











 text new page (beta)
text new page (beta) Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
Permalink
Introducción
Los sistemas agroforestales (SAF) son reservorios de biodiversidad, en la que están incluidos los hongos micorrízicos arbusculares (HMA -Glomeromycota-) (Udawatta, Rankoth y Jose, 2019; Fahad et al., 2022). Estos hongos forman la asociación micorrízica arbuscular con más del 80% de las plantas terrestres, incluyendo las de interés forestal y agrícola (Smith y Read, 2010). Se ha reportado que, el micelio de estos hongos establece redes de intercambio de nutrientes entre las plantas cultivadas y perennes dentro de los SAF (Marney y Borden, 2019; Gutiérrez-Nuñez y Gavito, 2024). Sin embargo, son escasas las investigaciones que consideran a los HMA de SAF como un recurso biológico nativo con potencial para favorecer el desarrollo de los cultivos en ecosistemas secos (Joshi, Deepa, Joshi y Sharma, 2023). En los ecosistemas semiáridos, donde existe un elevado estrés ambiental y los SAF han sido escasamente explorados comparados con los ecosistemas húmedos, los HMA asociados a estos SAF semiáridos podrían ser cruciales para la sobrevivencia de las plantas cultivadas en vistas de mantener la salud del suelo, producción de alimento y alcanzar la soberanía agroalimentaria (Fahad et al., 2022; Joshi et al., 2023).
Hace una década, se señaló la importancia de elaborar biofertilizantes a partir de microorganismos nativos (Coutiño-Puchuli, Peña e Infante, 2023). Dichos inoculantes deben ser de alta calidad, lo que significa que deben estar constituidos por más de una especie, tener abundantes propágulos (esporas, raíces colonizadas y micelio externo) con capacidad de colonizar a la planta hospedera y ser efectivos para el desarrollo de su hospedero a través del tiempo. Asimismo, el inóculo resultante debe estar libre de agentes fitopatógenos y, finalmente, mantenerse viable a largo plazo (González-Chávez, 2002; Rouphael et al., 2015).
Los SAF ofrecen nichos únicos con diferentes condiciones y recursos que promueven las interacciones bióticas y abióticas, las cuales se afectan por el manejo agrícola (Fahad et al., 2022). Por lo que conocer el papel funcional de los HMA como inoculantes en ecosistemas semiáridos, bajo contextos particulares de manejo del suelo, es prioritario para garantizar no sólo la concentración de propágulos y la conservación de los HMA, sino también para promover el desarrollo de los diferentes cultivos de interés agrícola (Montaño et al., 2012).
En San Luis Atolotitlán, en el Valle semiárido de Tehuacán-Cuicatlán, Puebla-Oaxaca, México, el matorral xerófilo con cactáceas columnares es uno de los tipos de vegetación nativa más importante que ha sido transformado, desde tiempos prehispánicos, en áreas para el cultivo mediante un manejo agroforestal sustentable tradicional que permite la conservación de la diversidad vegetal (Rendón-Sandoval et al., 2024). En este ecosistema semiárido, la “milpa-chichipera” es un tipo de manejo agroforestal tradicional integrado por islas de vegetación nativa rodeadas por áreas abiertas destinadas al cultivo de maíz y en las que predominan Myrtillocactus chichipe (Rol.-Goss.) P.V. Heath (=Polaskia chichipe (Rol.-Goss.) Backeb.) (Cactaceae), un cactus columnar endémico y otras especies vegetales nativas que mantienen los pobladores, razón por la que a este SAF se le denominó “milpa-chichipera” (Moreno-Calles, Casas, García y Torres, 2012). Este SAF tradicional, además de favorecer el mantenimiento de la biodiversidad de flora y fauna (Moreno-Calles et al., 2010), como otros SAF tradicionales en la región (Rendón-Sandoval et al., 2024), contiene plantas altamente micotróficas que promueven la presencia y diversidad de los HMA (Camargo-Ricalde, Dhillion y Jiménez, 2003; Chimal-Sánchez, Montaño, Camargo, García y Hernández, 2022), los cuales podrían ser críticos en el crecimiento y nutrición de las plantas en estas condiciones de aridez y de baja fertilidad del suelo (García-Licona et al., 2017) y, por lo tanto, podrían conformar biofertilizantes nativos potenciales para la agricultura en esta región semiárida de México.
