











 text new page (beta)
text new page (beta) Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
Permalink
Introducción
El cambio de uso de suelo, el aprovechamiento forestal sin una ordenación adecuada, la tala clandestina, los incendios forestales y las plagas y enfermedades han reducido la cobertura forestal y modificado la composición de los bosques, lo cual ha llevado a que algunas especies, como Pinus chiapensis (Martínez) Andresen, se encuentren bajo alguna categoría de riesgo. Esta conífera crece naturalmente en los estados de Chiapas, Guerrero, Oaxaca, Puebla y Veracruz en México y en el departamento de Huehuetenango de Guatemala (del Castillo y Acosta 2002). P. chiapensis es una de las especies arbóreas más abundantes en las primeras etapas sucesionales del bosque mesófilo de montaña, vegetación húmeda, generalmente libre de heladas, en pendientes pronunciadas y suelos ácidos, con precipitación anual de 1500-2000 mm, temperatura media de 20-22 °C y altitudes de 250 a 2 300 m (del Castillo et al. 2010). P. chiapensis es ampliamente considerada para plantaciones comerciales por su rápido crecimiento y calidad de su madera (Fierros-Mateo et al. 2017). Sin embargo, muchos de sus rodales naturales presentan un alto grado de fragmentación, razón por la cual ha sido clasificada como especie vulnerable por la Unión Internacional para la Conservación de la Naturaleza y catalogada en la categoría de protección especial en la NOM-059 (del Castillo et al. 2010).
Se ha documentado la respuesta del crecimiento en vivero de procedencias de Pinus en efecto de su gradiente altitudinal de origen, por ejemplo, P. pseudostrobus Lindl. se observó crecimiento superior en altura de plantas de familias de baja elevación (26.6 cm) que las de sitios más elevados (18.6 cm) (Antonio-Bautista et al. 2025). Patrones de crecimiento similares fueron reportados para P. chiapensis de poblaciones del sur de México por Dvorak et al. (1996) al observar mejor desempeño en campo de las progenies conforme su elevación de origen era menor, pues las de sitios de baja altitud superaron en 0.5 m a las de procedencias más elevadas, esto a los cinco años de edad. En especies forestales con amplia distribución geográfica, es común observar una alta variación genética entre procedencias, usualmente asociada con gradientes ambientales y aislamiento geográfico (Ahlinder et al. 2021); en México, esto se ha reportado para Pinus pseudostrobus (Escobar-Alonso et al. 2024) y P. patula Schiede ex Schltdl. et Cham. (Sáenz-Romero et al. 2016). Esta variación se manifiesta en una plasticidad fenotípica en características fisiológicas y morfológicas, que se relacionan con la adaptación a condiciones ambientales limitantes (Flores et al. 2018).
La diversidad genética de Pinus chiapensis es importante para su uso productivo y la adaptación a futuros escenarios de cambio climático, así análisis con marcadores moleculares en poblaciones de Oaxaca y Chiapas, México revelan una variabilidad genética significativa entre éstas, pero también señales de cuellos de botella genéticos en poblaciones pequeñas y aisladas (Newton et al. 2002, Syring et al. 2007). Las zonas de menor humedad que rodean a los bosques húmedos en donde crece P. chiapensis junto con las numerosas cordilleras de la región, pueden constituir barreras significativas para el flujo genético entre sus poblaciones (Newton et al. 2002).
Los ensayos de procedencias y progenie permiten evaluar la respuesta de distintos genotipos, tanto en vivero como en campo, desde etapas tempranas de desarrollo (Capilla-Dinorin et al. 2021, Pérez-Luna et al. 2024). El estudio de la variación morfológica durante las primeras etapas del crecimiento de las plántulas es útil al proporcionar información sobre la adaptación inicial de las poblaciones. Características como la longitud y peso de semillas, el número y longitud de hojas cotiledonares, así como la altura y el diámetro de las plántulas, pueden mostrar diferencias entre procedencias (Fuentes-Amaro et al. 2021, Herrera-Hernández et al. 2024). Asimismo, evaluar la diferenciación del crecimiento entre procedencias permite determinar rasgos adaptativos, información que se vuelve más relevante en especies amenazadas en su hábitat de distribución natural (Orquera et al. 2025). Por lo anterior, el objetivo fue determinar la variación existente en las características morfológicas, crecimiento inicial (altura y diámetro) y supervivencia de plántulas de siete poblaciones de Pinus chiapensis, a través de gradientes ambientales en los estados de Puebla y Veracruz.
