











 text new page (beta)
text new page (beta) Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
Permalink
Introducción
En ecosistemas áridos y semiáridos, donde los recursos son escasos, dispersos y sujetos a una fuerte estacionalidad, el agua superficial constituye un recurso ecológico crítico cuya disponibilidad influye en la distribución, el comportamiento y las interacciones de los vertebrados terrestres (Cain et al., 2006; Atwood et al., 2011; Harris et al., 2015). En particular, los aguajes permanentes representan lugares donde se concentra la presencia y actividad de la fauna, lo que incrementa la probabilidad de encuentros entre especies con requerimientos hídricos similares (Atwood et al., 2011; Hall et al., 2013). Esta concentración espacial puede intensificar las interacciones competitivas y de depredación dentro de gremios como el de los carnívoros (Palomares & Caro, 1999; Fedriani et al., 2000; Donadio & Buskirk, 2006; Hunter & Caro, 2008). Interacciones interespecíficas como la competencia por interferencia y los desplazamientos por depredación intragremial pueden alterar tanto el uso espacial como los patrones temporales de actividad de los carnívoros, como se ha observado en otros sistemas semiáridos (Krag et al., 2023; Morin et al., 2024).
Los mecanismos de partición del nicho, en particular la segregación espacial y la partición temporal, resultan esenciales para facilitar la coexistencia (Schoener, 1974; Kronfeld-Schor & Dayan, 2003; Monterroso et al., 2014). Mientras que en regiones más húmedas la segregación espacial puede bastar para evitar conflictos, en zonas donde los puntos de agua son escasos y predecibles, el espacio disponible para evitar el encuentro entre especies puede estar limitado. En estos casos, el tiempo se vuelve una dimensión ecológica clave cuya partición permite reducir encuentros directos y minimizar el riesgo de competencia o depredación (Kronfeld-Schor & Dayan, 2003; Di Bitetti et al., 2009; Edwards et al., 2015).
En los desiertos subtropicales, donde la estacionalidad marca fuertes contrastes entre la temporada seca y la de lluvias, se espera que las dinámicas de competencia y estrategias de evitación se intensifiquen durante los meses más secos (Harris et al., 2015; Ochoa et al., 2021). En estas condiciones, la demanda fisiológica de agua y la distribución de presas pueden modificar el comportamiento de carnívoros y alterar sus patrones de uso del espacio y el tiempo (Hall et al., 2013). Sin embargo, existe poca información sobre cómo las comunidades de carnívoros medianos y pequeños estructuran su coexistencia en torno a aguajes naturales en ambientes áridos y cómo estos patrones responden a la variación estacional.
Estudios recientes han mostrado que carnívoros simpátricos pueden coexistir mediante ajustes finos en sus patrones de actividad, incluso cuando comparten hábitat o recursos (Di Bitetti et al., 2009; Monterroso et al., 2014; Branney et al., 2023). Los mesocarnívoros cumplen roles ecológicos clave como depredadores, competidores y dispersores, y suelen ser naturalmente más abundantes que los grandes carnívoros. En regiones donde estos últimos han sido extirpados o son escasos, los mesocarnívoros desempeñan un papel aún más destacado en la estructuración de las comunidades (Roemer et al., 2009), lo que hace relevante entender cómo se organiza su coexistencia en torno a recursos limitados. En estos contextos, la segregación temporal contribuye a reducir la competencia y el riesgo de encuentros agresivos, en particular cuando las especies son morfológicamente similares y comparten hábitos tróficos (Marinho et al., 2020). De forma adicional, las diferencias en tamaño corporal entre especies pueden modular la intensidad de la competencia, la tolerancia a encuentros directos y el riesgo de interferencia o depredación. Diversos estudios han propuesto que estas diferencias de tamaño influyen en el grado de solapamiento horario y en la coocurrencia espacial entre especies (Donadio & Buskirk, 2006; Atwood et al., 2011; Ochoa et al., 2021), actuando como un eje de partición adicional que refuerza los mecanismos de coexistencia.
