











 text new page (beta)
text new page (beta) English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
Permalink
INTRODUCCIÓN
La Laguna de Términos en el sur del Golfo de México es uno de los ecosistemas con mayor número de estudios ecológicos realizados, presenta relevancia histórica regional por la pesca y la producción petrolera en su zona costera (Ramos-Muñoz et al., 2020). Es un ecosistema dinámicamente complejo y productivo, con alta diversidad de flora y fauna, y recursos pesqueros, ya que incluye cuatro sistemas fluvio-lagunares asociados en interacción, desde los marinos hasta los dulceacuícolas (Day et al., 2013; Yáñez-Arancibia et al., 2013). Uno de estos sistemas es el Pom-Atasta, con características ambientales y ecológicas que determinan una variedad de hábitats, los cuales condicionan los patrones espacio temporales de distribución, diversidad y abundancia de su comunidad de peces. La comunidad íctica en este sistema tiene relevancia ecológica, ya que se han registrado especies que lo utilizan para completar parte de sus ciclos vida, además varias tienen importancia económica en la pesca artesanal para consumo local en la región (Aguirre-León et al., 2010).
El sistema Pom-Atasta cuenta con variados estudios sobre dinámica hidrológica, y ecológicos de su comunidad y poblaciones de peces (Amezcua-Linares & Yáñez-Arancibia, 1980; Aguirre-León & Díaz-Ruiz, 2000, 2006; Pérez-Velázquez et al., 2007; Aguirre-León et al., 1998, 2010; Ruiz-Marín et al., 2009; Aragón-Flores et al., 2021). En este estudio se incluye el análisis de ocho años en diferentes temporadas climáticas con un enfoque de estructura de la comunidad, lo cual permite conocer el comportamiento ambiental y comunitario en una escala espacio temporal mayor y comprender el desarrollo y evolución del sistema. Los objetivos de este trabajo fueron: determinar la variación de los parámetros fisicoquímicos en el sistema, analizar la estructura de la comunidad de peces y establecer los conjuntos de peces y su relación con los ambientes que condicionan su composición y distribución en el sistema.
MATERIALES Y MÉTODOS
Área de estudio. El sistema Pom-Atasta forma parte del Área de Protección de Flora y Fauna (Plymouth APFF) de la Laguna de Términos (SEMARNAT, 1996), con categoría de Sitio Ramsar (Ficha Informativa de los Humedales de Ramsar, 2006). Se encuentra localizado hacia el oeste de dicha laguna entre los 18º 30’ y 18º 35´ latitud norte y 91º 50’ y 92º 20’ longitud oeste (Fig. 1). Es paralelo a la costa, limitado y comunicado al este por la Boca de Atasta con la Laguna de Términos y al oeste por la Laguna Colorada. Tiene una profundidad promedio de 2.7 m y una superficie aproximada de 190 km2 (Aguirre-León et al., 1998). Incluye nueve lagunas interiores de dimensiones variables, Loros, Puerto Rico, Palmas, San Carlos, El Corte, Palancares, Atasta, Pom y Colorada. Presenta un canal interno y sinuoso llamado “Torno Largo” que separa a las lagunas Palancares, Atasta y Pom del resto. No cuenta con ríos asociados, solo recibe un volumen de agua dulce a través del canal artificial llamado “del Gallego” que desemboca en la Laguna de Pom y se encuentra conectado con los ríos San Pedro y San Pablo hacia el suroeste de Tabasco y Campeche. Presenta un gradiente semipermanente de salinidad de este a oeste de 28.5 a 0.0 UPS. La vegetación ribereña incluye bosques de manglar representada por Rhizophora mangle Linneo (1753), Avicennia germinans Linneo (1764) y Laguncularia racemosa (L) Gaertner (1807). Se presentan tres temporadas climáticas, secas de febrero a mayo, lluvias de junio a septiembre y nortes de octubre a enero, cuya duración está determinada por la precipitación estacional, el patrón e intensidad de los vientos, el volumen de la descarga de los ríos y la temperatura ambiental (Ruiz-Marín et al., 2009; Aguirre-León et al., 2010).
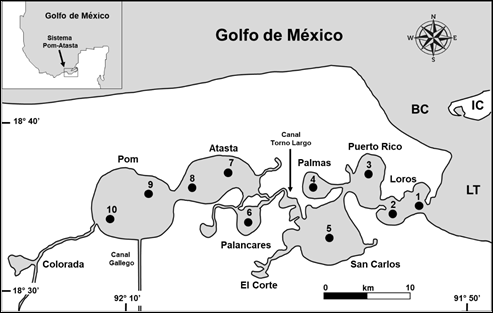
Figura 1 Ubicación geográfica del sistema Pom-Atasta en la Laguna de Términos, Campeche. Se indica la ubicación de las estaciones de muestreo (/), sus lagunas interiores y las principales características fisiográficas de la zona de estudio. LT = Laguna de Términos, BC = Boca del Carmen, IC = Isla del Carmen.
Actividades de campo. Se llevaron a cabo campañas durante los meses de octubre 2005 (O5); febrero (F6); octubre 2006 (O6); febrero 2007 (F7); octubre 2007 (O7); abril 2008 (A8); noviembre 2008 (N8); marzo 2009 (M9); febrero 2011 (F11); octubre 2011 (O11); febrero 2012 (F12); febrero 2013 (F13); septiembre 2013 (S13) y febrero 2014 (F14). En cada campaña se realizaron recolectas de peces en 10 sitios del sistema (Fig. 1) con una red de arrastre de 5.0 m de largo, boca de trabajo de 2.5 m, luz de malla de 1.8 cm, con lances de 15 min a una velocidad promedio de dos nudos para cubrir un área aproximada de arrastre de 2250m2, para un total de 140 capturas. Los peces fueron fijados con formaldehído al 10% y depositados en bolsas plásticas etiquetadas para su posterior procesamiento en el laboratorio. En cada sitio de muestreo se registró la salinidad (UPS), temperatura (oC) y oxígeno disuelto (mg/L) en superficie y fondo con un multianalizador YSI-85, la profundidad con una sonda sónica Speedtech (m) y la transparencia de la columna de agua con un disco de Secchi (m).
