucatus, Dictyotosporites complex, Microcachryidites antarcticus, Neoraistrickia densata, N. elongate, N. equalis, N. rugobacula, Nevesisporites undatus, Perotrilites whitfordensis, Retitriletes circolumenus, R. facetus, R. huttonensis, R. neofacetus, R. nodosus, R. proxiradiatus, Sculptisporis moretonensis, Staplinisporites manifestus, Trilites volkheimeri, Trilobosporites antiquus, Trisaccites microsaccatus, Tuberculatosporites westbournensis, Uvaesporites verrucosus. Consequently, comparing the studied palynofloras with those reported from Gondwana and Eurasia reveals that it is more similar to those from Eurasia.
The tripartite dinoflagellate cyst biozonation (Cribroperidinium crispum, Dichadogonyaulax sellwoodii and Subzone “a” of the D. sellwoodii Zone) identified in the Namin section is very similar to coeval northern Iran (Ghasemi-Nejad et al., 2012; Mafi et al., 2014; Dehbozorgi, 2014; Hashemi Yazdi, 2015; Mohammad-khani and Zarei, 2018; Sajjadi et al., 2018), Northwest European (Woollam and Riding, 1983; Riding and Thomas, 1992; Poulsen and Riding, 2003) and the Russian Platform (Riding et al., 1999). The close similarities of the dinoflagellate cyst assemblages of northern Iran with those of Northwest Europe and the northwestern Tethys during the Middle Jurassic indicate a direct marine connection and faunal exchange between the two areas (Ghasemi-Nejad et al., 2012). The ammonite fauna of the Middle Jurassic in Central Alborz Ranges shows very close relations and similarities to the fauna from other parts of Alborz Ranges, Kopeh-Dagh, East-Central Iran as well as European and Submediterranean regions (Seyed- Emami, et al., 2008).
Table 1 Inferred botanical affinities of miospore taxa identified in this study. Principal sources of information concerning botanical relationships of sporae dispersae: Potonié, 1962, 1967; Couper, 1960; Dettmann, 1963, 1986, 1994; Mädler, 1964; Pocock, 1970; Filatoff, 1975; Filatoff and Price, 1988; de Jersey and Raine, 1990; Vakhrameev, 1991; Dettmann and Clifford, 1992; Boulter and Windle, 1993; Balme, 1995; Batten and Dutta 1997; Hubbard and Boulter 1997; Abbink, 1998; McKellar, 1998; Sajjadi and Playford, 2002; Roghi, 2004; Barrón et al., 2006.
| Miospores | Botanical affinity | ||
| Spores | Anapiculatisporites | Pteridophyta, incertae sedis | Pterophyta |
| Biretisporites | Hymenophyllaceae?, incertae sedis | ||
| Cibotiumspora | Cyatheaceae, Dicksoniaceae | ||
| Concavissimisporites | Cyatheaceae, Schizaeaceae , Dicksoniaceae | ||
| Contignisporites | Pteridaceae | ||
| Converrucosisporites | Dicksoniaceae | ||
| Cyathidites | Cyatheaceae, Dipteridaceae, Dicksoniaceae, Matoniaceae | ||
| Deltoidospora | Cyatheaceae, Dipteridaceae, Dicksoniaceae, Matoniaceae | ||
| Dictyophyllidites | Dipteridaceae, Dicksoniaceae, Cyatheaceae, Matoniaceae | ||
| Gleicheniidites | Gleicheniaceae | ||
| Granulatisporites | Dipteridaceae | ||
| Ischyosporites | Schizaeaceae | ||
| Klukisporites | Schizaeaceae | ||
| Kyrtomisporis | Gleicheniaceae | ||
| Matonisporites | Matoniaceae, Dipteridaceae | ||
| Murospora | Matoniaceae, Dipteridaceae | ||
| Osmundacidites | Osmundaceae, Marattiaceae | ||
| Punctatisporites | Marattiaceae? | ||
| Striatella | Pteridaceae | ||
| Todisporites | Osmundaceae | ||
| Toripustulatisporites | Pterophyta | ||
| Verrucosisporites | Osmundaceae | ||
| Densoisporites | Lycophyta | ||
| Foveosporites | Lycopodium | ||
| Limbosporites | Lycophyta | Lycophyta | |
| Lycopodiacidites | Lycopodiaceae | ||
| Neoraistrickia | Lycopodiaceae, Selginellaceae | ||
| Retitriletes | Lycopodium | ||
| Zebrasporites | Lycophyta | ||
| Calamospora | Sphenophyta | Pteridospermophyta | |
| Pollen grains | Alisporites | Corystospermaceae | |
| Cycadopites | Peltaspermaceae/Ginkgophyta/Cycadophyta | ||
| Araucariacites | Coniferophyta (Araucariaceae) | Coniferophyta | |
| Callialasporites | Araucariaceae) | ||
| Cerebropollenites | Coniferophyta (Taxodiaceae? or Pinaceae?) | ||
| Podocarpidites | (Podocarpaceae) | ||
| Chasmatosporites | Ginkgophyta/Cycadophyta | ||
| Sulcosaccispora | Unknown | ||
Plant macrofossils are useful in paleobotanical and paleoecological studies. By determining the parental plants of miospores, it is possible to provide paleoecological interpretations reflecting a history of the kind of parent vegetation (Traverse, 2007). Inferred botanical affinities of the miospores presented herein (Table 1) have been obtained from various sources (e.g., Potonié, 1962, 1967; Couper, 1960; Dettmann, 1963, 1986, 1994; Mädler, 1964; Pocock, 1970; Filatoff, 1975; Filatoff and Price, 1988; de Jersey and Raine, 1990; Vakhrameev, 1991; Dettmann and Clifford, 1992; Boulter and Windle, 1993; Balme, 1995; Batten and Dutta, 1997; Hubbard and Boulter, 1997; Abbink, 1998; McKellar, 1998; Sajjadi and Playford, 2002; Roghi, 2004; Barrón et al., 2006).