En este estudio evaluamos el uso espacial y temporal de aguajes por parte de cinco especies de carnívoros (coyote, Canis latrans Say, 1823; lince, Lynx rufus Schreber, 1777; zorra gris, Urocyon cinereoargentus Schreber, 1775; zorrillo, Spilogale gracilis Merriam, 1890; mapache, Procyon lotor Linnaeus, 1758), en un ecosistema semiárido del noroeste de México, caracterizado por la ausencia de flujos de agua permanentes. Nuestro objetivo fue analizar los patrones de uso de aguajes permanentes por parte de cinco especies de mesocarnívoros en un ecosistema árido, evaluando la participación de mecanismos de partición temporal y espacial como vía para su coexistencia. Planteamos dos hipótesis generales: (i) La partición temporal y la segregación espacial funcionan como mecanismos de coexistencia en torno a recursos hídricos compartidos. Bajo esta hipótesis, esperamos que las especies eviten el uso simultáneo de los aguajes, ya sea distribuyéndose en el tiempo o utilizando sitios distintos, lo que se reflejará en patrones diferenciados de actividad diaria y en una coocurrencia espacial menor a la esperada por azar (Hall et al., 2013; Harris et al., 2015; Lesmeister et al., 2015). (ii) La estacionalidad y la diferencia de tamaño corporal influyen en la intensidad de dicha partición. Durante la estación seca, el acceso al agua se restringe y la presión de uso sobre los aguajes se intensifica, lo que podría reforzar los mecanismos de partición para evitar encuentros directos. Asimismo, las diferencias de tamaño entre especies podrían reflejar asimetrías en dominancia o tolerancia al riesgo, facilitando una partición del recurso basada en relaciones de subordinación o estrategias de evitación (Atwood et al., 2011). De acuerdo con ello, esperamos, (a) que la partición temporal entre especies se intensifique durante la temporada seca, y (b) que la diferencia de tamaño corporal se relacione negativamente con el solapamiento horario y con la coocurrencia espacial entre especies.
Materiales y métodos
Área de estudio. El estudio se realizó en el piedemonte de la Sierra Cacachilas, dentro de la Región del Cabo, Baja California Sur, aproximadamente a 25 km al sur de la ciudad de La Paz (24.071° N, -110.054° W). La altitud en el área varía entre 120 y 1,053 m y se ubica en la transición entre el Desierto Sonorense y la Región Tropical del Cabo. La vegetación dominante varía con la altitud, en las zonas bajas (< 500 m) predomina el matorral tropical, mientras que las partes más elevadas (> 500 m) están ocupadas por selva seca (González-Abraham et al., 2010) (Fig. 1). Según datos de la estación climatológica de Los Robles, ubicada en las cercanías (24.036° N, -110.125° W), la precipitación anual promedio es de 322 mm, la temperatura mensual promedio es de 22.1° C, y las temperaturas mínima y máxima promedio son de 15.1° C y 29.1° C, respectivamente (CONAGUA, 2025). En el área de estudio no existen flujos de agua permanentes; el agua superficial está disponible exclusivamente en forma de aguajes permanentes y, durante la temporada de lluvias, también en aguajes efímeros y arroyos temporales.
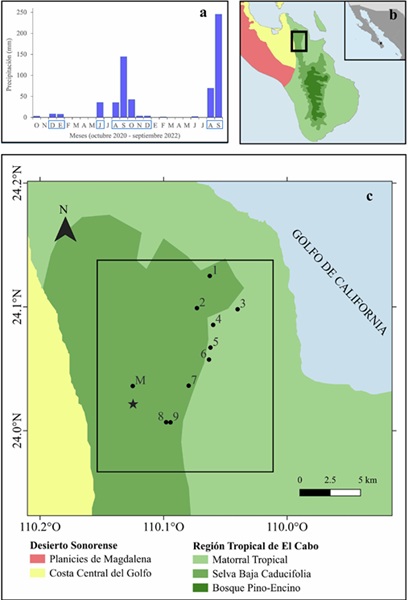
Figura 1 Localización del área de estudio en la Región del Cabo, Baja California Sur, México. (a) Precipitación mensual registrada en 11 pluviómetros instalados en el área de estudio entre octubre de 2020 y septiembre de 2022; los meses enmarcados indican el periodo de lluvias. (b) Ubicación regional del área de estudio. (c) Distribución de los nueve aguajes monitoreados con cámaras trampa (círculos, 1-9) y localización de la estación meteorológica utilizada para estimar la precipitación histórica (estrella, M).