En el mundo, una de las hortalizas reconocidas por su alto valor nutricional, comercial y de consumo es la lechuga (Lactuca sativa L. -Asteraceae-) (Medina-Lozano, Bertolín y Díaz, 2021). En México, esta hortaliza es producida principalmente en Baja California, Zacatecas, Aguascalientes, Jalisco, Guanajuato y Puebla, durante los periodos febrero-abril y julio-agosto y el país exporta el 49% de la producción anual (SIAP, 2018; Shatilov, Razin y Ivanova, 2019). El cultivo de lechuga se lleva a cabo en todas las épocas del año (SIAP, 2018), por lo que está expuesto a variaciones estacionales de la lluvia y es altamente dependiente del riego, lo que aumenta su susceptibilidad al estrés hídrico y reduce sus posibilidades de cultivarse en regiones semiáridas (Pertierra y Quispe, 2020). Algunos estudios indican consorcios particulares de HMA asociados a la lechuga (Acoltzi-Conde, Chimal, Tovar y Díaz, 2024), con un efecto favorable que posiblemente varía en magnitud con la composición fúngica (Tapia-Goné et al., 2010; Ley-Rivas, Aliaga, Moron y Furrazola, 2011; Ley-Rivas, Ricardo, Sánchez, Furrazola y Gómez, 2016; Cela et al., 2022). Ante este escenario, es necesario considerar a los HMA de los SAF como posibles biofertilizantes micorrízicos nativos que otorgarían beneficios para asegurar la supervivencia y producción de esta hortaliza en suelos con estrés hídrico y nutrimental como en los ecosistemas semiáridos.
Por lo anterior, el objetivo del presente estudio fue evaluar la efectividad de tres inóculos de HMA procedentes de tres condiciones de uso del suelo: islas de vegetación y áreas abiertas del sistema agroforestal “milpa-chichipera” y matorral xerófilo en el Valle semiárido de Tehuacán-Cuicatlán, Puebla-Oaxaca, México, sobre el crecimiento y la nutrición de la lechuga. La hipótesis fue que los tres inóculos micorrízicos favorecerían el crecimiento y nutrición de las plantas de lechuga, pero su efectividad variaría en función de su procedencia según la condición de uso del suelo.
Materiales y Métodos
Área de estudio
Los inóculos de hongos micorrízicos arbusculares (HMA) se recolectaron en la localidad de San Luis Atolotitlán, ubicada dentro del Valle semiárido de Tehuacán-Cuicatlán, Puebla-Oaxaca, México, en las coordenadas 17° 32’‑18° 53’ N y 96° 55’- 97º 44’ O. El clima es semiárido, con una temperatura y precipitación medias anuales de 21 °C y 434 mm, respectivamente, con una marcada estacionalidad de la lluvia (Valiente-Banuet, 19911). La vegetación predominante es matorral xerófilo con alta densidad de cactáceas columnares (Valiente-Banuet et al., 2009). El suelo dominante en el área de colecta es Leptosol (IUSS, 2015).
Colecta de suelo y hongos micorrízicos arbusculares
En el área de estudio se ubicaron tres condiciones de uso del suelo: i) áreas abiertas al cultivo de maíz combinado con frijol y calabaza (milpa) rodeadas por ii) islas de vegetación ambos dentro del sistema agroforestal (SAF) “milpa-chichipera” denominado así por Moreno-Calles et al. (2012), y iii) matorral xerófilo nativo. En cada condición se establecieron cinco parcelas de 20×50 m (1000 m2) separadas por al menos un kilómetro. En cada parcela se establecieron tres transectos en los que se colectaron 15 submuestras de suelo que fueron mezcladas para conformar una muestra compuesta a partir de la cual se aislaron e identificaron los HMA con base en los rasgos morfológicos de las esporas y en las descripciones originales de las especies de HMA. El suelo se colectó de los primeros 15 cm de profundidad, debido a que en esta primera capa de suelo se concentra la mayor biomasa de raíces, microorganismos y abundancia de esporas de HMA (Pavón y Briones, 2000; Rodríguez-Zaragoza et al., 2008; Chimal-Sánchez et al., 2022). El suelo, con los HMA y fragmentos de raíces, procedente de cada condición fue utilizado como inóculo natural. La caracterización taxonómica de los HMA de cada inóculo permitió identificar 30 morfoespecies de HMA en el suelo de las islas de vegetación, 26 en el suelo de las áreas abiertas y 25 en el suelo del matorral xerófilo (Cuadro 1).