Materiales y métodos
Recolección de semillas y producción de planta
El germoplasma se recolectó en siete rodales (poblaciones) de P. chiapensis en los estados de Puebla y Veracruz; cada rodal se ubica en una localidad diferente (Tabla 1). Por población se muestrearon entre 15 y 22 árboles, separados a una distancia mínima de 50 m entre individuos para minimizar la endogamia; los árboles fueron seleccionados por su porte superior, altura y diámetro normal dominante, fuste recto, poda natural eficiente y libres de plagas y enfermedades (Capilla-Dinorin et al. 2021). De cada árbol se recolectaron 10 conos, y se colocaron 15 días en condiciones de invernadero a temperatura de 20 a 30 °C (Capilla-Dinorin et al. 2021), para la posterior extracción de las semillas. La germinación se realizó en cajas plásticas de cierre hermético con perlita y agua destilada en una cámara de crecimiento Biotromette Plant Growth Chamber, AB-Line Instruments, Inc., Melrose Park, Illinois, EUA, a 25 °C y 100% de humedad. Doce días después, las plántulas fueron trasplantadas a tubetes individuales (310 ml), identificadas con el árbol progenitor.
Tabla 1 Datos geográficos y climáticos, número de árboles muestreados y densidad de árboles por localidad de las poblaciones de Pinus chiapensis (Martínez) Andresen (Capilla-Dinorin et al. 2021).
| Localidad-Municipio | Lat. N† 19° | Long. O 97° | Altitud (m) | Temp‡ °C | Prec mm | Índice* Aridez | DensŦ | Na |
|---|---|---|---|---|---|---|---|---|
| 1. Tinajas, Atzalan, Ver. | 53'31" | 12'53" | 664 | 21.2 | 2676 | 0.0287 | 139 | 17 |
| 2. Atotocoyan, Yaonáhuac, Pue. | 57'03" | 26'25" | 773 | 20.4 | 3427 | 0.0218 | 96 | 17 |
| 3. Coatectzin, Tlatlauquitepec, Pue. | 57'45" | 26'31" | 778 | 20.4 | 3446 | 0.0217 | 100 | 19 |
| 4. Cruz Alta, Hueytamalco, Pue. | 53'16" | 17'47" | 1262 | 17.7 | 2657 | 0.0254 | 123 | 15 |
| 5. La Pagoda, Hueyapan, Pue. | 53'57" | 24'39" | 1568 | 15.9 | 2287 | 0.0273 | 136 | 22 |
| 6. Aire Libre, Teziutlán, Pue. | 53'03" | 23'48" | 1600 | 15.8 | 2170 | 0.0286 | 143 | 19 |
| 7. Atoluca, Teziutlán, Pue. | 51'47" | 20'47" | 1703 | 15.4 | 1881 | 0.0325 | 44 | 16 |
†Las coordenadas y la elevación determinadas con un geoposicionador en el centro de cada localidad con Datum WGS84, ‡Temperatura media anual, precipitación media anual y DD5 a través de Worldclim, *Índice de Aridez = DD50.5/Prec, más valor menos húmedo; Ŧ Densidad de árboles de P. chiapensis ha-1, Na número de árboles muestreados.
Se empleó una mezcla de 3/4 de composta de corteza de pino y un 1/4 de pumita volcánica (tepezil) y fertilizante Osmocote® como sustrato (15-9-12 N-P-K) de lenta liberación a razón de 6 g L-1. Para evitar el ataque de hongos se aplicó una solución de captan 1 g L-1 una vez por semana por los primeros tres meses. Posteriormente, se aplicó 1g L-1 de Trichoderma harzianum Rifai en el cuarto mes, en intervalos de dos semanas cada aplicación y se redujeron los riegos para evitar la pudrición de la raíz por exceso de humedad. Las plantas fueron mantenidas por 15 meses (hasta mayo de 2018) en un invernadero de plástico calibre 720 con malla sombra de 50% a 1 m sobre la siembra.
Variables evaluadas
Un mes después del trasplante se determinó la altura del hipocótilo, número y longitud de hojas cotiledonares en una muestra de 50 plántulas por procedencia. A partir del segundo mes y hasta los 15 meses se determinó la supervivencia, la altura total y el diámetro basal con regla graduada en 1 mm y vernier digital Truper® calibrado en 0.01 mm, respectivamente. A los 15 meses se cuantificó el número de ramas y se calculó el índice de esbeltez (relación altura/diámetro), que a menor valor indica mayor vigor de la plántula (Aldrete et al. 2024).