Actividades en laboratorio. La identificación taxonómica de las especies se determinó con los trabajos de Castro Aguirre et al. (1999), Carpenter (2002) y Miller et al. (2009). El arreglo sistemático de las especies se basó en Nelson et al. (2016) y la autoridad y nomenclatura actual de estas fue corroborada con Fricke et al. (2023). En cada sitio, los parámetros de diversidad (Shannon-Weaver H´), riqueza de especies (Margalef D), equitatividad (Pielou J´), así como la densidad (ind/m2), biomasa (g /m2) y el peso promedio (g/ind.) de la comunidad se estimaron según lo descrito por Aguirre-León et al. (2020). Las especies dominantes se determinaron mediante el Índice de Valor de Importancia (IVI= N%+P%+F%) descrito en Díaz-Ruiz et al. (2023). Los diferentes habitantes de la comunidad fueron identificados con los criterios de Potter et al. (2015) como: marinos estuarinos-oportunistas (MEO), marinos estuarinos-dependientes (MED), solo estuarinos (SE) y solo dulceacuícolas (SD). Las categorías fueron confirmadas para las especies colectadas utilizando Froese & Pauly (2023) y Robertson et al. (2023). Las categorías tróficas se definieron por el tipo de alimentación reportada en la literatura, y estas se clasificaron con los criterios propuestos por Elliott et al. (2007) y Froese & Pauly (2023) en: zooplanctívoros (ZP), especies que se alimentan predominantemente de plancton; detritívoros (DE) , especies que se alimentan principalmente de detritus y/o microfitobentos; omnívoros (OM), especies que se alimentan de plantas, epifauna e infauna; piscívoros (PI), peces que se alimentan de peces e invertebrados nectónicos y zoobentívoros (ZB), peces que se alimentan de invertebrados asociados al sustrato.
Análisis estadístico. Se aplicó análisis de varianza para determinar la existencia de diferencias significativas en los parámetros fisicoquímicos entre los sitios de muestreo en el sistema y los meses de recolecta, el mismo análisis se utilizó para los parámetros ecológicos de la comunidad para las mismas escalas de espacio y tiempo. Se requirió la justificación de la normalidad y homocedasticidad de los datos, y en todos los casos, se revisó el cumplimiento de los supuestos mediante la prueba de normalidad de Omnibus de D’Agostino-Pearson y la homogeneidad de varianzas con la prueba Levene modificada (Levene, 1960). Cuando fue necesario, se realizaron transformaciones logarítmicas. Si se cumplieron los supuestos de normalidad e igualdad de varianza, se utilizó la prueba ANOVA-F, si no se cumplieron estos supuestos, se utilizó la prueba Kruskal-Wallis-H. Se utilizó un nivel de significancia de p < 0.05 (Heiman, 2014). Estos análisis se realizaron con los programas NCSS V7 y STATISTICA V10. Se realizó Análisis de Componentes Principales (ACP) utilizando los valores espacio temporales de las variables fisicoquímicas con la finalidad de definir ambientes en el sistema, utilizando el programa Primer E V6.0 (Clarke et al., 2014).
El esfuerzo de muestreo en la comunidad se validó estimando curvas de acumulación de especies a partir de los estimadores no-paramétricos Chao2, Jackknife1, Jackknife2 y Bootstrap. Estos calculan la riqueza total de especies basada en registros de presencia-ausencia y estiman la rareza de estas por su incidencia (especies únicas y duplicadas) (Willott, 2001; Magurran, 2011). Se utilizó una base de datos de la abundancia de las especies por sitios y meses de muestreo. La construcción de estas curvas se realizó con 9999 aleatorizaciones utilizando el programa estadístico Primer-E (Clarke et al., 2014).
Para comprobar los cambios en la composición espacio temporal de la comunidad y su relación con los parámetros ambientales, fue aplicado un Análisis de Escalamiento Multidimensional no Métrico (NMDS), utilizando una base de datos de las abundancias de las especies en los sitios de muestreo, los cuales fueron transformados a raíz cuadrada para obtener una matriz de similaridad de Bray-Curtis y definir conjuntos de peces relacionados con los ambientes del sistema determinados por el ACP. Se utilizó el valor de estrés para evaluar este análisis, así como una prueba de similitud ANOSIM por pares para estimar diferencias estadísticas de la composición de las especies entre ambientes. Se utilizó el programa Primer E + Permanova V6.0 (Clarke et al., 2014).
RESULTADOS
Variación espacio temporal de los parámetros fisicoquímicos. Estos parámetros mostraron gradientes espaciales y temporales semipermanentes en su mayoría de este a oeste. En escala espacial se presentaron diferencias estadísticas significativas entre sitios, con intervalos promedio para la salinidad en superficie de 10.1 a 0.26 UPS (F=10.68, p<0.0001), en fondo de 12.3 a 0.23 UPS (F=13.13, p<0.0001), en profundidad 1.5 a 2.8 m (F=17.92, p<0.0001) y transparencia del agua de 0.51 a 0.92 m (F=2.56, p=0.0095). Asimismo, entre meses para salinidad en superficie de 10.41 a 0.16 UPS (F=5.71, p<0.0001), y de fondo 10.43 a 0.16 UPS (F=4.73, p<0.0001), temperatura en superficie de 31.28 a 23.18 oC (H=114.13, p<0.0001), y de fondo de 30.29 a 22.77 oC (H=122.46, p<0.0001), oxígeno disuelto en superficie de 8.3 a 4.0 mg/L (H=80.04, <0.0001), y de fondo de 5.88 a 2.92 mg/L) (H=57.23, p<0.0001), profundidad de 2.37 a 2.7 m (F=2.08, p=0.0195) y transparencia de 1.14 a 0.45 m (H=30.62, p=0.0038).
El comportamiento de los gradientes ambientales se reflejó en la ordenación del ACP (Fig. 2) de los sitios de muestreo en el sistema. Este análisis indicó que el 86.7% de la variación ambiental fue explicada por los primeros 2 componentes. El eje 1 con eigenvalor de 4.76, se relacionó con la salinidad, temperatura, profundidad y transparencia. El eje 2 con eigenvalor de 2.18 tuvo mayor relación con el oxígeno disuelto. Se observó la formación de tres grupos de sitios que representan ambientes con características fisicoquímicas similares: estaciones 1 a 3 con predominancia mesohalina (sup. = 8.22, fon. = 9.32 UPS) menor profundidad y transparencia (= 1.77 m y = 0.57 m) respectivamente; estaciones 4 a 8 como oligohalinas (sup. = 2.22, fon. = 2.23 UPS), profundidad y transparencia mayores ( = 2.02 m y = 0.75 m) respectivamente, y estaciones 9 y 10 principalmente dulceacuícolas (sup. = 0.30, fon. = 0.30 UPS) con el máximo de profundidad y transparencia ( = 2.57 m, y = 0.89 m) respectivamente.