Cuadro 1: Morfoespecies de hongos micorrízicos arbusculares en tres condiciones de uso del suelo: áreas abiertas e islas de vegetación del sistema agroforestal “milpa-chichipera” y matorral xerófilo en el Valle semiárido de Tehuacán-Cuicatlán, Puebla-Oaxaca, México.
Table 1: Morphospecies of arbuscular mycorrhizal fungi in different land uses: open areas and vegetation islands from agroforestry system “milpa-chichipera”, and xeric shrub at semiarid Valley of Tehuacan-Cuicatlan, Puebla-Oaxaca, Mexico.
| Sistema agroforestal "milpa-chichipera" | Matorral xerófilo | ||
| Morfoespecies de hongos micorrízicos arbusculares | Áreas abiertas | Islas de vegetación | |
| Acaulospora aff. colliculosa Kaonongbua, J.B. Morton & Bever | X | X | X |
| Acaulospora sp.1 | X | X | X |
| Acaulospora excavata Ingleby & C.Walker | X | X | X |
| Acaulospora laevis Gerd. & Trappe | X | X | |
| Acaulospora mellea Spain & N.C.Schenck | X | X | X |
| Acaulospora morrowiae Spain & N.C. Schenck | X | X | X |
| Acaulospora reducta Oehl, B.T. Goto & C.M.R. Pereira | X | ||
| Acaulospora rehmii Sieverd. & S. Toro | X | X | |
| Acaulospora scrobiculata Trappe | X | X | |
| Acaulospora spinosissima Oehl, Palenz., I. C. Sánchez, Tchabi, Hount. & G. A. Silva | X | ||
| Acaulospora sp. 2 | X | ||
| Acauslospora sp. 3 | X | ||
| Acaulospora sp. 4 | X | ||
| Acaulospora sp. 5 | X | ||
| Acaulospora sp. 6 | X | ||
| Acaulospora sp.7 | X | X | |
| Ambispora reticulata Oehl & Sieverd., E. | X | ||
| Ambispora appendicula (Spain, Sieverd. & N.C. Schenck) C. Walker | X | ||
| Ambispora sp. 1. | X | ||
| Diversispora sp. 1 | X | ||
| Diversispora spurca (C.M. Pfeiff., C. Walker & Bloss) C. Walker & Schüβler | X | X | X |
| Diversispora trimurales C. Walker & Schüβler | X | ||
| Diversispora sp.3 | X | X | |
| Sieverdingia tortuosa (N.C. Schenck & G.S. Sm.) Blaszk., Niezgoda & B.T. Goto. | X | ||
| Corymbiglomus globiferum (Koske & C. Walker) Błaszk. & Chwat. | |||
| Corymbiglomus sp. 1 | X | ||
| Entrophospora claroidea (N.C. Schenck & G.S. Sm.) Blaszk., Niezgoda, B.T. Goto & Magurno. | X | X | X |
| Entrophospora etunicata (W.N.Becker & Gerd.) Błaszk., Niezgoda, B.T. Goto & Magurno. | X | X | |
| Entrophospora infrequens (I.R. Hall) R.N. Ames & R.W. Schneid | X | X | X |
| Entrophospora sp. 1 | X | ||
| Entrophospora sp. 2 | X | X | |
| Gigaspora candida Bhattacharjee, Mukerji, J.P. Tewari & Skoropad | X | X | X |
| Racocetra gregaria (N.C.Schenck & T.H.Nicolson) Oehl, F.A. Souza & Sieverd. | X | ||
| Centraspora pellucida (T.H. Nicolson & N.C. Schenck) Oehl, F.A. Souza & Sieverd. | X | X | X |
| Paraglomus bolivianum (Sieverd. & Oehl) Oehl & G.A. Silva. | X | X | |
| Glomus macrocarpum Tul. & C. Tul. | X | X | |
| Glomus spinuliferum Sieverd. & Oehl | X | X | |
| Glomus sp. 1 | X | ||
| Rhizophagus fasciculatum (Thaxt.) C. Walker & A. Schüßler | X | X | |
| Funneliformis geosporum (T.H. Nicolson & Gerd.) C. Walker & A. Schüβler | X | X | X |
| Funneliformis mosseae (T.H.Nicolson & Gerd.) C.Walker & A. Schüβler | X | X | X |
| Septoglomus mexicanum E. Chimal-Sánchez, C. Senés-Guerrero, N.M. Montaño, L. Varela, et al. | X | X | X |
| Septoglomus constrictum (Trappe) Sieverd., G.A.Silva & Oehl | X | ||
| Paraglomus occultum C.Walker) J.B.Morton & D.Redecker | X | X | |
Establecimiento del experimento
