











 text in
text in  English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
Permalink
Introducción
Los bosques tropicales amazónicos son sumideros cruciales de carbono que contribuyen a mitigar el cambio climático (Malhi & Grace, 2000; Phillips et al., 1998). Dentro de estos ecosistemas, el Hotspot Andino-Amazónico destaca por su biodiversidad y su papel en los ciclos de carbono. Ecuador es uno de los 17 países megadiversos (Bravo, 2014), alberga aproximadamente 17 000 especies de plantas superiores, incluidas 4 000 endémicas (Neill, 2012). Sin embargo, la expansión agrícola y otras actividades humanas han reducido drásticamente la cobertura forestal en la región (Balslev et al., 2015).
La familia Arecaceae (palmas) tiene un rol fundamental en estos ecosistemas debido a su abundancia, diversidad y capacidad para almacenar carbono (Kristiansen et al., 2009). Especies como Iriartea deltoidea Ruiz & Pav. y Oenocarpus bataua Mart. destacan por su gran cantidad de biomasa que llegan a acumular en zonas de gradiente altitudinal (Balslev et al., 2017; Goodman et al., 2013). Sin embargo, su contribución ha sido subestimada por algunas de sus características morfológicas particulares (Goodman et al., 2013).
La diversidad de palmas varía con la altitud, tipo de suelo y precipitación (Alvez-Valles et al., 2018). Esto sugiere que los gradientes altitudinales, presentes en el Hotspot Andino-Amazónico, tienen una influencia determinante en la estructura de las comunidades de palmas, ya que condicionan su distribución y composición (Balslev et al., 2015).
Las especies de palmas son cruciales no solo para la mitigación del cambio climático (Miranda et al., 2025), sino también para las comunidades locales, al proveer recursos esenciales como alimentos y diversos materiales (Zambrano et al., 2021). Dado su valor ecológico y económico, es imprescindible estudiar cómo los gradientes altitudinales afectan su estructura y biomasa (Torres et al., 2020). Los objetivos del presente estudio buscan: (A) Evaluar la diversidad y riqueza de la familia Arecaceae en un gradiente altitudinal del bosque siempreverde piemontano que forma parte del Hotspot Andino-Amazónico; y (B) Cuantificar la reserva de carbono en la biomasa aérea y en el suelo.
Materiales y Métodos
Área de estudio
El estudio se llevó a cabo en los bosques andino-amazónicos de la provincia de Napo, Ecuador en colaboración con el Centro Experimental de Investigación y Producción Amazónica (CEIPA), ubicado en la provincia de Pastaza, Cantón Carlos Julio Arosemena Tola, en el kilómetro 44 de la vía Puyo-Tena (Figura 1).
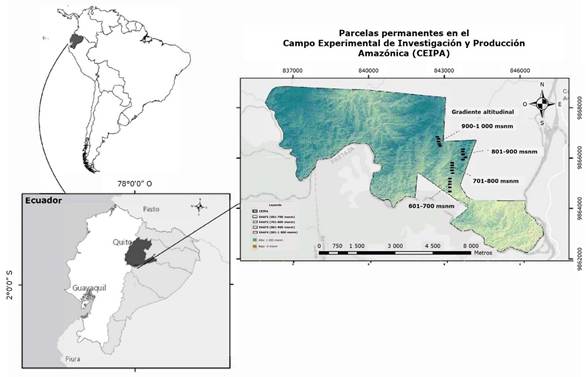
El área de estudio se localiza en una región montañosa dominada por crestas altas y colinas compuestas de rocas volcánicas y sedimentarias de origen reciente (Galeas & Guevara, 2012). Las condiciones bioclimáticas predominantes varían entre húmedo pluvial e hiperhúmedo (Galeas & Guevara, 2012). La altitud del área varía desde 580 hasta 1 120 m; la temperatura media anual es de 23 °C, precipitación anual de 4 119 mm y una humedad relativa de 77 % (Galeas & Guevara, 2012).