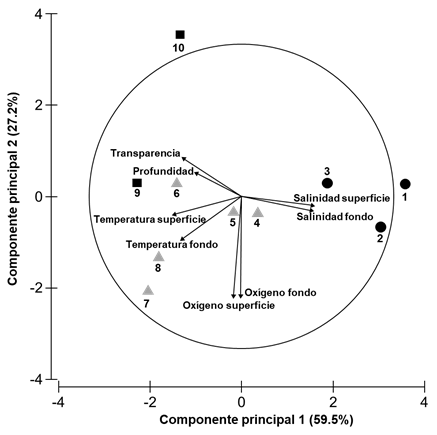
Figura 2 Análisis de componentes principales (ACP) de las variables fisicoquímicas en escala espacio temporal en el sistema Pom-Atasta, Campeche. El círculo respresenta las correlaciones entre las variables ambientales y los sitios de muestreo. Los símbolos corresponden a los ambientes mesohalino (/), oligohalino (/) y dulceacuícola (/).
Composición de la comunidad. Se recolectaron un total de 5359 individuos pertenecientes a 10 órdenes, 26 familias, 50 géneros y 59 especies (Tabla 1). Las familias mejor representadas fueron Sciaenidae con 8 especies, Gerreidae con 6, Carangidae y Cichlidae con 4. El número de especies varió entre sitios de 35 (59.3%) en la estación 1 a 14 (23.7%) en la estación 9. Entre meses de 35 (59.3%) en octubre 2007 a 15 (25.4%) en febrero 2006. Las especies más abundantes fueron Cathorops aguadulce (Meek, 1904) (30.8%), Diapterus rhombeus (Cuvier, 1829) (24.0%) y Eugerres plumieri (a) (10.4%) (Tabla 1). Se registraron tres especies exóticas Oreochromis niloticus (Linnaeus, 1758), Pterygoplichthys pardalis (Castelnau,1855) y Pterygoplichthys disjunctivus (Weber, 1991).
Tabla 1 Lista de familias y especies colectadas en el sistema fluvio deltaico Pom-Atasta, Campeche
| Especies | AB | Número | N (%) | Peso (g) | P (%) | Frec. (%) | IVI (%) | CE | CT | DA |
| Dasyatidae | ||||||||||
| Hypanus sabinus (Lesueur, 1824) | Hs | 69 | 1.30 | 42403.5 | 25.700 | 5.74 | 32.74 | MED | ZB | O |
| Lepisosteidae | ||||||||||
| Atractosteus tropicus Gill, 1863 | At | 1 | 0.02 | 2000 | 0.130 | 0.13 | 0.28 | SD | PI | D |
| Engraulidae | ||||||||||
| Anchoa mitchilli (Valenciennes, 1848) | Am | 64 | 1.20 | 58.2 | 0.040 | 3.99 | 5.23 | MEO | ZP | O |
| Clupeidae | ||||||||||
| Dorosoma anale Meek, 1904 | Dan | 5 | 0.10 | 23 | 0.014 | 0.39 | 0.50 | MED | ZP | O |
| Dorosoma petenense (Günther, 1867) | Dp | 15 | 0.30 | 335.5 | 0.648 | 0.65 | 1.60 | MED | ZP | O |
| Opisthonema oglinum (Lesueur, 1818) | Oo | 4 | 0.07 | 24.6 | 0.010 | 0.39 | 0.47 | MEO | ZB | M |
| Locariidae | ||||||||||
| Pterygoplichthys pardalis (Castelnau,1855) | Pp | 12 | 0.21 | 2316.3 | 1.400 | 1.20 | 2.81 | SD | DE | O |
| Pterygoplichthys disjunctivus (Weber, 1991) | Pd | 1 | 0.02 | 297.8 | 0.180 | 0.20 | 0.40 | SD | DE | D |
| Ariidae | ||||||||||
| Ariopsis felis (Linnaeus, 1766) | Af | 17 | 0.30 | 2362.6 | 1.400 | 1.55 | 3.25 | MED | ZB | O |
| Bagre marinus (Mitchill, 1815) | Bm | 33 | 0.60 | 298.6 | 0.200 | 0.65 | 1.45 | MEO | ZB | M |
| Cathorops aguadulce* (Meek, 1904) | Ca | 1652 | 30.80 | 39673.8 | 24.000 | 12.75 | 67.55 | SE | ZB | O |
| Synodontidae | ||||||||||
| Synodus foetens (Linnaeus, 1766) | Sfo | 16 | 0.20 | 299.9 | 0.200 | 0.65 | 1.05 | MEO | PI | M |
| Batrachoididae | ||||||||||
| Opsanus beta (Goode & Bean, 1880) | Ob | 28 | 0.50 | 1678.8 | 1.000 | 2.07 | 3.57 | MED | ZB | O |
| Syngnathidae | ||||||||||
| Syngnathus louisianae Günther, 1870 | Slo | 1 | 0.02 | 3.6 | 0.002 | 0.13 | 0.15 | MEO | ZB | M |
| Syngnathus scovelli (Evermann & Kendall, 1896) | Ss | 1 | 0.02 | 0.2 | 0.000 | 0.13 | 0.15 | MEO | ZB | O |
| Centropomidae | ||||||||||
| Centropomus parallelus Poey, 1860 | Cpa | 20 | 0.40 | 1878.1 | 1.100 | 1.81 | 3.31 | MED | ZB | O |
| Centropomus undecimalis (Bloch,1792) | Cu | 5 | 0.10 | 912.2 | 0.600 | 0.55 | 1.25 | MED | PI | O |
| Carangidae | ||||||||||
| Caranx hippos (Linnaeus, 1766) | Ch | 1 | 0.02 | 3.8 | 0.002 | 0.13 | 0.15 | MEO | PI | M |
| Chloroscombrus chrysurus (Linnaeus, 1766) | Cc | 4 | 0.07 | 16.7 | 0.010 | 0.39 | 0.47 | MEO | PI | M |
| Hemicaranx amblyrhynchus (Cuvier, 1833) | Ha | 13 | 0.20 | 50 | 0.030 | 0.39 | 0.62 | MEO | PI | M |
| Oligoplites saurus (Bloch & Schneider, 1801) | Os | 1 | 0.02 | 1 | 0.001 | 0.13 | 0.15 | MEO | PI | M |
| Lutjanidae | ||||||||||
| Lutjanus synagris (Linnaeus, 1758) | Ls | 8 | 0.15 | 332.2 | 0.200 | 0.26 | 0.61 | MEO | PI | M |
| Gerreidae | ||||||||||
| Diapterus auratus Ranzani, 1848 | Da | 12 | 0.20 | 98.3 | 0.060 | 0.65 | 0.91 | MEO | ZB | O |
| Diapterus rhombeus * (Cuvier, 1829) | Dr | 1283 | 24.00 | 5000.5 | 3.028 | 8.90 | 35.93 | MEO | OM | M |
| Eucinostomus argenteus Baird & Girard, 1855 | Ear | 110 | 2.10 | 706.2 | 0.430 | 2.44 | 4.97 | MEO | OM | M |
| Eucinostomus gula (Quoy & Gaimard, 1824) | Eg | 199 | 3.70 | 897.7 | 0.540 | 2.30 | 6.54 | MED | ZB | M |