Las semillas de lechuga (Lactuca sativa L.) se sembraron en macetas de polipropileno con capacidad de 1 kg, las cuales se lavaron y desinfectaron previamente con una solución hipoclorito de sodio al 10% por 5 minutos. Cada maceta se llenó hasta la mitad con el sustrato, que consistió en una mezcla de suelo con arena sílica 50/50 (v/v) que fue homogenizada y esterilizada en autoclave a 120 °C y 15 lb de presión durante 1h, en dos ocasiones. Dicho sustrato tuvo una textura franco-arenosa (arena 75%, limo 12%, arcilla 13%) determinada por la pipeta americana; pH activo = 7.2 con potenciómetro de electrodo (suelo:agua 1:2 p/v); conductividad eléctrica= 3.2 dS m-1 con conductímetro (suelo:agua 1:2 p/v); CIC= 10.03 cmol(+) kg-1 con acetato de amonio pH = 7 y pasta de saturación; concentración de Na+= 0.12, K+= 0.58, Ca++= 3.48 y Mg++= 15.84 (cmol(+) kg-1) por pasta de saturación; CaCO3= 0.11 eq por titulación ácida; Ntotal = 0.21% por el método de micro-Kjeldalhl con H2SO4 + H2O2; PO4 -= 59.7 mg kg-1 con el método Olsen; COS= 3.3% con Walkley-Black por digestión húmeda con K2Cr2O7 y H2SO4; MOS= 6.4% a partir del valor de COS y densidad aparente = 1.3 g cm-3 por terrón parafinado; todos determinados con base en Van Reeuwijk (2003).
Los tratamientos con inóculo de HMA consistieron en 50 g del suelo colectado que contenían en promedio de 80 esporas de HMA y cuya procedencia correspondió a alguna de las tres condiciones de uso del suelo: islas de vegetación y áreas abiertas en el SAF “milpa-chichipera”, y matorral xerófilo; además de un tratamiento control (sin inocular). Las macetas con inóculo se completaron en volumen con el mismo sustrato y se sembraron las semillas de lechuga previamente desinfectadas con hipoclorito de sodio al 3% por 2 minutos. Una planta de cada maceta correspondió a la unidad experimental. Las plantas se mantuvieron en condiciones de invernadero (37 °C y humedad relativa de la Ciudad de México, abril-mayo) y se regaron con agua destilada, cada tres o cuatro días, durante tres meses.
Evaluación del crecimiento y nutrición en las plantas
Al final el periodo experimental, la parte aérea de las plantas se cosechó en cada tratamiento y se evaluó el peso fresco y seco con una balanza analítica OHAUS®. Las plantas se secaron en un horno ECOSHEl® a 65 °C durante 72 h hasta peso constante. El área foliar se midió en el programa IMAGEJ y la clorofila en hojas con un medidor SPAD marca ECO- TYS®. Además, se calculó el contenido de agua en la planta, a través de la resta del peso fresco menos el seco de la masa de las plantas y se determinó la concentración de nitrógeno (N) y fósforo (P) en el tejido vegetal por digestión húmeda microKjeldahl y titulación con ácido sulfúrico en el caso del N (Isaac y Johnson, 1976; Jones y Case, 1990) y en el caso del P, se siguió el método de Murphy y Riley (1962). El contenido de N y P en la planta se estimó multiplicando la concentración de cada bioelemento por la biomasa seca de las plantas al tiempo de la cosecha. Únicamente se evaluó la parte aérea de las plantas de lechuga, dado que la subterránea se incorporó como parte de los inoculantes micorrízicos que fueron entregados a los campesinos de la región semiárida de San Luis Atolotitlán en el Valle de Tehuacán-Cuicatlán.
Diseño experimental y análisis de datos
El diseño experimental fue completamente al azar con cinco réplicas (n= 5) por tratamiento. Los datos se analizaron con un ANOVA de una vía seguido de pruebas de medias de Tukey (P < 0.05) para evaluar el efecto de los tratamientos. Los datos se ajustaron a los supuestos de normalidad y homocedasticidad cuando fue necesario, pero se presentan en su escala original de medición. Los análisis se realizaron en el programa SAS® versión 9.4 (SAS Institute, 2014).