Para el establecimiento de las 20 parcelas permanentes (PP), se realizó un reconocimiento preliminar del área mediante ortofotos obtenidas de la base de datos del Instituto Geográfico Militar (IGM) del Ecuador, imágenes satelitales proporcionadas por la plataforma Copernicus Sentinel-2, y bases cartográficas del Sistema de Información Geográfica del Ministerio del Ambiente y Agua del Ecuador (Galeas & Guevara, 2012). Las parcelas se establecieron en cuatro altitudes del Bosque Andino-Amazónico Perennifolio (EAAF), con pendientes que variaron entre 3 y 12 % y el ángulo de pendiente fue de 3 a 30 % (Cuadro 1).
Cuadro 1 Número de palmas y especies en cuatro gradientes altitudinales a pequeña escala.
| Código | Altura (msnm) |
Área de muestreo (m) |
Coordenadas | Número de especies |
Individuos ha-1 |
|---|---|---|---|---|---|
| EAAF1 | 601-700 | 100×10 | S 1.223822° O 77.917784° |
6 | 275 |
| EAAF2 | 701-800 | 100×10 | S 1.212358° O 77.916004° |
5 | 297 |
| EAAF3 | 801-900 | 100×10 | S 1.211196° O 77.912133° |
8 | 364 |
| EAAF4 | 901-1 000 | 100×10 | S
1.206227° O 77.920118° |
8 | 980 |
EAAF = Bosque Andino-Amazónico Perennifolio.
Protocolo de muestreo
Se instalaron transectos permanentes, generalmente utilizados en el muestreo exploratorio rápido, denominados muestreo tipo Gentry (Gentry, 1982; Montufar & Pintaud, 2006), modificando las parcelas originales de 500×2 m a parcelas de 100×10 m (Figura 2), es decir de 0.1 ha-1 (García-Cox et al., 2023). De esta manera, en cada parcela se registró el número de individuos de las especies de la familia Arecaceae, los cuales se marcaron, colectaron, herborizaron y depositaron en el Herbario ECUAMZ-Ecuador Amazónico. La identificación taxonómica se realizó con el apoyo de claves, catálogos ilustrados y recursos digitales, como Tropicos (Bravo, 2014) y The Plant List (Neill, 2012), para asegurar la precisión en la nomenclatura (Cazzolla et al., 2022; Gentry, 1982; Ter Steege, 1998).
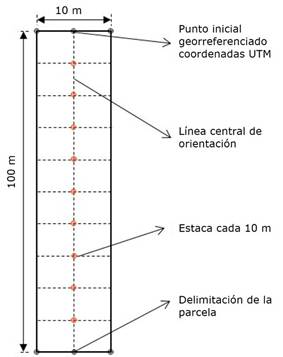
Figura 2 Esquema de los transectos permanentes implementados para el inventario, cálculos de diversidad y carbono en el gradiente.
Se midieron altura (m) y el diámetro a la altura del pecho, es decir, a 1.3 m del suelo (DAP, cm), de los individuos, mediante un hipsómetro Haglöf ® Vertex IV y un calibrador Haglöf ® Mantax Blue. Se hicieron mediciones diferenciadas para aquellos individuos con un DAP≥10 cm y para los menores de ese tamaño.
Determinación de la composición florística y carbono
Se estimó la riqueza de especies (S), Índice de diversidad de Shannon-Wiener (H’), el Índice de Simpson (λ) e índices de equidad de Pielou (J’) y de dominancia de Simpson (Eλ) (Cuadro 2) (Jost & González-Oreja, 2012). El peso ecológico de las especies se determinó mediante el Índice de Valor de Importancia Ecológica (Medrano et al., 2017).