| Eucinostomus melanopterus (Bleeker, 1863) | Em | 20 | 0.40 | 168.7 | 0.100 | 1.30 | 1.80 | MED | ZB | M |
| Eugerres plumieri* (Cuvier, 1830) | Ep | 559 | 10.40 | 32556.2 | 19.710 | 12.34 | 42.45 | MED | OM | O |
| Haemulidae | ||||||||||
| Conodon nobilis (Linnaeus, 1758) | Cn | 1 | 0.02 | 1.2 | 0.001 | 0.13 | 0.15 | MEO | ZB | O |
| Sparidae | ||||||||||
| Archosargus probatocephalus (Walbaum, 1792) | Ap | 10 | 0.20 | 3010 | 1.820 | 1.30 | 3.32 | MEO | OM | O |
| Calamus penna (Valenciennes, 1830) | Cp | 1 | 0.02 | 1 | 0.001 | 0.13 | 0.15 | MEO | ZB | O |
| Polynemidae | ||||||||||
| Polydactylus octonemus (Girard, 1858) | Po | 25 | 0.50 | 234.4 | 0.140 | 0.26 | 0.90 | MEO | ZB | M |
| Sciaenidae | ||||||||||
| Bairdiella chrysoura (Lacepéde, 1802) | Bc | 19 | 0.40 | 337.8 | 0.200 | 0.91 | 1.51 | MED | ZB | M |
| Bairdiella ronchus (Cuvier, 1830) | Br | 295 | 5.50 | 9559.5 | 5.790 | 0.91 | 12.20 | MEO | PI | M |
| Cynoscion arenarius Ginsburg, 1930 | Car | 43 | 0.80 | 210.1 | 0.130 | 1.42 | 2.35 | MED | ZB | M |
| Cynoscion nebulosus (Cuvier, 1830) | Cne | 4 | 0.07 | 212.5 | 0.130 | 0.26 | 0.46 | MEO | PI | M |
| Cynoscion nothus (Holbrook, 1848) | Cno | 25 | 0.50 | 124.7 | 0.080 | 0.52 | 1.10 | MEO | ZB | M |
| Menthicirrhus americanus (Linnaeus, 1758) | Ma | 1 | 0.02 | 7 | 0.103 | 0.13 | 0.25 | MEO | ZB | M |
| Micropogonias undulatus (Linnaeus, 1766) | Mun | 156 | 2.90 | 878.2 | 0.530 | 5.44 | 8.87 | MED | ZB | M |
| Stellifer lanceolatus (Holbrook, 1855) | Sl | 6 | 0.10 | 109.9 | 0.070 | 0.26 | 0.43 | MEO | ZB | M |
| Cichlidae | ||||||||||
| Mayaheros urophthalmus (Günther,1862) | Mu | 1 | 0.02 | 169.7 | 0.100 | 0.13 | 0.25 | SD | OM | O |
| Oreochromis niloticus (Linnaeus, 1758) | On | 3 | 0.06 | 1612.7 | 1.000 | 0.39 | 1.45 | SD | OM | O |
| Oscura heterospila (Hubbs, 1936) | Oh | 10 | 0.19 | 1488.8 | 0.900 | 0.78 | 1.87 | SD | ZB | O |
| Thorichthys helleri (Steindachner, 1864) | Th | 8 | 0.15 | 314.4 | 0.200 | 0.26 | 0.61 | SD | ZB | O |
| Eleotridae | ||||||||||
| Dormitator maculatus (Bloch, 1792) | Dm | 2 | 0.04 | 109.8 | 0.070 | 0.26 | 0.37 | SE | DE | D |
| Eleotris amblyopsis (Cope, 1871) | Ea | 1 | 0.02 | 12.1 | 0.010 | 0.13 | 0.16 | SE | ZB | D |
| Gobiidae | ||||||||||
| Bathygobius soporator (Valenciennes, 1837) | Bs | 1 | 0.02 | 0.7 | 0.000 | 0.13 | 0.15 | MEO | ZB | O |
| Gobionellus oceanicus (Pallas, 1770) | Go | 26 | 0.49 | 226.5 | 0.140 | 1.55 | 2.18 | MEO | OM | O |
| Ephippidae | ||||||||||
| Chaetodipterus faber (Broussonet, 1782) | Cf | 1 | 0.02 | 750.4 | 0.450 | 0.20 | 0.67 | MEO | OM | M |
| Trichiuridae | ||||||||||
| Trichiurus lepturus Linnaeus, 1758 | Tl | 3 | 0.06 | 80.2 | 0.050 | 0.26 | 0.37 | MEO | PI | M |
| Paralichthyidae | ||||||||||
| Citharichthys spilopterus Günther, 1862 | Cs | 177 | 3.30 | 1314.9 | 0.800 | 5.71 | 9.81 | MED | ZB | M |
| Cyclopsetta fimbriata (Goode & Bean, 1885) | Cfi | 5 | 0.09 | 20.4 | 0.010 | 0.39 | 0.49 | MEO | ZB | M |
| Etropus crossotus Jordan & Gilbert, 1882 | Ec | 4 | 0.07 | 37.7 | 0.020 | 0.49 | 0.58 | MEO | ZB | M |
| Achiridae | ||||||||||
| Achirus lineatus (Linnaeus, 1758) | Al | 91 | 1.70 | 467.7 | 0.300 | 5.55 | 7.55 | MED | ZB | O |
| Trinectes maculatus (Bloch & Schneider, 1801) | Tm | 26 | 0.50 | 131 | 0.080 | 1.98 | 2.56 | MED | ZB | O |
| Cynoglossidae | ||||||||||
| Symphurus civitatum Ginsburg, 1951 | Sc | 1 | 0.02 | 7.7 | 0.005 | 0.13 | 0.15 | MEO | ZB | M |
| Symphurus plagiusa (Linnaeus, 1766) | Sp | 1 | 0.02 | 49.5 | 0.030 | 0.13 | 0.18 | MEO | ZB | M |
| Tetraodontidae | ||||||||||
| Sphoeroides nephelus (Goode & Bean, 1882) | Sn | 20 | 0.40 | 116 | 0.070 | 0.40 | 0.87 | MEO | DE | M |
| Sphoeroides testudineus (Linnaeus, 1758) | St | 238 | 4.40 | 9170 | 5.600 | 7.23 | 17.23 | MEO | ZB | M |
| TOTALES | 59 | 5359 | 100 | 165164.50 | 100 | 100 | 300 |
Índice de Valor de Importancia (IVI). Especies dominantes (*). Componente ecológico (CE): MED = marino estuarino-dependiente, MEO = marino estuarino-oportunista, SE = solo estuarino, SD = solo dulceacuícola, categoría trófica (CT): ZP = zooplanctónicos, OM = omnívoros, DE= detritívoros, PI = piscívoros, ZB = zoobentívoros. Distribución por ambiente (DA): (M) = Mesohalino. (O) = Oligohalino, (D) = Dulceacuícola. Abreviaturas (AB) utilizadas en la Figura 6.