Resultados y Discusión
Los tratamientos de inoculación con hongos micorrízicos arbusculares (HMA) estimularon el crecimiento de las plantas de lechuga, reflejando mayor biomasa fresca y seca, y contenido de agua en la planta (Figura 1 a, b, c), así como un incremento del área foliar e índice de clorofila en hojas en comparación con el tratamiento control (Figura 2 a, b). El crecimiento vegetal favorecido por la inoculación con HMA ha sido reportado en muchas otras plantas de ecosistemas semiáridos (Camargo-Ricalde, Montaño, Reyes, Jiménez y Dhillion, 2010; Robles-Martínez, Robles, Rivera, Ortega y Pliego, 2013; Montaño-Arias et al., 2023) y hortalizas, incluida la lechuga (Cuenca, Cáceres, Oirdobro, Hasmy y Urdaneta, 2007; Tapia-Goné et al., 2010; Wee, Li, Kim y Sohn, 2010; Rouphael et al., 2015; Ley-Rivas et al., 2016; Shi et al., 2016; Carballar-Hernández, Hernández, Montaño, Ferrera y Alarcón, 2018).
Figura 1: Efectividad de inóculos de hongos micorrízicos arbusculares (HMA) provenientes de tres diferentes usos de suelo: áreas abiertas e islas de vegetación del sistema agroforestal “milpa-chichipera” y matorral xerófilo en el Valle semiárido de Tehuacán-Cuicatlán, México sobre: A) Peso fresco, B) Peso seco y C) Contenido de agua de plantas de lechuga (Lactuca sativa L.). Control = sin inocular HMA.
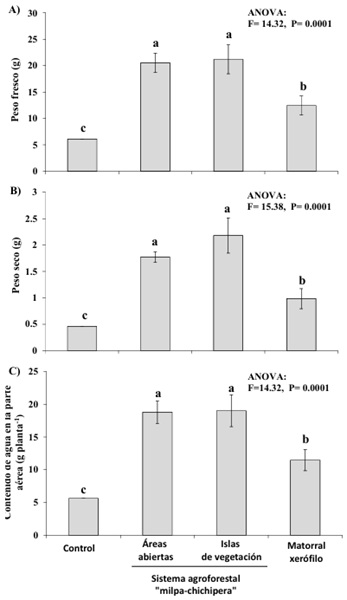
Figure 1: Effectiveness of arbuscular mycorrhizal fungal (AMF) inocula from three different land uses: open areas and vegetation islands of agroforestry system “milpa-chichipera” and xeric scrub at semiarid Tehuacan-Cuicatlan Valley on: A) Fresh weight, B) Dry weight, and C) Water content in lettuce plants (Lactuca sativa L.). Control = without AMF inoculum.
Asimismo, la inoculación con HMA promovió la acumulación de N y P en el tejido de las plantas de lechuga, por lo que el contenido de estos bioelementos, en general, fue mayor en las plantas inoculadas con HMA provenientes de las tres condiciones de uso del suelo que en las plantas sin inocular (Figura 3 a, b), con excepción del N que no difirió entre las plantas control y las inoculadas con HMA del matorral xerófilo (Figura 3b). Este resultado también coincide con el efecto positivo de la inoculación con HMA reportado en otras plantas y hortalizas como la lechuga (Azcón, Ambrosano y Charest, 2003; Cuenca et al., 2007; Rouphael et al., 2015; Ley-Rivas et al., 2016; Carballar-Hernández et al., 2018; Cela et al., 2022). Los HMA, al formar la micorriza arbuscular, contribuyen a través de su micelio a una mayor captura de agua y nutrientes del suelo por parte de las raíces (Clark y Zeto, 2000; Miransari, 2011), lo que favorece el desarrollo y la actividad fotosintética de las plantas hospederas al abastecerlas de éstos y posiblemente otros recursos, lo que podría promover la síntesis de clorofila y azúcares en las plantas (Smith y Read, 2010). En las plantas de lechuga, la simbiosis con los HMA de ambientes semiáridos podría coadyubar a subsanar las deficiencias de agua y disponibilidad de N y P en el suelo, como se ha demostrado en otros escenarios para esta hortaliza (Azcón et al., 2003; Vicente-Sánchez et al., 2014).
Los inóculos de HMA pertenecientes al SAF “milpa-chichipera”, tanto los provenientes de las islas de vegetación como los de áreas abiertas, fueron más efectivos en comparación con el inóculo proveniente del matorral xerófilo, así las plantas de lechuga de estos tratamientos registraron mayor crecimiento, almacenamiento de agua (Figura 1 a, b), actividad fotosintética (Figura 2 a, b) y contenido nutrimental de N y P en su tejido (Figura 3 a, b). No obstante, el contenido de clorofila en las hojas fue mayor y tuvo valores similares entre plantas inoculadas con HMA provenientes del matorral xerófilo e islas de vegetación del SAF “milpa-chichipera” (Figura 2b); mientras que, las plantas inoculadas con los HMA de áreas abiertas del SAF “milpa-chichipera”, tuvieron valores intermedios, pero superiores a las plantas control (Figura 2b).