All palynomorphs occurring in three slides from palyniferous samples are counted. The relative abundance of the encountered miospores per sample and their parent plants are shown in Figures 5 and 7. As a result, miospores (53.51%) are the dominant palynomorph component in the Namin section. Their inferred botanical affinities are: Pterophyta (72%), Coniferophyta (8%), Pteridospermophyta (7%), Lycophyta (9%), Ginkgophyta (2%), Cycadophyta (1%) and Sphenophyta (1%); (Figure 7). 0.11% of miospores are unassigned as their affinities are unknown. This inferred botanical composition implies ferns dominated the source vegetation (Figures 5 and 7). From Figure 7 and Table 1, it is apparent that ferns comprise the following plant families: Osmundaceae, Schizaeaceae, Dipteriaceae, Matoniaceae, and Dicksoniaceae. Such a composition implies that the parental vegetation flourished in a warm to warm-temperate, high-humidity climate (Van Konijnenburg-Van Cittert, 2002). Spores of Lycophytic affiliation (both Lycopodium and Selaginellid types) quantitatively constitute the second largest miospores group, although much lower in number than the fern spores. Lycopodium and Selaginella both flourish in moist tropical environments (Sajjadi and Playford, 2002; Tryon and Tryon, 2012). The least common pteridophyte representative in the material examined is Calamospora, an equisetalean-like spore, indicating derivation from arthrophytes. Assuming analogy with modern equivalents, horsetails, the parent plant of these palynomorphs would indicate wet habitats of temperate to subtropical climatic zones (Sajjadi and Playford, 2002; Sajjadi et al., 2015). Dispersed conifer pollen in the Namin sediments is thought to have derived from Podocarpaceae, and Araucariaceae. The Araucariaceae-type pollen in particular consistently dominates the Shemshak Formation palynofloras. Conifers, dominated by Araucariaceae and Podocarpaceae (Table 1) are known to indicate a warm, non-seasonal, possibly coastal environment (Mohr, 1989; Abbink, 1998). The co-occurrence of some index warm to moderate water dinoflagellate cysts (Riding and Hubbard, 1999) such as Pareodinia halosa, Ctenidodinium continuum, and Pareodinia ceratophora confirms this overview.
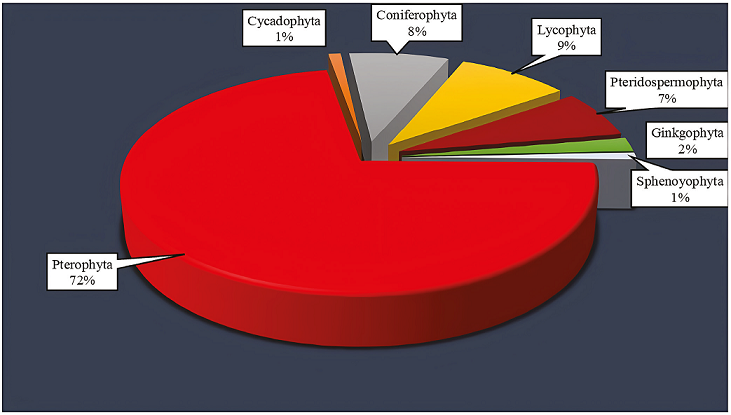
7. Conclusions
The Shemshak palynofloras at the Namin section include in total 83 species of palynomorphs, including 44 spore species (30 genera), 19 pollen species (eight genera), and 20 dinoflagellate cysts species (11 genera). Relative abundances of miospore species vary significantly among samples; however, representatives of K. variegatus and Callialasporites dampieri are particularly abundant. Vertical distribution of certain miospores species enables the introduction of one palynozone, here informally termed the Klukisporites variegatus-Striatella spp.-Contignisporites burgeri assemblage zone.
The presence of multiple Striatella species, as well as, Contignisporites burgeri and Murospora florida collectively indicates a Middle Jurassic (late Bajocian-early Callovian) age. Dinoflagellate cyst species distribution led to the identification of two biozones, which in ascending order are: Cribroperidinium crispum Total Range Zone (late Bajocian), Dichadogonyaulax sellwoodii Interval Zone (Bathonian-early Callovian), and Subzone “a” (early-middle Bathonian). The overall age range of the studied unit, late Bajocian-early Callovian, concurs well with that assessed on the basis of miospores and dinoflagellate cysts.
From a paleofloristic viewpoint, inferred natural relationships of the dispersed spores and pollen imply derivation from a diverse flora, comprising in descending quantitative order, Pterophyta, Coniferophyta, Pteridospermophyta, Lycophyta, Ginkgophyta, Cycadophyta and Sphenophyta. Most miospores are related to ferns indicating that this group was a dominant component of the source vegetation. Comparison with modern plant ecology indicates a moist warm to warm-temperate climate during deposition of the host strata. The presence of some index intermediate to warm water dinoflagellate cysts such as Pareodinia halosa, Ctenidodinium continuum, and Pareodinia ceratophora confirms this overview.
In paleobigeographical terms, most of the miospores and dinoflagellate cysts are shared between the Eurasian and Gondwanan provinces, with a slight preference for Eurasian taxa. This characterization suggests that the Middle Jurassic Shemshak Formation in the western part of the Alborz Ranges was deposited in the southern margin of Eurasia.