Las curvas de acumulación de especies (Fig. 3) mostraron que el esfuerzo de muestreo representó el número de especies en cada sitio y mes de colecta. Los estimadores sugieren que la riqueza promedio es de 74 especies, por lo que, faltarían 15 especies para completar el elenco. La representatividad del muestro varió entre el 63 % y el 88% de las especies esperadas. El máximo número de especies (81) se obtuvo con Jackknife2, y el mínimo (66) con Bootstrap.
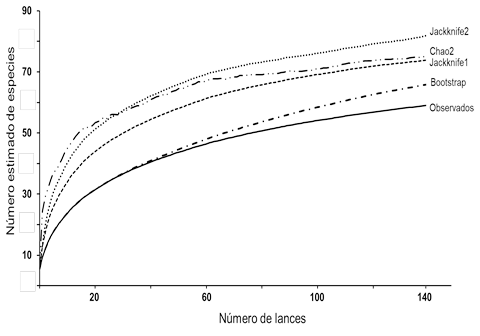
Figura 3 Curvas de acumulación de especies de peces capturadas en el sistema Pom-Atasta. Se indican las tendencias de los diferentes estimadores estadísticos calculados.
Diversidad y abundancia. La variación espacial de la diversidad H’, riqueza D y equitatividad J’ (Fig. 4 A, B, C) fue similar y estuvo relacionada con el gradiente salino, con los promedios más altos de H’ (1.56 ± 0.33) y D (2.05 ± 0.48) en las estaciones 1 y 2 con características mesohalinas. La J’ (0.86 ± 0.09) fue mayor en el ambiente oligohalino. Los promedios más bajos de H’ (0.64 ± 0.08), D (0.90 ± 0.76) y J’ (0.60 ± 0.46) se registraron en las estaciones 9 y 10 del ambiente dulceacuícola. El análisis de varianza indicó diferencias significativas para H’ (F=5.50, p<0.0001) y D (F=4.21, p<0.0001) entre sitios de muestreo. La variación temporal de la diversidad y riqueza fue similar entre meses de estudio, los promedios más altos se registraron para H’ (1.42 ± 0.33) en abril 2008, D (1.82 ± 0.97) en octubre 2007 y J’ (0.91 ± 0.06) en febrero 2013 (Fig. 4 D, E, F). Los más bajos de H’ (0.81 ± 0.47) y D (1.00 ± 0.64) en octubre 2006 y de J’ (0.55±0.23) en octubre 2007. No se encontraron diferencias significativas entre meses (P > 0.05).
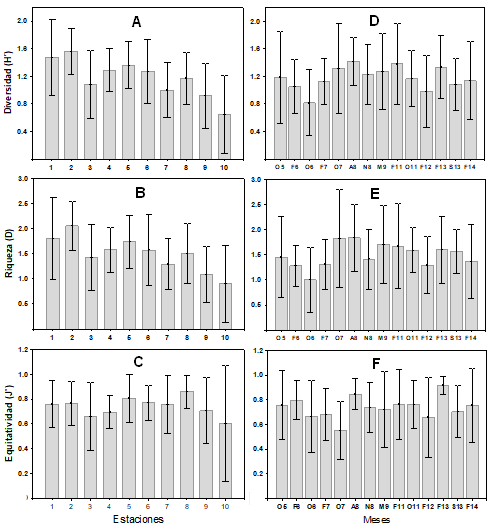
Figura 4 Variación espacial y temporal promedios (± 1 DE) de los índices Diversidad (H’), Riqueza (D) y Equitatividad (J’) de la comunidad de peces en el sistema Pom-Atasta, Campeche.
La variación espacial de la abundancia se muestra en la figura 5 (A, B, C), el valor promedio más alto de densidad (0.037 ± 0.03 ind/m2) se registró en la estación 1, la biomasa (1.02 ± 0.82 g/m2) en la estación 4 y el peso promedio (181.00 ± 178.95 g/ind.) en la estación 10. Los promedios más bajos en densidad (0.002 ± 0.001 ind/m2) y biomasa (0.29 ± 0.23 g/m2), se registraron en la estación 10 y el peso promedio (16.79 ± 10.83 g/ind) en la estación 1. Se encontraron diferencias estadísticas significativas para la densidad (H=37.45, p<0.0001) y peso promedio (H=33.05, p<0.0001) entre sitios. Temporalmente la densidad, biomasa y peso promedio (Fig. 5 D, E, F) tuvieron un comportamiento similar. Los valores promedio más altos en densidad (0.062 ± 0.056 ind/m2) y biomasa (3.15 ± 0.522 g/m2) se registraron en octubre 2007 y el peso promedio (123.68 ± 114.42 g/ind) en febrero 2013. La menor densidad (0.003 ± 0.003 ind/m2) se registró en febrero 2014, la biomasa (0.22 ± 0.10 g/m2) en septiembre 2013 y el peso promedio (29.82 ± 23.19 g/ind) en noviembre 2008. El análisis de varianza indicó diferencias significativas para la densidad (H= 25.8695, p<0.0021) y biomasa (H=25.1264, p<0.0028) entre los meses.