Cuadro 2 Parámetros ecológicos e Índice de Valor de Importancia evaluados.
| Parámetros | Ecuaciones | Referencias | |
|---|---|---|---|
|
Índice de
Shannon- Wiener (H’) |
|
García-Cox et al. (2023) | (1) |
| Índice de Margalef (D) |
|
García-Cox et al. (2023) | (2) |
| Índice de Simpson (𝛌) |
|
Aryal et al. (2019) | (3) |
| Índice de Pielou (J’) |
|
Medrano et al. (2017) | (4) |
|
Abundancia relativa
(Ar) |
|
García-Cox et al. (2023) | (5) |
|
Dominancia relativa
(Dr) |
|
García-Cox et al. (2023) | (6) |
| Área basal (G) |
|
García-Cox et al. (2023) | (7) |
|
Frecuencia relativa
(Fr) |
|
García-Cox et al. (2023) | (8) |
|
Índice
de Valor de
Importancia (IVI) |
|
García-Cox et al. (2023) | (9) |
pi = Proporción de individuos de la especie
Muestreo de suelos
En cada parcela se recolectaron cinco submuestras de suelo a dos profundidades (0-10 y 10-30 cm), que posteriormente se homogeneizaron para obtener una muestra de suelo representativa para cada profundidad de suelo (Bravo-Medina et al., 2021). Para este estudio, el carbono total almacenado (CTA) resultó de la suma de los siguientes componentes: carbono aéreo (CA), carbono subterráneo de la raíz (CSR), carbono de la biomasa de la hojarasca (CBH) y carbono orgánico del suelo (COS) (López-Santiago et al., 2019). Sin embargo, esa Ecuación (10) puede estar formada por más o menos componentes según el criterio y el interés de los investigadores (Dantas et al., 2021; Pradhan et al., 2012).
Donde:
CTA = Carbono total almacenado en Mg C ha-1; y el Mg CO2 ha-1 es resultado del producto entre el Mg C ha−1 y 3.67 que corresponde al peso molecular del CO2
Para estimar el carbono almacenado en las palmas a lo largo del gradiente altitudinal, se utilizó la ecuación alométrica propuesta por Chave et al. (2005), adaptada para incluir todas las palmas con un diámetro a la altura del pecho DAP≥10 cm. La Ecuación 11 se basa en la densidad de la madera (ρ, en g cm3) y el DAP (en cm), y se expresa como sigue:
Donde:
BSS = Biomasa sobre el suelo por palma en Megagramos (Mg)
ρ = Peso específico de las especies en g cm3
DAP = Diámetro a la altura del pecho (cm) para individuos con DAP≥10 cm
Los valores de biomasa sobre el suelo (BSS) se extrapolaron por hectárea para estimar la biomasa aérea total en Megagramos por hectárea. Para especies sin datos específicos (Chave et al., 2009) de densidad de la madera, se utilizó el promedio de 0.632 g cm3 de la base de datos de Montufar y Pintaud (2006).
En este estudio se llevó a cabo una revisión exhaustiva de la literatura para comprender los usos tradicionales y el valor socioeconómico de las palmas amazónicas (Borchsenius & Moraes, 2006). Proceso que se incorporó con el objetivo de proporcionar un contexto más completo sobre la importancia de las palmas, se abarcó tanto perspectivas ecológicas como socioeconómicas (De la Torre et al., 2008). Esta aproximación integradora permitió elaborar un panorama amplio de como las palmas contribuyen al bienestar de las comunidades locales como a la conservación global (Miranda et al., 2025); con ello se asegura que la discusión sobre su conservación contemple aspectos biológicos y socioculturales.
Análisis estadístico
El análisis de los datos incluyó dos enfoques principales: (1) Caracterización de la composición florística a lo largo de un gradiente altitudinal, evaluando la diversidad mediante el Índice de Shannon-Wiener y el número efectivo de especies (Alvez-Valles et al., 2018; Balslev et al., 2015); y (2) Estimación del aporte de las especies de Arecaceae para almacenar carbono. Se verificaron los supuestos de normalidad y homocedasticidad con las pruebas de Kolmogorov-Smirnov y Bartlett (Berlanga & Rubio, 2012) (p<0.001), lo que permitió aplicar análisis de varianza (ANOVA). Para las variables que no cumplían con los supuestos de normalidad, se empleó la prueba no paramétrica de Kruskal-Wallis como alternativa (Berlanga & Rubio, 2012).