Figura 2: Efectividad de inóculos de hongos micorrízicos arbusculares (HMA) provenientes de tres diferentes usos de suelo: áreas abiertas e islas de vegetación del sistema agroforestal “milpa-chichipera” y matorral xerófilo en el Valle semiárido de Tehuacán-Cuicatlán, México sobre: A) Área foliar y B) Índice de clorofila de plantas de lechuga (Lactuca sativa L.). Control = sin inocular HMA.
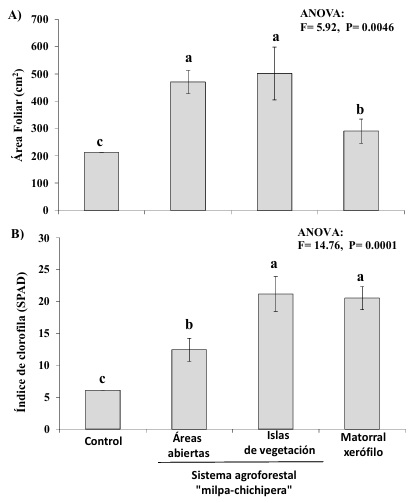
Figure 2: Effectiveness of arbuscular mycorrhizal fungal (AMF) inocula from three different land uses: open areas and vegetation islands of agroforestry system “milpa-chichipera” and xeric scrub at semiarid Tehuacan-Cuicatlan Valley on: A) Leaf area, and B) Chlorophyll index in lettuce plants (Lactuca sativa L.). Control = without AMF inoculum.
Figura 3: Contenido de fósforo (A) y nitrógeno (B) de la parte aérea de plantas de lechuga (Lactuca sativa L.) de diferentes inóculos de hongos micorrízicos arbusculares provenientes de tres diferentes condiciones de uso del suelo: áreas abiertas e islas de vegetación del sistema agroforestal “milpa-chichipera” y matorral xerófilo en el Valle semiárido de Tehuacán-Cuicatlán, México. Control = sin inocular HMA.
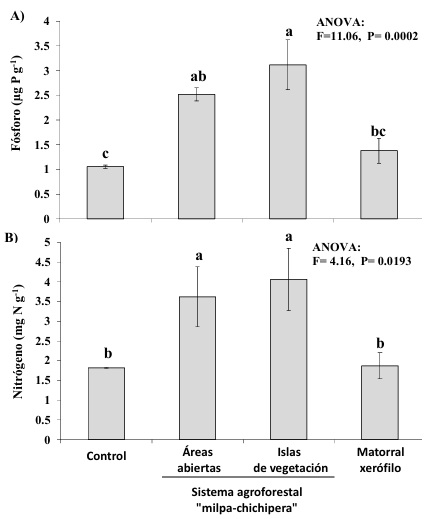
Figure 3: Phosphorus (A) and nitrogen (B) content in aboveground biomass of lettuce plants (Lactuca sativa L.) inoculated with arbuscular mycorrhizal fungi coming from three different land uses: open areas and vegetation islands of agroforestry system “milpa-chichipera” and xeric scrub at semiarid Tehuacan-Cuicatlan Valley, Mexico. Control = without AMF inoculum.
Los resultados sugieren que, en este SAF tradicional, los HMA, al interactuar con las plantas nativas preservadas en las islas de vegetación y con la milpa, son más efectivos para el desarrollo de la lechuga que los HMA que están en el matorral xerófilo. Es posible que los HMA en el suelo de este SAF estén sometidos a constantes fluctuaciones ambientales que los HMA del matorral xerófilo; por ejemplo, mayores cambios en humedad, temperatura y disponibilidad de nutrientes en el suelo, así como a diferentes hospederos a lo largo del año, por lo que la plasticidad funcional de estos hongos podría estar dirigida a contrarrestar el estrés (Branco, Schauster, Liao y Ruytinx, 2022). Esta exposición constante a condiciones ambientales estresantes, además de determinar el ensamblaje de especies de HMA en este SAF respecto del registrado en el matorral xerófilo, también podría contribuir a moldear la efectividad de los HMA para colonizar a sus hospederos y aportarles beneficios, dado que estarían más adaptados al estrés ambiental que aquellos HMA menos expuestos a variaciones en condiciones y recursos (Branco et al., 2022), como podría ocurrir en el suelo del matorral xerófilo, donde la humedad y nutrientes son mayores que en el suelo de islas de vegetación y áreas abiertas del SAF “milpa-chichipera” (datos no publicados). Como consecuencia, el matorral xerófilo contiene un inóculo micorrízico arbuscular con similar riqueza de morfoespecies de HMA que el SAF, pero con HMA menos eficientes que los inóculos micorrízicos asociados a las islas de vegetación y áreas abiertas del SAF “milpa-chichipera”, sugiriendo que el manejo tradicional podría estar seleccionando de alguna manera a largo plazo la efectividad de los HMA.