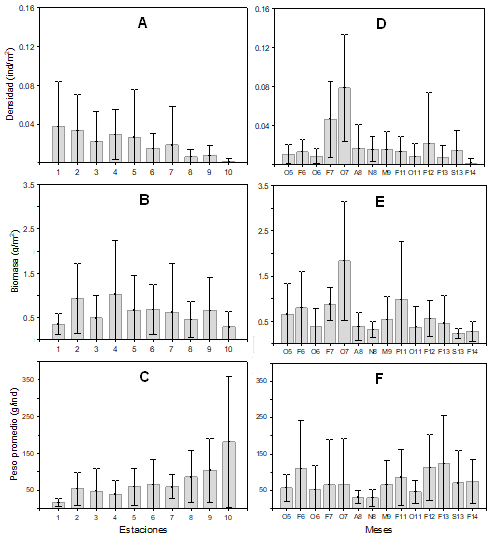
Figura 5 Variación espacial y temporal promedios (± 1 DE) de la densidad (ind/m2), biomasa (g/m2) y peso promedio (g/ind) de la comunidad de peces en el sistema Pom-Atasta, Campeche.
Especies dominantes, componentes ecológicos y categorías tróficas. El IVI indicó que tres especies fueron dominantes en el sistema, C. aguadulce (IVI=67.6), E. plumieri (IVI=42.5) y D. rhombeus (IVI=35.9), las cuales representaron el 65.2% en número y el 46.8% en peso de la captura total, con frecuencias de aparición entre 49 a 70% (Tabla 1). Otras especies menos abundantes como Hypanus sabinus (Lesueur, 1824) y Bairdiella ronchus (Cuvier, 1830) contribuyeron por su peso a la comunidad. Los componentes ecológicos estuvieron representados por 34 especies (58.0%) marinas estuarinas-oportunistas (MEO) como Sphoeroides testudineus (Linnaeus, 1758); 15 (25.0%) marinas estuarinas-dependientes (MED) como Eucinostomus gula (Quoy & Gaimard, 1824) con amplia distribución local; 7 especies (11.9%) sólo dulceacuícolas (SD) como Oscura heterospila (Hubbs, 1936) y 3 especies (5.1%) solo estuarinas (SE) como Dormitator maculatus (Bloch, 1792) (Tabla 1). La comunidad estuvo compuesta por cinco categorías tróficas, 3 especies (5.1%) zooplanctónicas (ZP) como Anchoa mitchilli (Valenciennes, 1848); 4 (6.8%) detritívoras (DE) como Sphoeroides nephelus (Goode & Bean, 1882); 8 (13.6%) omnívoras (OM) como Gobionellus oceanicus (Pallas, 1770); 11(18.6%) piscívoras (PI) como Hemicaranx amblyrhynchus (Cuvier, 1833) y 33 (55.9%) zoobentívoras (ZB) como Micropogonias undulatus (Linnaeus, 1766) (Tabla 1).
Conjuntos de especies. El NMDS (Fig. 6) muestra la ordenación de la composición y abundancia espacio temporal de las especies de la comunidad entre los ambientes del sistema, donde se observa que dicha composición se relacionó con las características ambientales que prevalecieron en cada hábitat. El ambiente mesohalino agrupó 30 especies, de las cuales 14 son exclusivas, siendo B. ronchus la más abundante. En el hábitat oligohalino 24 especies se presentaron más dispersas con 7 exclusivas, destacando M. undulatus. Estos dos últimos grupos de especies fueron los más similares (R=0.31, p= 0.001), los cuales presentaron 18 comunes. El grupo de especies del hábitat dulceacuícola sólo incluyó 4 exclusivas, y fue el más disímil respecto a los otros dos, el cual presentó a la especie exótica P. disjunctivus. Los grupos de especies entre los ambientes oligohalino y dulceacuícola fueron los menos parecidos (R=0.56, p=0.001), donde se registraron 3 comunes. Los más contrastantes fueron los conjuntos del ambiente mesohalino y dulceacuícola (R=0.82, p=0.001). En los tres ambientes se presentaron 13 especies comunes incluyendo las 3 dominantes del sistema. El estrés de la prueba fue de 0.13, siendo aceptable para este análisis. El análisis ANOSIM indicó diferencias estadísticas significativas en la composición de las especies entre los ambientes del sistema con una R global de 0.409 y p=0.001 entre grupos.

Figura 6 Análisis no-paramétrico de escalamiento multidimensional (NMDS) de las especies de peces asociada a los ambientes presentes en el sistema Pom-Atasta. Abreviaturas de las especies en Tabla 1. Especies asociadas principalmente al ambiente mesohalino (/), especies asociadas principalmente al ambiente oligohalino ( ), especies exclusivas del ambiente dulceacuícola ( ).
DISCUSIÓN
La dinámica ambiental del sistema Pom-Atasta se caracteriza por la presencia de gradientes semipermanentes de los parámetros fisicoquímicos que se relacionan con el intercambio de agua a través de su boca de conexión con el patrón de circulación de la Laguna de Términos, con la geomorfología del sistema, con los cambios climáticos regionales y el patrón de vientos asociados. Estas características condicionan el establecimiento de ambientes o hábitats en escalas espacial y temporal en el sistema como reflejo de los factores hidrográficos previamente mencionados. El análisis ambiental realizado por Aguirre-León et al. (1998) evidenció la presencia de un hábitat mesohalino y otro oligohalino que variaban en extensión a través de las temporadas climáticas de la región. En el presente estudio los parámetros fisicoquímicos que mostraron cambios espaciales significativos fueron sobre todo la salinidad, la profundidad y la transparencia del agua definiéndose tres hábitats; mesohalino, oligohalino y dulceacuícola. No obstante, en escala temporal todos los parámetros tuvieron variaciones significativas como consecuencia de la estacionalidad climática. Un rasgo fundamental para la prevalencia de los ambientes definidos también en este trabajo es la geomorfología del sistema Pom-Atasta, la cual fue analizada por Gutiérrez-Estrada et al. (1982). Este se configura por varias lagunas internas y el canal intermedio “Torno Largo” que divide a dicho sistema en dos conjuntos de cuerpos de agua, y condiciona el intercambio de agua que ingresa desde la Laguna de Términos por efecto de la marea. Otro rasgo relevante fue la construcción por parte de PEMEX del “Canal del Gallego” que desemboca en la laguna de Pom. Este canal permanece abierto desde 1992 e introduce de manera permanente agua a esta parte del sistema lo que ha condicionado la dulcificación gradual de la laguna Pom donde se localizan los sitios 9 y 10 (Aguirre-León et al., 2010). La variación de los parámetros ambientales de los hábitats definidos condiciona a su vez la organización de la ictiofauna y permite comprender en diferentes escalas los cambios comunitarios (Barletta et al. 2008; Peralta-Meixueiro & Vega-Cendejas, 2011; Molina et al., 2020).