El ANOVA comparó los índices de diversidad y densidad de palmas a lo largo del gradiente, utilizando letras superíndices para diferencias significativas entre categorías. Los análisis se realizaron en RStudio (versión 3.6.0) (de Lima et al., 2023), mediante los paquetes BiodiversityR versión 2.16-1 y Vegan versión 2.6-10 para el calcular y analizar los índices de diversidad (Fernandez, 2019).
Resultados y Discusión
Diversidad y riqueza en el gradiente altitudinal
Se registraron 1 916 individuos, distribuidos en 11 especies de palmas. La mayor diversidad se registró entre los 801 y los 1 000 msnm, mientras que entre 701 y 800 msnm se obtuvieron los menores valores (Cuadro 3). Este patrón sugiere que condiciones climáticas de humedad y temperaturas moderadas favorecen la diversidad, resultados que concuerdan con otros estudios previos sobre el particular (Alvez-Valles et al., 2018; Balslev et al., 2017).
Cuadro 3 Promedios±desviación estándar, índices de Riqueza, Shannon, Simpson, Margalef, Pielou y Densidad de la familia Arecaceae.
| Índices | Todos | 601-700 msnm |
701-800 msnm |
801-900 msnm |
901-1 000 msnm |
Valor p † |
|---|---|---|---|---|---|---|
| Riqueza | 4.65 ±1.03 |
3.80a ±0.45 |
3.80a ±0.45 |
5.40b ±0.54 |
5.60b ±0.89 |
*** |
| Shannon | 1.03 ±0.29 |
0.91a ±0.21 |
0.82a ±0.24 |
1.00a ±0.23 |
1.38b ±0.15 |
** |
| Simpson | 0.51 ±0.15 |
0.48a ±0.12 |
0.43a ±0.14 |
0.47a ±0.13 |
0.68b ±0.04 |
** |
| Margalef | 1.33 ±0.38 |
1.03a ±0.16 |
1.03a ±0.20 |
1.56b ±0.20 |
1.69b ±0.36 |
*** |
| Pielou | 0.67 ±0.14 |
0.68a ±0.11 |
0.61a ±0.15 |
0.60a ±0.14 |
0.80b ±0.03 |
* |
| Densidad
(palmas ha-1) |
95.80
±73.20 |
55.00a ±36.84 |
59.40a ±48.68 |
72.80a ±33.43 |
196.00b ±60.49 |
*** |
†ANOVA. Valor p: * = p<0.05; ** = p<0.01; *** = p<0.001; n. s. = No significativo entre grupos. Las letras diferentes representan diferencias significativas entre las medias de las diferentes altitudes (p<0.05).
En altitudes de 801 a 900 y 901 a 1 000 msnm, los índices de Margalef y Shannon alcanzaron sus valores más altos, que indican comunidades diversas y equilibradas (Alvez-Valles et al., 2018). El Índice de Pielou reflejó una alta equidad en estas zonas, lo cual sugiere un ecosistema equilibrado y resiliente, en el que las especies están uniformemente distribuidas y desempeñan roles importantes en la funcionalidad del bosque (Pintaud et al., 2008).
La densidad de palmas fue mayor entre los 901 y los 1 000 msnm (196 palmas ha-1). Destaca su capacidad para prosperar en condiciones ambientales estables y su papel crucial en el almacenamiento de carbono (Torres et al., 2020). Especies dominantes como Iriartea deltoidea y Oenocarpus bataua contribuyen significativamente al secuestro de carbono y a la sostenibilidad ecológica del bosque amazónico (Goodman et al., 2013). Estos hallazgos subrayan la importancia de proteger áreas críticas en altitudes superiores para conservar la biodiversidad y optimizar el almacenamiento de carbono en la Amazonía Ecuatoriana (Balslev et al., 2017; Malhi & Grace, 2000).