Análisis estadístico
El experimento se estableció con un diseño experimental completamente al azar y se evaluaron 50 plantas de cada una de las siete procedencias de P. chiapensis. Las plantas estuvieron bajo competencia completa. El análisis se realizó mediante el procedimiento MIXED de SAS® v9.4 (Littell et al. 2016), con la opción LSMEANS se obtuvieron los valores de las medias ajustadas. El modelo utilizado fue:
Dónde: Yijk es la variable respuesta en la i-ésima población, j-ésimo árbol y k-ésima repetición, µ es el efecto de la media general, Pi es el efecto del i-ésima población, A(P)ij el efecto del árbol anidado en población y eijk es el error experimental. El número de cotiledones resultó normal en la prueba con el procedimiento Univariate del SAS v9.4 (SAS 2022). La supervivencia se transformó con función arcoseno para reducir el efecto de las observaciones extremas (Burbidge et al. 1988). Se realizaron correlaciones de Pearson con los valores de altitud, temperatura y precipitación de cada población. Además, se ajustaron modelos de tiempo de fallo acelerado y de riesgos proporcionales de Weibull para estimar el efecto del gradiente altitudinal sobre la mortalidad (Pérez-Luna et al. 2020):
Dónde: Ln(T) es el logaritmo natural del tiempo de fallo acelerado (tiempo estimado de mortalidad), α es el parámetro de forma, δx es el estimador atribuido al gradiente altitudinal y σε es el parámetro de escala atribuido al error del modelo. Además, se ajustó el modelo de Riesgos Proporcionales de Weibull (RR) (Pérez-Luna et al. 2020) para determinar el riesgo de muerte de las progenies y se calculó la probabilidad de muerte.
Dónde: RR es la razón de riesgo de Weibull, β es el estimador atribuido al rango altitudinal y λ es el parámetro de forma del modelo. Para el ajuste se usaron variables dicotómicas: planta viva = 0 y muerta = 1 y rango altitudinal: bajo = 1 (600 a 1 000 m), medio = 2 (1 001 a 1 500 m) y elevado = 3 (> 1 500 m). Los análisis se realizaron con los procedimientos LIFETEST y PHREG de SAS v9.4 (SAS 2022).
Resultados
Características morfológicas de plántulas
Se observaron diferencias significativas (p ≤ 0.05) entre y dentro de poblaciones en todas las variables morfológicas y de crecimiento evaluadas (Tabla 2). En promedio, el 22.4% de la varianza total se atribuyó a diferencias entre poblaciones, el 27.8% a diferencias entre árboles dentro de poblaciones, y el 49.8% al error residual (Tabla 2).
Tabla 2 Componentes de varianza estimados entre y dentro de poblaciones para las características morfológicas y crecimiento de plántulas en Pinus chiapensis de Puebla y Veracruz.
| Características | Componentes de varianza (%) | Varianza total | ||
|---|---|---|---|---|
| Población | Árbol (población) | Error | ||
| Supervivencia | 6.60** | 93.40** | 0.00 | 109.88 |
| Altura del hipocótilo | 33.52** | 13.91** | 52.57 | 0.71 |
| Número de hojas cotiledonares | 7.95** | 4.17** | 87.88 | 0.57 |
| Longitud de hojas cotiledonares | 31.57** | 12.24** | 56.18 | 0.14 |
| Altura total 15 meses | 33.50** | 29.57** | 36.93 | 262.08 |
| Diámetro a la base 15 meses | 35.11** | 23.16** | 41.72 | 0.63 |
| Índice de esbeltez | 22.43** | 21.15** | 56.42 | 12.65 |
| Número de ramas | 8.64** | 25.06** | 66.30 | 24.58 |
**Significativos con P ≤ 0.05.
La altura del hipocótilo fue de 3.59 cm en promedio con diferencias significativas entre las poblaciones. La menor altura se observó en las plántulas de Tinajas con 2.74 cm y la mayor en Atoluca con 4.16 cm (Tabla 3). El número de hojas cotiledonares presentó poca variación entre poblaciones, con valores promedio que fluctuaron de siete a ocho cotiledones. La localidad de Coatectzin presentó mayor frecuencia de plántulas con siete hojas. En todas las demás poblaciones se encontró el mayor porcentaje de plántulas con ocho hojas cotiledonares. La longitud de las hojas cotiledonares presentó variación entre las poblaciones y el promedio fue de 2.49 cm. Los valores extremos se presentaron en la localidad de Atoluca (2.84 cm) y Atotocoyan (2.21 cm) (Tabla 3).