Diversos estudios han indicado que las variables ambientales de los sistemas costeros como la temperatura y salinidad influyen de manera diferenciada en la composición de las especies, las cuales los utilizan de acuerdo con sus requerimientos biológicos, lo que permite explicar su comportamiento ecológico y ocurrencia dentro del sistema lagunar a través de las diferentes temporadas climáticas de una región (Barletta et al., 2005; Gerson Araújo et al., 2018). En este estudio, el número de especies registrado fue mayor a los registros previos en trabajos realizados en el mismo subsistema, así Amezcua Linares &Yáñez Arancibia (1980) reportaron 33 y Aguirre-León et al. (1998, 2010) 53. Debe señalarse que las especies exóticas dulceacuícolas O. niloticus, P. disjunctivus y P. pardalis se incluyeron como parte del elenco registrado desde el año 2007, las cuales han sido identificadas como uno de los riesgos ambientales críticos a los que están sujetos los hábitats acuáticos por los cambios en las características físicas del agua, así como por el desplazamiento y competencia con especies nativas (Barba-Macias et al., 2014).
Las curvas de acumulación de especies indicaron que el valor promedio estimado por estas es cercano al observado en este estudio. Este análisis sugiere que el sistema Pom-Atasta podría contener un mayor número de especies incrementando los muestreos estandarizados durante los meses de secas y lluvias, lo que resultaría en el registro de otras que lo utilizan en esas temporadas. Diversos estudios realizados en lagunas costeras tropicales y subtropicales han reconocido que las especies de peces usan de manera alterna los ambientes de estos sistemas en diferentes condiciones climáticas, reflejando la variación en la composición y riqueza de especies (Aguirre-León et al., 2014; Romero-Berny et al., 2020; Díaz-Ruiz et al., 2023).
En el sistema Pom-Atasta se observaron cambios significativos en diversidad y riqueza entre sus ambientes, caracterizados por su dinámica ambiental con valores más altos en el ambiente mesohalino debido a la presencia de especies marinas estuarinas oportunistas (MEO) como Polydactylus octonemus (Girard, 1858), Cynoscion nothus (Holbrook, 1848), Gobionellus oceanicus (Pallas, 1770) y Symphurus civitatum Ginsburg, 1951 y especies estuarinas dependientes (MED) como Dorosoma petenense (Günther, 1867), Bagre marinus (Mitchill, 1815) y Centropomus parallelus Poey, 1860. Este comportamiento es similar para la Laguna Pueblo Viejo (Castillo-Rivera et al., 2002), Laguna La Mancha (Díaz-Ruiz et al., 2018) y la Laguna del Ostión (Aguirre-León et al., 2020), donde la mayor diversidad y riqueza se presentaron en ambientes cercanos a la boca de conexión. En el ambiente dulceacuícola se registraron los valores más bajos de estos parámetros, lo que indica que un menor número de especies utilizan este hábitat (Barletta et al., 2005) como Atractosteus tropicus Gill, 1863 y Thorichthys helleri (Steindachner, 1864). Temporalmente en el mes de abril 2008 (secas) la diversidad y riqueza fue mayor debido a la presencia de especies como Opsanus beta (Goode & Bean, 1880), S. civitatum, Trichiurus lepturus Linnaeus, 1758 y Synodus foetens (Linnaeus, 1766) que sólo se presentaron durante este mes. Sin embargo, también se observan valores altos durante septiembre 2013 (lluvias), debido a que ingresan al sistema especies marinas estuarinas dependientes (MED) en busca de protección, alimento y reproducción como Centropomus undecimalis (Bloch,1792). Se ha observado que este comportamiento está relacionado con el incremento en la producción de materia orgánica y nutrientes en el sistema, incrementando la disponibilidad de alimento (Perera-García et al., 2011; Hernández-Vidal et al., 2014; Aguirre-León et al., 2020).
La abundancia de los peces se encuentra relacionada con la biología de las especies y con la variación estacional de los parámetros fisicoquímicos. Se observó que la densidad y biomasa fue mayor en el ambiente mesohalino, debido al desplazamiento de especies marinas estuarinas oportunistas (MEO) y dependientes (MED) como juveniles y preadultos, que migran desde el mar o Laguna de Términos hacia el sistema Pom-Atasta, para alimentarse, crecer y madurar durante los meses del año como B. ronchus, Citharichthys spilopterus Günther, 1862, E. gula, S. testudineus y M. undulatus, mostrando un incremento diferenciado en su abundancia en las temporadas del año (Aguirre-León & Díaz-Ruiz, 2000, 2006; Arceo-Carranza et al., 2013). Otras especies marinas estuarinas dependientes (MED), oportunistas (MEO) y solo estuarinas (SE) relacionadas con los cambios estacionales presentan mayor abundancia en el ambiente oligohalino como E. plumieri y C. aguadulce, la cuales presentaron peso promedio alto, así como D. rhombeus y Achirus lineatus (Linnaeus, 1758) igualmente abundantes pero con peso promedio bajo, las cuales diferencian sus ciclos de vida a lo largo del año de acuerdo con sus estrategias biológicas (Peralta-Meixueiro & Vega-Cendejas, 2011; Aguirre-León et al., 2020; Molina et al., 2020).
El estudio de las especies dominantes en sistemas costeros permite analizar las interacciones con otras especies presentes, ya que las primeras regulan la abundancia de estas últimas, por lo que influyen en la estructura de la comunidad (Potter et al., 2015; Gerson Araújo et al., 2018). En este estudio C. aguadulce, D. rhombeus y E. plumieri fueron especies dominantes bien adaptadas a los gradientes fisicoquímicos y estructura biótica del sistema Pom-Atasta, lo que ha permitido su coexistencia y utilización de sus ambientes con diferentes fines biológicos a través del tiempo. En estudios previos de este sistema en diferentes escalas temporales, Aguirre-León et al. (1998, 2010) reportaron a S. testudineus, A. mitchilli, E. gula, Ariopsis felis (Linnaeus, 1766), M. undulatus y B. ronchus también como dominantes. Todas las especies referidas han sido registradas como dominantes o abundantes en sistemas litorales del Golfo de México por Aguirre-León et al. (2020), Aragón-Flores et al. (2021), Díaz-Ruiz et al. (2018, 2023).