Abundancia y Valor de Importancia ecológica (IVI)
El análisis del Índice de Valor de Importancia (IVI) destaca a Iriartea deltoidea como especie dominante a lo largo del gradiente altitudinal estudiado, con altos valores en abundancia, frecuencia y dominancia relativas, con valores de 39.25 entre los 600 a 700 msnm y 56.03 de los 801 a 900 msnm (Cuadro 4). Ese patrón refleja la capacidad de adaptarse a diversas condiciones microambientales y su importancia en la estructura de los bosques tropicales húmedos (Balslev et al., 2017). En altitudes que varían de 901 a 1 000 m destaca Oenocarpus bataua, con un valor de IVI de 30.14, debido a su alta abundancia relativa (43.06 %) y su contribución significativa a la biomasa forestal (Goodman et al., 2013).
Cuadro 4 Índice de Valor de Importancia de especies de la familia Arecaceae.
| Especie | AR (%) | FR (%) | DR (%) | IVI |
|---|---|---|---|---|
| EAAF1 (601-700 msnm) | ||||
| Iriartea deltoidea Ruiz & Pav. | 21.45 | 69.98 | 26.32 | 39.25 |
| Wettinia maynensis Spruce | 51.27 | 5.38 | 15.79 | 24.15 |
| Oenocarpus bataua Mart. | 10.55 | 21.71 | 21.05 | 17.77 |
| Geonoma macrostachys Mart. | 15.64 | 0.00 | 26.32 | 13.98 |
| Ceroxylon amazonicum Galeano | 0.73 | 2.21 | 5.26 | 2.73 |
| Astrocaryum murumuru Mart. | 0.36 | 0.72 | 5.26 | 2.12 |
| EAAF2 (701-800 msnm) | ||||
| Iriartea deltoidea Ruiz & Pav. | 45.12 | 93.87 | 26.32 | 55.10 |
| Wettinia maynensis Spruce | 39.39 | 6.13 | 21.05 | 22.19 |
| Geonoma macrostachys Mart. | 11.45 | 0.00 | 26.32 | 12.59 |
| Oenocarpus bataua Mart. | 3.70 | 0.00 | 21.05 | 8.25 |
| Phytelephas tenuicaulis (Barfod) A. J. Hend. | 0.34 | 0.00 | 5.26 | 1.87 |
| EAAF3 (801-900 msnm) | ||||
| Iriartea deltoidea Ruiz & Pav. | 56.87 | 92.71 | 18.52 | 56.03 |
| Oenocarpus bataua Mart. | 18.68 | 1.20 | 18.52 | 12.80 |
| Geonoma macrostachys Mart. | 15.66 | 0.00 | 18.52 | 11.39 |
| Phytelephas tenuicaulis (Barfod) A. J. Hend. | 5.77 | 0.00 | 14.81 | 6.86 |
| Wettinia maynensis Spruce | 1.37 | 0.00 | 11.11 | 4.16 |
| Socratea exorrhiza (Mart.) H. Wendl. | 0.82 | 3.86 | 7.41 | 4.03 |
| Euterpe precatoria Mart. | 0.55 | 2.23 | 7.41 | 3.39 |
| Aiphanes ulei (Dammer) Burret | 0.27 | 0.00 | 3.70 | 1.33 |
| EAAF4 (901-1 000 msnm) | ||||
| Iriartea deltoidea Ruiz & Pav. | 38.98 | 61.94 | 17.86 | 39.59 |
| Oenocarpus bataua Mart. | 43.06 | 29.52 | 17.86 | 30.14 |
| Geonoma macrostachys Mart. | 9.59 | 0.00 | 17.86 | 9.15 |
| Euterpe precatoria Mart. | 1.12 | 7.03 | 14.29 | 7.48 |
| Aiphanes ulei (Dammer) Burret | 1.12 | 0.00 | 14.29 | 5.14 |
| Wettinia maynensis Spruce | 5.61 | 1.51 | 7.14 | 4.76 |
| Socratea exorrhiza (Mart.) H. Wendl. | 0.31 | 0.00 | 7.14 | 2.48 |
| Bactris gasipaes Kunth | 0.20 | 0.00 | 3.57 | 1.26 |
AR = Abundancia relativa; FR = Frecuencia relativa; DR = Densidad relativa; IVI = Índice de Valor de Importancia; EAAF = Bosque Andino-Amazónico Perennifolio.