Tabla 3 Medias (± error estándar) de características morfológicas de plántulas de Pinus chiapensis de Puebla y Veracruz.
| Población (Elevación) | Altura del hipocótilo (cm) | Hojas cotiledonares* | Longitud hoja Longitud hoja cotiledonares (cm) | |||||
|---|---|---|---|---|---|---|---|---|
| Media | 6 | 7 | 8 | 9 | 10 | |||
| Tinajas (664 m) | 2.74 ± 0.10d | 8.23 ± 0.11a | 1 | 15 | 50 | 33 | 2 | 2.34 ± 0.04cd |
| Atotocoyan (773 m) | 3.40 ± 0.17bc | 7.80 ± 0.19ab | 37 | 51 | 12 | 2.21 ± 0.08d | ||
| Coatectzin (778 m) | 3.75 ± 0.11ab | 7.69 ± 0.13b | 51 | 31 | 15 | 3 | 2.39 ± 0.05cd | |
| Cruz Alta (1262 m) | 4.13 ± 0.10a | 7.81 ± 0.09ab | 37 | 50 | 13 | 2.51 ± 0.04b | ||
| La Pagoda (1568 m) | 3.20 ± 0.10c | 7.78 ± 0.09ab | 38 | 48 | 14 | 2.40 ± 0.04c | ||
| Aire Libre (1600 m) | 3.81 ± 0.10ab | 8.02 ± 0.10ab | 21 | 58 | 19 | 2 | 2.73 ± 0.04ab | |
| Atoluca (1703 m) | 4.04 ± 0.10a | 7.83 ± 0.07ab | 38 | 40 | 22 | 2.84 ± 0.03a | ||
Poblaciones ordenadas de menor a mayor elevación. *Media y porcentaje según la frecuencia. Letras iguales dentro de misma columna son estadísticamente iguales (Tukey, P ≤ 0.05).
La supervivencia media fue del 93%, la menor se observó en la población de Atotocoyan (75%). La supervivencia fue mayor que 90% en las demás poblaciones (Tabla 4). El crecimiento de las plantas de P. chiapensis mostró diferencias significativas entre poblaciones (Tabla 4). La altura a los 15 meses osciló entre 24.3 cm (Atotocoyan) y 56.9 cm (Atoluca), con un promedio de 39.9 cm. El diámetro promedio resultó de 3.29 mm. Las plántulas de la población Tinajas fueron las de mayor diámetro con 3.9 mm y las de Atotocoyan las de menor talla con 2.59 mm.
Tabla 4 Medias (± error estándar) de las características morfológicas, el crecimiento inicial y la supervivencia de plántulas de Pinus chiapensis de Puebla y Veracruz.
| Población (elevación) | Supervivencia (%) | Altura (cm) | Diámetro (mm) | Esbeltez (cm/mm) | Número de ramas |
|---|---|---|---|---|---|
| Tinajas (664 m) | 96.0 ± 1.8ab | 47.2 ± 2.6b | 3.90 ± 0.08a | 12.1 ± 0.4b | 6.1 ± 0.7bc |
| Atotocoyan (773 m) | 75.0 ± 3.2d | 25.7 ± 3.7e | 2.59 ± 0.14d | 9.9 ± 0.8c | 8.5 ± 1.2a |
| Coatectzin (778 m) | 94.7 ± 2.1bc | 37.6 ± 3.0cd | 3.32 ± 0.10c | 11.1 ± 0.5bc | 9.2 ± 0.8a |
| Cruz Alta (1262 m) | 96.1 ± 1.8ab | 46.7 ± 2.7b | 3.78 ± 0.08a | 12.2 ± 0.4b | 5.5 ± 0.7bc |
| La Pagoda (1568 m) | 90.9 ± 1.8c | 36.6 ± 2.5d | 2.79 ± 0.08d | 12.7 ± 0.4b | 6.4 ± 0.7b |
| Aire Libre (1600 m) | 96.3 ± 1.8ab | 40.9 ± 2.6c | 3.31 ± 0.08c | 12.5 ± 0.6b | 6.7 ± 0.7ab |
| Atoluca (1703 m) | 98.2 ± 1.8a | 56.5 ± 2.7a | 3.51 ± 0.08b | 15.9 ± 0.4a | 4.5 ± 0.7c |
Poblaciones ordenadas de menor a mayor elevación. Letras iguales dentro de la misma columna son estadísticamente iguales (Tukey, P ≤ 0.05).
El índice de esbeltez varió significativamente entre poblaciones, con valores de 9.8 (Atotocoyan) a 14.5 (Atoluca). Las plantas de Coatectzin presentaron mayor número de ramas (9.2 ± 0.8), incluso, superó en un 50% a Atoluca (Tabla 4). Se encontraron diferencias significativas entre procedencias en el crecimiento acumulado en altura desde los 5 meses de edad. Las plantas de Atoluca, Tinajas, Cruz Alta y Aire Libre presentaron mayor crecimiento que las demás. Esta tendencia fue continua y las diferencias en altura y en diámetro aumentaron a los 15 meses de edad, con mayor proporción en la altura (Figura 1). El crecimiento mensual promedio en altura y diámetro fue de 2.7 cm y 0.22 mm, respectivamente. Las plantas de Tinajas presentaron el valor más alto en crecimiento mensual, y las de Atotocoyan los más bajos, una diferencia de más del 50% en ambas variables.