Obtención de datos. Se seleccionaron nueve aguajes permanentes entre los disponibles en el área de estudio, procurando una representación amplia. La distancia mínima entre pares de aguajes varió entre 0.47 y 3.6 km, y la distancia entre los extremos del conjunto fue de 14 km. Se utilizaron datos de nueve cámaras trampa Bushnell (Trophy Cam E3 Essential Trail Camera) con sensor infrarrojo y un alcance de 30.5 m. Las cámaras se colocaron entre 50 y 150 cm de altura, a una distancia de 2 a 5 m del aguaje. El muestreo abarcó dos años completos, de octubre de 2020 a septiembre de 2022 (730 días). Las cámaras estuvieron activas las 24 horas, se revisaron mensualmente y se registraron las especies de mamíferos fotografiadas, junto con fecha y hora. Se definió noche-trampa como el periodo de 24 horas entre las 12:00 h de un día y las 12:00 h del día siguiente. De manera paralela, se instalaron 11 pluviómetros para registrar la precipitación local. Se definieron como meses de lluvia aquellos con 3 mm ó más de precipitación acumulada y como meses de seca, los que registraron menos de 3 mm. Durante el primer año (octubre 2020 - septiembre 2021) se registraron 235 mm de precipitación y 367 mm en el segundo (octubre 2021 - septiembre 2022) (Fig. 1a). En septiembre de 2022, el huracán Kay provocó una saturación en algunos pluviómetros, por lo que los 246 mm registrados en este mes representan una subestimación de la precipitación ocurrida.
No todos los aguajes contaron con cámaras funcionales de manera continua durante el periodo de estudio. Algunas cámaras permanecieron inactivas por meses completos, lo que generó una variación en el esfuerzo de muestreo con entre 6 y 9 cámaras activas por noche. Esta variación fue incorporada explícitamente en los análisis. Se acumularon 5,931 noches trampa: 3,423 en meses de seca y 2,508 en meses con lluvia (Cuadro 1). Se consideró como un registro la aparición de una especie en una foto, independientemente del número de individuos presentes. Fotografías sucesivas de la misma especie en un mismo sitio se contaron como registros distintos únicamente si estuvieron separadas por al menos 30 minutos (Atwood et al., 2011; Hall et al., 2013). Bajo estos criterios se obtuvieron 4,060 registros de especies, 2,762 en meses de seca y 1,298 en meses de lluvia (Cuadro 1). Estos datos fueron utilizados en todos los análisis del estudio. El peso promedio de referencia considerado para cada especie fue: coyote, 11.5 kg (Bekoff, 1977); lince, 8.2 kg (Larivière & Walton, 1997); mapache, 6.3 kg (Lotze & Anderson, 1979); zorra, 5.0 kg (Fritzell & Haroldson, 1982); y zorrillo, 2.7 kg (Verts & Carraway, 2001).
Cuadro 1 Número de registros de las cinco especies de mesocarnívoros en cada estación del año y esfuerzo de muestreo realizado (número de noches-trampa) en la Región del Cabo, Baja California Sur, México. El periodo de muestreo abarcó dos años (730 noches), con 424 noches en la estación de secas y 306 noches en la de lluvias.
| Registros de especies | Secas | Lluvias | Total |
| Coyote | 521 | 122 | 643 |
| Lince | 99 | 36 | 135 |
| Mapache | 668 | 593 | 1,261 |
| Zorra | 1,291 | 423 | 1,714 |
| Zorrillo | 183 | 124 | 307 |
| Total registros | 2,762 | 1,298 | 4,060 |
| Noches trampa | 3,423 | 2,508 | 5,931 |
Análisis de datos
Frecuencia de visita. Se estimó la tasa promedio de visitas diarias por especie a los aguajes en meses de seca y de lluvias. Para cada especie y cada noche i, se calculó la tasa de visitas (T i ) como T i = V i /C i , donde V i es el número de visitas de la especie registradas esa noche (incluyendo noches con cero visitas), y C i es el número de sitios con cámaras activas esa misma noche. Posteriormente, se calculó el promedio de T i para las 424 noches correspondientes a la estación seca y las 306 noches de la estación de lluvias. Utilizamos modelos lineales (con errores asumidos normales) para comparar, en cada especie, la distribución de frecuencias de visita diaria entre estaciones y evaluamos la normalidad de los residuales mediante la prueba de Shapiro-Wilk (Midway & White, 2025). Dado que en ninguna especie los residuales cumplieron con el supuesto de normalidad (p < 0.05), utilizamos modelos lineales generalizados con distribución quasi-Poisson y función de enlace logarítmica, que permite incluir valores de frecuencia igual a cero y ofrece un comportamiento robusto ante la sobredispersión en los datos.