Especies como Geonoma macrostachys Mart. y Wettinia maynensis Spruce mostraron una mayor variabilidad en su contribución, fueron más abundantes en altitudes intermedias, pero con menor frecuencia relativa en comparación con Iriartea deltoidea. Esta distribución más dispersa, refuerza la resiliencia del ecosistema mediante una alta diversidad funcional (Alvez-Valles et al., 2018). Los resultados enfatizan el papel esencial de las palmas en el ciclo del carbono y la importancia de las políticas de conservación para preservar las especies dominantes y su capacidad de almacenamiento en los ecosistemas tropicales (Balslev et al., 2017; Keeling & Phillips, 2007; Montufar & Pintaud, 2006).
Reservas de carbono en el gradiente altitudinal
El carbono en palmas es mayor en el piso de 601-700 msnm (11.20 Mg ha-1, p<0.05), disminuye en altitudes intermedias de 701-800 msnm (6.26 Mg ha-1) y en cotas superiores es aún más bajo: 801-900 msnm (7.80 Mg ha-1) y de 901 a 1 000 msnm (3.11 Mg ha-1) (Cuadro 5). Estos resultados destacan la relevancia de las zonas bajas en el secuestro de carbono y refuerzan el papel clave de las palmas en los ciclos de carbono en los bosques estudiados (Balslev et al., 2015; Dauber et al., 2000).
Cuadro 5 Promedios±desviación estándar de las variables estimadas.
| Variables (Mg ha-1) |
Todos | (601-700 msnm) |
(701-800 msnm) |
(801-900 msnm) |
(901-1 000 msnm) |
Valor p † |
|---|---|---|---|---|---|---|
| Carbono palmas | 7.10 ±4.98 |
11.20b ±6.14 |
6.26a,b ±5.09 |
7.80a,b ±3.16 |
3.11a ±1.36 |
* |
| CO 2eq_palmas | 26.03 ±18.27 |
41.14b ±22.52 |
22.97a,b ±18.65 |
28.58a,b ±11.58 |
11.42a ±4.97 |
* |
| Carbono suelo | 29.69 ±7.33 |
25.82 ±1.76 |
31.02 ±10.52 |
31.63 ±10.48 |
30.29 ±2.43 |
n. s. |
| CO 2eq_suelo | 108.96 ±26.92 |
94.75 ±6.45 |
113.83 ±38.62 |
116.08 ±38.46 |
111.17 ±8.92 |
n. s. |
| Carbono total | 36.79 ±9.02 |
37.04 ±6.30 |
37.28 ±15.27 |
39.43 ±9.04 |
33.40 ±2.96 |
n. s. |
| CO 2eq_total | 134.98
±33.10 |
135.89
±23.10 |
136.80 ±56.04 |
146.67
±33.17 |
122.58
±10.86 |
n. s. |
Carbono palmas = Carbono en palmas; CO 2eq_palmas = Dióxido de carbono equivalente en palmas; Carbono suelo = Carbono en suelo; CO 2eq_suelo = Dióxido de carbono equivalente en suelo; Carbono total = Carbono total; CO 2eq_total = Dióxido de carbono equivalente total. †ANOVA. Valor p: * = p<0.05; n. s. = No significativo entre los grupos. Las letras diferentes representan diferencias significativas entre las medias de las distintas altitudes (p<0.05).
Iriartea deltoidea y Oenocarpus bataua representan 65 % de las palmas registradas, sobresalen por su alta biomasa y contribución al almacenamiento de carbono, ya que acumulan entre 3.11 y 11.20 Mg ha-1 de carbono aéreo a lo largo del gradiente altitudinal evaluado (Figura 3; Cuadro 4), lo que refleja su predominancia en las zonas bajas y medias. Estas observaciones son consistentes con estudios previos en la Amazonía Ecuatoriana (García-Quintana et al., 2021), en los que se ha demostrado que las palmas, a pesar de ser menos estudiadas que las especies maderables, contribuyen sustancialmente al almacenamiento de carbono en los bosques tropicales (Torres et al., 2020).