Análisis de correlación
De las tres variables de plántulas, la longitud de las hojas cotiledonares presentó correlaciones significativas, positiva con la altitud de origen, y negativa con la temperatura y la precipitación (Tabla 5). La supervivencia a los 15 meses de edad mostró una correlación no significativa, aunque positiva con la altitud y negativa con la temperatura y la precipitación. A esta edad, el diámetro basal del tallo no mostró asociación significativa con ninguna variable geográfica.
La altura mostró una correlación moderada no significativa con la altitud y temperatura, y aunque su asociación con relación a la precipitación del sitio de origen fue elevada, no fue significativa (r = 0.70, p < 0.10, Tabla 5). El índice de esbeltez mostró una correlación positiva significativa con la altitud de origen, mientras que la temperatura y la precipitación se correlacionaron negativamente. La mayor correlación se obtuvo entre el índice de esbeltez y el índice de aridez (r = 0.90, p < 0.01). Por otro lado, el número de ramas mostró una correlación positiva con la precipitación, mayor formación de ramas en las plantas juveniles de poblaciones originarias de zonas con mayor pluviosidad, y correlación alta y negativa con el índice de aridez (r = -0.87, p < 0.01).
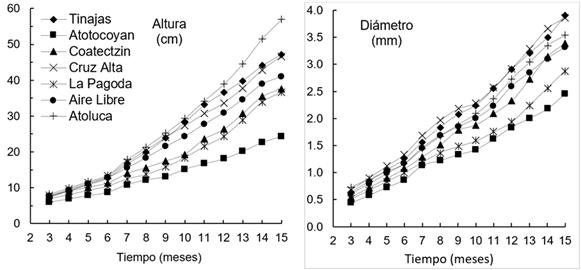
Figura 1 Crecimiento acumulado en altura y diámetro a la base de siete poblaciones de Pinus chiapensis de Puebla y Veracruz a los 15 meses de edad
Tabla 5 Coeficientes de correlación de Pearson para las características morfológicas y de crecimiento de plántulas de Pinus chiapensis con variables ambientales (n = 7).
| Variable | Altitud | Temperatura | Precipitación | Densidad | Índice de aridez |
|---|---|---|---|---|---|
| Altura del hipocótilo | 0.48 | -0.49 | -0.15 | -0.47 | 0.04 |
| Número de hojas cotiledonares | -0.08 | 0.09 | -0.31 | 0.45 | 0.48 |
| Longitud de hojas cotiledonares | 0.80** | -0.79** | -0.81** | -0.35 | 0.76** |
| Supervivencia a 15 meses | 0.40 | -0.38 | -0.60 | 0.05 | 0.62 |
| Altura a 15 meses | 0.43 | -0.40 | -0.70* | -0.30 | 0.80** |
| Diámetro a 15 meses | 0.01 | 0.02 | -0.32 | 0.08 | 0.44 |
| Índice de esbeltez | 0.74* | -0.71* | -0.87** | -0.47 | 0.90*** |
| Número de ramas | -0.63 | 0.61 | 0.87** | 0.21 | -0.87*** |
Significativo con *p < 0.10, **p < 0.05 y ***p < 0.01.
La altura media de las plantas a 15 meses mostró una correlación positiva y significativa con la supervivencia y con la longitud de las hojas cotiledonares (r = 0.74, p = 0.10). El diámetro solo se correlacionó positivamente con la supervivencia (r = 0.86, p = 0.05), y su relación con otras características morfológicas fue positiva, pero no significativa.
Tiempo de fallo acelerado y razón de riesgo de Weibull
El gradiente altitudinal tuvo un efecto significativo (p < 0.05) sobre el tiempo promedio de muerte de las plántulas (Tabla 6). Este tiempo promedio fue de 236, 311 y 409 días para las plántulas originarias de sitios de baja (Tinajas, Atotocoyan y Coatectzin), media (Cruz Alta) y alta elevación (La Pagoda, Aire Libre y Atoluca), respectivamente.