En el sistema Pom-Atasta los componentes ecológicos variaron a través del tiempo. Los mejor representados fueron los peces marinos estuarinos-oportunistas (MEO) como Eucinostomus argenteus Baird & Girard, 1855, A. mitchilli y B. ronchus que pueden distribuirse en todo el sistema. Asimismo, los marinos estuarinos-dependientes (MED) como H. sabinus, O. beta, M. undulatus y C. spilopterus, con amplia distribución en este sistema. Este tipo de especies migran desde la zona marina hacia los ambientes protegidos de las lagunas costeras donde encuentran áreas propicias que utilizan durante sus ciclos biológicos. Además desovan cerca de las bocas de conexión de estos sistemas y entran estacionalmente a ellos como juveniles a las áreas de crianza y como preadultos y adultos a las áreas de alimentación y protección (Sánchez-Gil et al., 2008; Franco et al., 2017; Reyes-Ramírez et al., 2018). Con menor porcentaje estuvo representado el componente solo estuarino (SE) con especies que completan sus ciclos de vida dentro del sistema como C. aguadulce optimizando los gradientes ambientales del sistema (Ayala-Pérez et al., 2008). El componente dulceacuícola (SD) presentó un porcentaje bajo de especies, cuya distribución está limitada a las salinidades más bajas. Del comportamiento de estos componentes ecológicos destaca la importancia que tienen los ecosistemas costeros para las especies de peces que los utilizan en alguna etapa de su ciclo de vida (Childs et al., 2015; Potter et al., 2015; Dames et al., 2017; Steffen et al., 2023).
La composición de las categorías tróficas permite analizar la estructura ecológica y función de los conjuntos de peces en ecosistemas costeros, así como la evolución natural de sistema en diferentes escalas (Elliott et al., 2007; Buchheister & Latour, 2015). El análisis de la composición trófica en el sistema Pom-Atasta mostró que las especies zoobentívoras como C. aguadulce, E. gula, M. undulatus, S. testudineus fueron las mejor representadas distribuyéndose en áreas de manglar donde es abundante la epifauna e infauna. Las especies piscívoras como C. undecimalis, Chloroscombrus chrysurus (Linnaeus, 1766), H. amblyrhynchus y Lutjanus synagris (Linnaeus, 1758) fueron el segundo grupo trófico en el sistema, mientras que los zooplanctívoros, omnívoros y detritívoros presentaron una frecuencia baja en este. Dicho comportamiento está relacionado con la disponibilidad de alimento, principalmente durante los meses fríos de la región donde las especies marinas estuarinas-oportunistas y estuarinas-dependientes entran al sistema. Se ha observado que la composición de los grupos tróficos cambia en función de las variaciones ambientales como la temperatura, salinidad, profundidad, turbidez y tipo de sustrato, así como por la presencia de vegetación sumergida y las etapas específicas de su biología como los cambios ontogenéticos, edad y sexo, por lo que es relevante reconocer que su alimentación y hábitos alimenticios reflejan los patrones estacionales de uso de los sistemas. Este enfoque ha sido utilizado por otros autores para interpretar el comportamiento de las categorías tróficas (Elliott et al., 2007; Abrantes et al., 2015; Díaz-Ruiz et al., 2023).
El análisis NMDS permitió identificar conjuntos de peces en cada ambiente del sistema condicionados por sus adaptaciones biológicas y por la interacción con la Laguna de Términos. El conjunto de especies asociadas al ambiente mesohalino, estuvo compuesto por especies marino-estuarinas oportunistas (MEO), algunas con abundancias bajas como B. marinus, Syngnathus scovelli (Evermann & Kendall, 1896), Cynoscion nebulosus (Cuvier, 1830) y S. nephelus, las cuales han sido registradas como especies dominantes en Laguna de Términos y La Sonda de Campeche en condiciones de mayor salinidad (Peralta-Meixueiro & Vega-Cendejas, 2011; Torruco et al., 2018). La boca de conexión del sistema Pom-Atasta se considera un área de tránsito donde se registra mayor riqueza de especies y menor abundancia; así, Oligoplites saurus (Bloch & Schneider, 1801), Caranx hippos (Linnaeus, 1766), S. civitatum, Symphurus plagiusa (Linnaeus, 1766) y Syngnathus louisianae Günther, 1870, se colectaron durante los meses de secas y lluvias en el ambiente mesohalino. Al interior del sistema, su geomorfología presenta áreas más protegidas con menor salinidad donde el conjunto de especies asociadas al ambiente oligohalino estuvo integrado por especies marino-estuarinas dependientes (MED) como D. petenense, C. parallelus, C. spilopterus y Trinectes maculatus (Bloch & Schneider, 1801) con abundancia intermedia. Entre estos dos primeros ambientes se comparten especies con mayor abundancia como B. ronchus, S. testudineus, E. gula, C. spilopterus, M. undulatus, E. argenteus, A. lineatus que utilizan el sistema para crecimiento y maduración. Asimismo, prevalecen en los tres ambientes las especies dominantes C. aguadulce, D. rhombeus y E. plumieri que se caracterizan por tolerar amplias variaciones de salinidad y optimizan el sistema para su reproducción y crianza (Aguirre-León & Díaz-Ruiz, 2000, 2006). El conjunto de especies que caracteriza al ambiente dulceacuícola presentó abundancias bajas como A. tropicus, D. maculatus, Eleotris amblyopsis (Cope, 1871) y P. disjunctivus que se registraron principalmente durante el mes de lluvias. Los conjuntos de peces definidos pueden variar principalmente por los cambios en las preferencias de utilización de los hábitats que ocurren durante los ciclos de vida, los patrones de migración y alimentación de cada especie, lo cual dificulta ubicarlos en una categoría ecológica específica relacionada con un ambiente particular (Childs et al., 2015; Potter et al., 2015; Steffen, 2023).
Este estudio integró información sobre la dinámica de la comunidad de peces en el sistema Pom-Atasta relacionada con la variación ambiental de sus hábitats, como marco de referencia para estudios futuros, la cual proporciona conocimiento complementario y criterios ecológicos que permitan la conservación de su biodiversidad en la región.