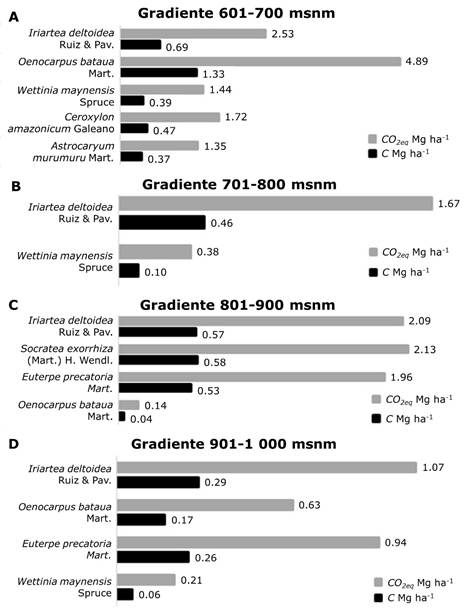
Gradiente (msnm): A = 601 a 700; B = 701 a 800; C = 801 a 900; D = 901 a 1 000.
Figura 3 Contribución de las especies de palmas al reservorio de carbono en un gradiente altitudinal.
El carbono almacenado en el suelo mostró mayor estabilidad a lo largo del gradiente (25.82-31.63 Mg ha-1, p>0.05), lo que equilibró las reservas totales del ecosistema. El carbono total (suma de carbono en palmas y suelo) presentó una variación limitada (33.40-39.43 Mg ha-1). Esto indica que, aunque las palmas contribuyen significativamente al carbono aéreo, el suelo desempeña un rol clave en el balance global de carbono en los bosques estudiados (Bravo-Medina et al., 2021).
CO 2eq almacenado en las especies de palmas
El CO2 equivalente alcanzó un máximo de 41.14 Mg ha-1 en los pisos bajos, y disminuyó a 22.97 y 28.58 Mg ha-1 en las altitudes intermedias. Las especies como Oenocarpus bataua e Iriartea deltoidea destacan por su capacidad de secuestro de carbono, esencial para las estrategias de conservación y mitigación del cambio climático. Entre las alturas de 601 a 700 msnm, Oenocarpus bataua almacena 4.89 Mg ha-1 de CO 2eq y 1.33 Mg ha-1 de C, seguida de Iriartea deltoidea con 2.53 Mg ha-1 de CO 2eq y 0.69 Mg ha-1 de C, lo que refleja condiciones favorables para la acumulación de biomasa. Otras especies, como Wettinia maynensis, Ceroxylon amazonicum Galeano y Astrocaryum murumuru Mart. contribuyen en menor medida, con valores de CO 2eq entre 1.35 y 1.72 Mg ha-1 y de C entre 0.37 y 0.47 Mg ha-1 (Figura 3A).
En la altitud de 701 a 800 msnm, el almacenamiento de carbono disminuye, con Iriartea deltoidea como la especie dominante (1.67 Mg ha-1 de CO 2eq ) (Figura 3B). De 801 a 900 msnm, se observa una ligera recuperación, con Socratea exorrhiza (Mart.) H. Wendl. (2.13 Mg ha-1 de CO 2eq ) e Iriartea deltoidea (2.09 Mg ha-1) como principales contribuyentes; mientras que Oenocarpus bataua muestra una caída significativa con solo 0.14 Mg ha-1 (Figura 3C). Finalmente, en el intervalo altitudinal de 901 a 1 000 msnm el almacenamiento disminuye en todas las especies, pero Iriartea deltoidea se mantiene como la dominante con 1.07 Mg ha-1 de CO 2eq (Figura 3D).
Estos resultados coinciden con estudios que resaltan el valor de las palmas en los bosques tropicales como capturadoras eficaces de carbono, debido a la cantidad de biomasa aérea que son capaces de almacenar y a su rápida regeneración (Keeling & Phillips, 2007). Su presencia en distintos niveles del dosel, desde el herbáceo hasta el arbóreo, garantiza el almacenamiento continuo de carbono: un aspecto esencial en la mitigación del cambio climático (Goodman et al., 2013).