Tabla 6 Parámetros de ajuste del modelo de tiempo de fallo acelerado de Weibull en el crecimiento de plantas de 15 meses de edad de Pinus chiapensis de Puebla y Veracruz.
| Parámetro | Estimador | |z| | Valor de P |
|---|---|---|---|
| Forma (α) | 5.856 | 23.9 | < 0.0001 |
| Efecto atribuido al gradiente altitudinal (β) | 0.275 | 2.01 | 0.04 |
| Escala (σ) | -0.666 | -3.3 | 0.0009 |
El modelo de riesgos proporcionales de Weibull también reveló un efecto significativo (p < 0.05) del gradiente altitudinal sobre la probabilidad de muerte de las plántulas (Tabla 7). Las plántulas provenientes de progenitores de alta elevación presentaron un riesgo de muerte inferior al 50% en comparación con aquellas de baja elevación.
Discusión
Características morfológicas de plántulas
Los resultados de la presente investigación destacan la importancia tanto del origen genético de las procedencias y árboles seleccionados. El número de hojas cotiledonares puede considerarse constante para todas las poblaciones, mas no en su longitud. Los cotiledones constituyen los primeros órganos fotosintetizadores hasta que el hipocótilo forma las hojas primarias. Viveros-Viveros et al. (2005) señalan que el número de cotiledones tiene un alto control genético, posiblemente por ser un carácter preformado en el embrión, que por ende no es afectado por el ambiente.
La supervivencia fue alta para la mayoría de las poblaciones (> 90%). No obstante, la elevada mortalidad presentada en las plantas de Atotocoyan sugiere una posible influencia de la endogamia (Juárez-Agis et al. 2006, Ahlinder et al. 2021). Al respecto, Williams y Savolainen (1996) señalan que, en poblaciones aisladas con fuerte depresión endogámica, los embriones que logran desarrollarse y germinar suelen presentar mutaciones recesivas letales que provocan la muerte de las plántulas en etapas tempranas. Por otra parte, a excepción de Atotocoyan y La Pagoda, el resto de las poblaciones presentaron plantas robustas por el crecimiento del diámetro a la base al superar los 3.0 mm. Por estas características dendrométricas, estas plantas se clasificarían como calidad media (Landis et al. 1990), pero debe considerarse los patrones de crecimiento de P. chiapensis, el cual parece ser mayor en altura a edades tempranas. El diámetro a la base es una característica importante en la evaluación de la calidad de la planta debido a que se encuentra asociada con el vigor, lo que resulta en mayor supervivencia en campo (Aldrete et al. 2024).
Estudios realizados en invernadero con especies de distribución restringida y con poblaciones discontinuas como Pseudotsuga menziesii (Mirb.) Franco muestran crecimiento diferenciado durante las etapas iniciales en función de su procedencia (Juárez-Agis et al. 2006). En algunas especies esta variación se asocia al gradiente altitudinal, con el mayor crecimiento en las de menor altitud (Castellanos-Acuña et al. 2015). No obstante, en este estudio se observó menor crecimiento en las plantas de Atotocoyan localizadas a baja elevación (773 m s.n.m).
En cuanto al índice de esbeltez, los valores obtenidos (> 9.0) en el presente estudio son altos según Aldrete et al. (2024). Sin embargo, no hay reportes de valores adecuados para plantas de rápido crecimiento como lo es P. chiapensis. Es de esperar que, en ambientes fértiles sin restricciones de humedad y temperatura de las localidades naturales de este pino, una altura mayor sea adecuada en virtud de competencia por luz. La ramificación mostrada por las plantas de Coatectzin podría brindarle mayor capacidad para su desarrollo en campo, pues en las etapas iniciales, un árbol con una copa más ramificada puede competir mejor por la luz y el espacio con la vegetación circundante; además, una mayor ramificación puede darle una ventaja para competir y dominar.
Algunos autores han encontrado un patrón clinal de crecimiento en plantas de poblaciones distribuidas en un gradiente altitudinal como Quercus coccifera L., Pinus hartwegii Lindl. o P. devoniana Lindl., donde la altura de la planta aumenta conforme disminuye la altitud de recolección de las poblaciones al tener un periodo de crecimiento más largo (Castellanos-Acuña et al. 2013, Iakovoglou et al. 2020, Sánchez-Mendoza et al. 2023). En otros casos, el clima óptimo de una especie se encuentra en altitudes intermedias de su rango natural de distribución, donde se encuentran las mejores condiciones para expresar su potencial de crecimiento (Barrera-Ramírez et al. 2024). En el presente estudio, la población de mayor elevación de la distribución altitudinal (Atoluca 1703 m) presentó un mayor crecimiento en altura y lo fue hasta los 15 meses de edad, mientras que el mayor diámetro fue la de menor elevación (Tinajas 664 m).