La diferencia en la composición de morfoespecies de HMA asociada a cada inóculo también podría explicar los resultados obtenidos en este estudio, ya que, diferentes especies de HMA tienen capacidades distintas para soportar condiciones adversas, lo que hace que el estrés ambiental afecte a las especies de manera diferente y tenga el potencial de generar cambios en la diversidad de hongos micorrízicos (Branco et al., 2022). Una mayor similitud en composición de HMA a nivel de género y morfoespecies fue registrada entre los suelos de las islas de vegetación y áreas abiertas del SAF milpa-chichipera y el suelo del matorral xerófilo (Cuadro 1), en donde morfoespecies de géneros como Acaulospora, Ambispora, Diversispora, Racocetra y Septoglomus, que sólo están presentes en uno o dos de los inóculos, podrían estar explicando la diferencia en la efectividad de los inóculos micorrízicos evaluados. La respuesta de la lechuga puede ser atribuida a las diferencias en la composición de HMA, debido a que se ha demostrado que las hortalizas como la lechuga albergan consorcios micorrízicos particulares en su rizosfera (Acoltzi-Conde et al., 2024). Asimismo, Wee et al. (2010), al evaluar efectividad de Glomus sp. en el desarrollo la lechuga, reportaron que este hongo incrementa el área foliar, peso fresco y contenido de P en las plantas inoculadas con respecto al control; aunque no la cantidad de clorofila, con valores que difieren marcadamente de los reportados en estudios que evalúan el efecto de otros HMA (Ley-Rivas et al., 2011; Ley-Rivas et al., 2016), lo cual sugiere que el desarrollo de la lechuga varía con la composición de especies del inóculo micorrízico.
El mayor contenido de clorofila en las hojas registrado en las plantas inoculadas con los HMA del matorral xerófilo y de las islas de vegetación con respecto a las inoculadas con los HMA del área abierta (Figura 2b), también pudiera deberse a las diferencias en la composición de HMA entre estos inóculos, ya que los HMA están sometidos a mayores entradas de nitrógeno al suelo a través de leguminosas nativas del matorral xerófilo y también presentes en las islas de vegetación del SAF (Moreno-Calles et al., 2012). Shi et al. (2016) reportaron que, tanto la actividad fotosintética como la concentración de clorofila y N de la planta hospedera, están correlacionadas y dependen de los HMA, donde A. scrobiculata, F. mosseae y R. intraradices favorecen su concentración en tejido vegetal comparado con plantas sin inocular. Asimismo, como ocurrió en el presente estudio, la efectividad de los HMA también se ha reflejado en la acumulación de clorofila, e incluso carotenoides y tocoferol en las hojas de la lechuga, lo que podría potenciar la ingesta de compuestos con propiedades antioxidantes durante el consumo de lechuga en la dieta (Baslam, Esteban, García y Goicoechea, 2013).
Las plantas de lechuga que crecieron con los inóculos del SAF más similares en composición de HMA tuvieron mejor nutrición y mayor actividad fotosintética, lo que se reflejó en más crecimiento en términos de biomasa (Figura 4). Cuenca et al. (2007) indicaron que algunos HMA tienen una alta compatibilidad micorrízica que favorece el crecimiento de la lechuga. Asimismo, la efectividad individual y en consorcio de los HMA aumenta la biomasa aérea y subterránea, así como el diámetro del tallo de la lechuga (Ley-Rivas et al., 2016). Esto sugiere que la efectividad de los HMA en el SAF “milpa-chichipera” pudiera deberse a su acción como consorcios micorrízicos nativos con una particular composición de especies (Trejo et al., 2011), como se ha demostrado tanto en plantas cultivadas de chile (Carballar-Hernández et al., 2018) y frijol (Paredes-Jácome et al., 2022), como en plantas de ambientes semiáridos como el Agave (Quiñones-Aguilar, Montoya, Rincón, Lobit y López, 2016) y varias especies de Mimosa spp. (Camargo-Ricalde et al., 2010; Montaño-Arias et al., 2023), donde los consorcios de HMA nativos incrementan la cantidad de hojas, altura y biomasa fresca y seca de las plantas hospederas.