Usos tradicionales y valor socioeconómico de las palmas amazónicas
Las especies de Arecaceae desempeñan un papel crucial en el sustento de las poblaciones humanas amazónicas (Cámara-Leret et al., 2014). Sus hojas son empleadas en la elaboración de techos y cestas, además de algunos usos medicinales tradicionales para tratar fiebres (Vormisto et al., 2004). Son altamente apreciadas por sus frutos ricos en aceite, también se usan en la preparación de productos para el cuidado personal y se utilizan en la alimentación, así como por sus aplicaciones medicinales para el tratamiento de afecciones cutáneas (Gutsche et al., 2008). Estos múltiples usos resaltan la importancia de las palmas en la vida cotidiana de las comunidades amazónicas, además de su valor ecológico (Cuadro 6).
Cuadro 6 Principales usos registrados para las palmas en las comunidades amazónicas a lo largo del gradiente estudiado.
| Especie | Nombre común |
Usos | |||
|---|---|---|---|---|---|
| Alimentación humana y ganadera |
Medicinal | Artesanía | Construcción | ||
|
Iriartea deltoidea Ruiz & Pav. |
Pambil, Chonta | √ | √ | √ | |
|
Geonoma macrostachys Mart. |
Suqui-suqui, Pashaco | √ | √ | √ | |
|
Oenocarpus
bataua Mart. |
Ungurahua, Seje | √ | √ | √ | √ |
|
Wettinia maynensis Spruce |
Cashapona, Palmito de monte | √ | √ | √ | |
|
Ceroxylon amazonicum Galeano |
Palma de cera | √ | √ | √ | |
|
Astrocaryum
murumuru Mart. |
Murumuru | √ | √ | √ | √ |
Fuente: De la Torre et al. (2008).
Conclusiones
Este estudio resalta el papel de la familia Arecaceae en el almacenamiento de carbono en bosque siempreverde piemontano de la Amazonía Ecuatoriana. Las altitudes de 901 a 1 000 msnm presentan la mayor densidad de palmas (196 palmas ha-1) y capacidad de secuestro de carbono, con contribuciones significativas también en el intervalo de 601 a 700 msnm. Especies dominantes como Oenocarpus bataua e Iriartea deltoidea destacan en el secuestro de carbono en los bosques amazónicos ecuatorianos. Entre el gradiente altitudinal de 601 a 700 msnm, Oenocarpus bataua presenta el mayor almacenamiento con 4.89 Mg ha-1 de CO 2eq y 1.33 Mg ha-1 de C, mientras que Iriartea deltoidea almacena 2.53 Mg ha-1 de CO 2eq y 0.69 Mg ha-1 de C. En altitudes superiores, Iriartea deltoidea sigue contribuyendo consistentemente, con 1.67 Mg ha-1 de CO 2eq en 701 a 800 msnm y 1.07 Mg ha-1 en 901 a 1 000 msnm. Oenocarpus bataua es más eficiente en altitudes bajas, mientras que Iriartea deltoidea mantiene su importancia en todos los gradientes, demostrando ser clave para la restauración y captura de carbono en el ecosistema amazónico a distintas altitudes.
La diversidad y distribución uniforme de las palmas aseguran un almacenamiento de carbono en el gradiente, lo que es vital para la estabilidad ecológica y la mitigación del cambio climático. Los beneficios socioeconómicos, como la provisión de alimentos, materiales de construcción y productos artesanales, refuerzan la necesidad de estrategias de manejo que equilibren su conservación y uso sostenible. Lo anterior es fundamental, especialmente, para las comunidades locales que dependen de las palmas. Este estudio también enfatiza la importancia de desarrollar modelos alométricos específicos para las palmas, mejorando las estimaciones de biomasa y capacidad de secuestro de carbono, lo que facilitará su inclusión efectiva en programas de conservación y compensación de carbono como REDD+.