Las diferencias en crecimiento y variables morfológicas pueden indicar una diferenciación geográfica relacionada con el ambiente, o debido a la fragmentación y el aislamiento de los rodales. Igualmente, este comportamiento diferencial puede ser atribuido a la plasticidad fenotípica como respuesta a las condiciones ambientales de cada rodal, situación que se ha observado en otras especies, pues algunas poblaciones son capaces de adaptarse y sobrevivir a condiciones de estrés como sequía, heladas y fauna nociva, tal es el caso de Pinus pseudostrobus (Escobar-Alonso et al. 2024) y P. patula (Pérez-Luna et al. 2024).
Cambios en las condiciones de temperatura, humedad o competencia con otras especies pueden causar que la estructura genética se modifica para aumentar la adaptación a condiciones más adversas, muchas veces se sacrifica el crecimiento para dar lugar a características de mayor valor adaptativo (Ahrens et al. 2024); por ejemplo, resistencia a heladas en poblaciones localizadas en la zona alta o a la sequía en poblaciones ubicadas en la parte baja de la distribución altitudinal (Flores et al. 2018, Barrera-Ramírez et al. 2024). Por otro lado, las plantas que presentan un crecimiento elevado durante sus primeros años pueden captar eficientemente los recursos (agua, luz y nutrientes), lo que les permite mayor ventaja competitiva y resiliencia, en consecuencia, altas probabilidades de supervivencia en campo (Aldrete et al. 2024, Escobar-Alonso et al. 2024).
Correlación entre variables de crecimiento y de origen de las procedencias
La moderada asociación entre la altura de las plantas y la elevación de las poblaciones sugiere un patrón clinal poco definido entre estas variables a edad temprana en P. chiapensis. Estudios previos han encontrado que las procedencias de menor altitud tienden a presentar un periodo de crecimiento más prolongado que las de altitudes mayores, y por tanto más talla (Iakovoglou et al. 2020, Viveros-Viveros et al. 2005). Sin embargo, en P. leiophylla Schiede ex Schltdl. & Cham. y Pinus pseudostrobus Lindl. se ha observado un mejor crecimiento en poblaciones de altitud intermedia (Castellanos-Acuña et al. 2013, Barrera-Ramírez et al. 2024). La disminución del crecimiento en P. hartwegii es más evidente en las poblaciones ubicadas en el límite inferior de su rango altitudinal (Astudillo-Sánchez et al. 2017). En el presente estudio, la población de mayor elevación (Atoluca, 1703 m s.n.m), seguida de la segunda de menor elevación, presentó un crecimiento superior al del resto de las poblaciones evaluadas. No obstante, se detectó cierto grado de asociación clinal de la altura de las plantas con relación a la precipitación del sitio de origen.
También se observó mejor proporción parte aérea/raíz a menor elevación y a mayor temperatura y precipitación del sitio de origen. Por otra parte, se detectó que los sitios más áridos tienden a producir plantas más esbeltas y con menos cantidad de ramas. Reducir el área foliar relativa, menos diámetro y ramas, esto es menos biomasa lateral, puede ser adecuado para disminuir la transpiración en condiciones no tan húmedas.
Estos resultados de correlación evidencian el papel de la procedencia geográfica y climática en la expresión fenotípica temprana, y subrayan la importancia de considerar el origen de la semilla como un factor clave en los programas de producción de planta y reforestación, especialmente bajo escenarios de cambio climático donde la aridez tiende a incrementarse. Esto último puede atribuirse a que una longitud mayor de estas hojas y mayor área fotosintética favorecen el crecimiento a muy temprana edad (Viveros-Viveros et al. 2005). En Pseudotsuga menziesii, la variación en la altura del hipocótilo se ha relacionado con las condiciones ambientales del sitio de origen de las poblaciones (Juárez-Agis et al. 2006); sin embargo, en las poblaciones de P. chiapensis analizadas en este estudio no se observó una relación significativa.
Consideraciones de conservación
Aunque no fue posible determinar con certeza las causas de mortalidad, la mayor tasa de muerte observada en las plántulas provenientes de sitios de menor elevación podría estar relacionada con una menor resistencia al estrés hídrico de estas procedencias. Esta situación pudo haberse agravado debido a las condiciones dentro del invernadero, ya que las temperaturas superiores a 30 °C dificultaron el mantenimiento de una humedad relativa constante y adecuada. Por otra parte, se ha reportado que las progenies de sitios de baja altitud suelen presentar un crecimiento más acelerado, lo que las hace más vulnerables a condiciones climáticas adversas (Castellanos-Acuña et al. 2013, Sánchez-Mendoza et al. 2023). En ese sentido, realizar ensayos para medir la resistencia a factores limitantes como sequía, heladas y calidad de sitio ampliará el conocimiento de los factores que favorecen la supervivencia de las progenies. Aunque no existe evidencia de la aplicación de modelos de riesgos, ni de tiempo de fallo acelerado en ensayos de progenies, en vivero o campo, algunas investigaciones en el ámbito de la propagación vegetativa de coníferas han demostrado su utilidad para determinar factores de riesgo en condiciones controladas (Pérez-Luna et al. 2020). Por ello, los resultados de este estudio sirven como guía para su futura aplicación en trabajos de mejoramiento genético en los cuales se trabaja con poblaciones diferenciadas por factores geográficos como lo son los gradientes altitudinales. El índice de esbeltez elevado podría no ser deseable desde el punto de vista de la calidad de planta. Por el alto valor del índice de esbeltez podría ser necesario aplicar poda apical o control de densidad en vivero para equilibrar la relación altura/diámetro antes de la plantación, particularmente en las localidades de menor precipitación.