Figura 4: Plantas de lechuga (Lactuca sativa L.) creciendo con y sin inóculo de hongos micorrízicos arbusculares (HMA) provenientes de áreas abiertas e islas de vegetación del sistema agroforestal (SAF) “milpa-chichipera” y matorral xerófilo del Valle semiárido de Tehuacán-Cuicatlán, Puebla-Oaxaca, México.

En este sentido, se ha registrado que, en otros SAF, los consorcios de HMA nativos estimulan el crecimiento vegetal en comparación con plantas no inoculadas y que, mientras más especies conformen el consorcio, mayor es la supervivencia de las plantas (Trejo et al., 2011). En este trabajo los dos inóculos provenientes del SAF “milpa-chichipera” (islas de vegetación y áreas abiertas) tuvieron mayor riqueza y similar composición de morfoespecies de HMA, lo que los hace fuentes de inóculos efectivos que favorecen el crecimiento de las hojas y de la planta, así como la nutrición y acumulación de agua en la parte aérea de las plantas de lechuga (Figuras 1, 2), características vinculadas a la calidad de esta hortaliza (Medina-Lozano et al., 2021). Asimismo, la efectividad de los inoculantes micorrízicos nativos en consorcios, no sólo favorecen la respuesta positiva de sus hospederos, en comparación con los inoculantes de HMA constituidos por una sola especie, sino que, en conjunto, también contribuyen a disminuir la aplicación de fertilizantes químicos hasta en un 50%, debido a que también son eficientes en la absorción de N y P, lo que aumenta el rendimiento de los cultivos, como en este caso con la lechuga (Miransari, 2011; Thioub, Ewusi, Sarkodie y Adjei-Gyapong, 2019; Aguégué et al., 2021). No obstante, la efectividad no sólo depende de los HMA, la planta hospedera y la fertilidad química del suelo, sino también está regulada por las características del suelo o el sustrato (Tapia-Goné et al., 2010; Jiménez-Martínez, González-Chávez, Gutiérrez-Castorena, Lara-Hernandez y García-Cué, 2014), por lo que la efectividad de los inóculos micorrízicos evaluados en este estudio podría variar in situ en función de las características físicas del suelo y, entonces, se requiere probar su efectividad en condiciones de campo.
En el contexto del SAF “milpa-chichipera”, los HMA podrían estar interconectando, a través de sus redes miceliares, a los cultivos (la milpa) que crecen en las áreas abiertas con las plantas nativas en las islas de vegetación, favoreciendo el crecimiento y nutrición de los cultivos (Marney y Borden, 2019; Gutiérrez-Núñez y Gavito, 2024), lo cual también podría ocurrir al cultivar hortalizas como la lechuga por lo que es una hipótesis por demostrar.
Conclusiones
Los inóculos de HMA del matorral xerófilo y SAF tradicional “milpa-chichipera” tienen impacto positivo en el crecimiento y contenido de N y P de las plantas de lechuga, por lo que son potenciales fuentes de biofertilizantes nativos multi-específicos que posiblemente conforman consorcios que favorecen la producción de esta hortaliza. Los inóculos micorrízicos arbusculares de las áreas abiertas y de las islas de vegetación del SAF “milpa-chichipera” fueron los más efectivos en la producción de lechuga, lo que revela que estos hongos podrían estar mejor adaptados a condiciones de mayor estrés ambiental que los HMA del matorral xerófilo, por lo que son más eficientes en los cultivos de este SAF tradicional. Este estudio sugiere que el SAF “milpa-chichipera” es un reservorio de inóculos de HMA potencialmente útiles en la agricultura dentro del Valle semiárido de Tehuacán-Cuicatlán, Puebla-Oaxaca, México, cuya efectividad deberá probarse en otras plantas cultivadas como el maíz o el frijol, o bien en algunas de interés para los pobladores como son el propio chichipe y los agaves, esto tanto en condiciones de invernadero como de campo, en donde se involucre a los agricultores para que generen a futuro sus propios biofertilizantes micorrízicos nativos.