Las poblaciones de P. chiapensis en Oaxaca y Chiapas presentan heterocigosidad reducida y sus semillas presentan una baja germinación, factor ligado a la depresión por endogamia, lo que conduce al declive poblacional y mortalidad (del Castillo y Trujillo 2008, del Castillo et al. 2010). La endogamia, resultado de la cruza entre individuos emparentados o de autopolinización, puede reducir la capacidad germinativa, la supervivencia y el crecimiento inicial de las plántulas (Ahlinder et al. 2021). En el caso de las poblaciones de Puebla y Veracruz, a pesar de la alta fragmentación y tamaño reducido de los rodales, y de una producción baja de semillas llena y un porcentaje de semillas vanas alto, se ha reportado una aceptable calidad germinativa y posibilidades de regeneración natural por semilla; en casi todas las poblaciones la germinación resultó mayor a 70 hasta 87%, salvo la población de Atotocoyan con 50%, la más lenta en germinar (Capilla-Dinorin et al. 2021). Además, estos autores indican que la población de Atotocoyan tuvo las semillas más pequeñas y fue la segunda peor en eficiencia de producción de semillas llenas y la segunda mayor con óvulos abortados. Se ha indicado que especies con distribuciones fragmentadas, como P. chiapensis, experimentan una menor depresión por endogamia en las etapas embrionarias tempranas en comparación con especies de poblaciones bien conectadas; sin embargo, en las primeras los efectos de la depresión endogámica tienden a manifestarse en las etapas posteriores (Ahlinder et al. 2021). Por lo que es de esperar que al madurar los árboles de poblaciones como Atotocoyan manifestarán mayores problemas de adaptación, más en el entorno del cambio climático.
El aumento sustancial de la temperatura y la reducción de la precipitación podrían reducir la producción de semillas de la especie y su capacidad de repoblación. Es necesario un manejo adecuado de las poblaciones de P. chiapensis bajo un escenario climático desfavorable, y la renovación de las poblaciones a través de programas de reforestación. La selección cuidadosa de individuos genéticamente adaptados con características adecuadas de crecimiento de brotes y fenología será crucial para minimizar los riesgos de factores adversos climáticos. Sin embargo, son necesarios ensayos de campo en varios sitios para validar la plasticidad fenotípica observada en vivero e identificar genotipos con mayor resiliencia al cambio climático (Escobar-Alonso et al. 2024).
Estas localidades de Puebla y Veracruz, por su lejanía de las poblaciones del sur de México en Guerrero, Oaxaca y Chiapas, adquieren valor estratégico para la conservación, al representar fuentes potenciales de germoplasma ante escenarios de cambio climático (Sáenz-Romero et al. 2016; Capilla-Dinorin et al. 2021), y también como material genético para establecer huertos semilleros y obtener germoplasma de calidad para plantaciones comerciales en regiones tropicales del país u otros donde se ha establecido (Fierros-Mateo et al. 2017). Estas recomendaciones permiten no solo optimizar el establecimiento de plantaciones de este pino, sino también contribuir a su conservación a largo plazo, para reforzar su papel como especie clave en los ecosistemas más septentrionales de las poblaciones de P. chiapensis
Conclusiones
Se encontraron diferencias significativas entre siete poblaciones de Pinus chiapensis de Puebla y Veracruz en características morfológicas y crecimiento de plantas a 15 meses en condiciones de vivero. A través del tiempo, la diferenciación en el crecimiento en altura y diámetro de las plántulas entre orígenes es mayor. El riesgo de muerte de las plántulas incrementó en las progenies de menor elevación. La mayor precipitación del sitio de origen se asoció a más ramas, menor longitud de hojas cotiledonares, altura a 15 meses y relación altura/diámetro, lo que indica selección por este factor. Es de importancia considerar la procedencia al planificar programas de restauración o reforestación con esta especie, para mantener alta diversidad genética dentro de las plantaciones.