Solapamiento temporal. Se analizó inicialmente la distribución de la actividad horaria de las especies considerando el total del periodo de muestreo (730 noches) y por estación (424 noches en temporada seca y 306 en la de lluvia). Posteriormente, se evaluó (i) el solapamiento anual entre las cinco especies, (ii) el solapamiento entre estaciones para cada especie (seca vs. lluvia), y (iii) el cambio del solapamiento entre pares de especies en secas frente a lluvias.
La densidad de actividad horaria se estimó con la función overlapEst del paquete overlap 0.3.9 (Meredith et al., 2024) en R (R-Core Team, 2024), mediante estimación de densidad kernel (KDE; Ridout & Linkie, 2009). Para corregir la variación en el esfuerzo de muestreo entre noches, derivada de que algunas cámaras permanecieron inactivas durante meses completos, se asignó a cada registro un peso proporcional al inverso del número de sitios (cámaras) activos en la noche correspondiente. Específicamente, el peso asignado a cada registro i fue w i = 1/C i , donde C i representa el número de cámaras activas en la noche en que ocurrió el registro. Estos pesos fueron posteriormente normalizados, multiplicando cada peso (w i ) por N / (w i , para que su suma fuera igual al número total de registros originales (N). Esta ponderación evita que las noches con menor cobertura influyan de manera desproporcionada en la estimación de la densidad. Los valores ponderados fueron usados para generar las curvas de actividad horaria y evaluar el solapamiento temporal entre especies. Para propósitos de visualización, las curvas de densidad se graficaron con la función density de R (R-Core Team, 2024), usando un ancho de banda fijo de 1 y con el eje horario ajustado a un ciclo de 12h-12h.
El solapamiento entre las curvas de actividad, tanto anuales como estacionales, se cuantificó con el coeficiente ∆4, cuando ambas muestras superaron los 75 registros, y mediante ∆1 cuando al menos una de ellas tuvo menos de 75 registros (Meredith et al., 2024). Ambos coeficientes estiman el área de superposición entre las dos curvas de actividad. La incertidumbre asociada se estimó con 1,000 repeticiones bootstrap, y se calcularon intervalos de confianza del 95% usando los percentiles 2.5 y 97.5 (Meredith et al., 2024). Para analizar el cambio estacional en el solapamiento de cada par de especies, se estimó la diferencia en el coeficiente de solapamiento (∆ i ) del par de especies entre estaciones como ∆ i _lluvia − ∆ i _seca. La incertidumbre asociada a esta diferencia se evaluó mediante un procedimiento de muestreo bootstrap (1,000 iteraciones), a partir del cual se calculó el intervalo de confianza del 95%. Este mismo conjunto de réplicas se utilizó para estimar la significancia estadística. En cada réplica, se recalculó la diferencia entre el solapamiento en lluvias y en secas. La significancia estadística se evaluó mediante un enfoque bidireccional, calculando la proporción (p) de réplicas en las que el valor absoluto de la diferencia simulada fue igual o superior al valor absoluto de la diferencia observada. La diferencia se consideró estadísticamente significativa si p < 0.05 (Legendre & Legendre, 2012; Manly, 2018).
Con el objeto de evaluar si la diferencia de tamaño corporal entre especies influye en el grado de solapamiento temporal de su actividad, analizamos la relación entre la diferencia de peso (kg) y el coeficiente de solapamiento horario (∆ i ) para cada uno de los 10 pares posibles entre las cinco especies estudiadas. El análisis se realizó por separado para la estación seca, la estación lluviosa y el conjunto anual. Dado el reducido tamaño de muestra (10 pares), exploramos la significancia estadística de esta relación mediante una prueba de permutación de la pendiente de la regresión lineal. Para cada periodo, se mantuvieron fijos los valores de solapamiento y se reordenaron aleatoriamente las diferencias de peso en 10,000 permutaciones, calculando la pendiente de la regresión lineal en cada una. La p empírica unidireccional se estimó como la proporción de permutaciones en que la pendiente fue igual o más negativa que la observada, bajo la hipótesis de que el solapamiento disminuye al aumentar la diferencia de tamaño corporal (Legendre & Legendre, 2012; Manly, 2018).
Segregación espacial. Para analizar si existía evitación espacial entre pares de especies, se seleccionaron únicamente las noches en que ambas estuvieron activas y se contabilizaron los casos en que coincidieron en un mismo sitio. Para evaluar si el grado de coocurrencia espacial observado difiere de lo esperado bajo un escenario de independencia espacial entre las especies, se aplicó un modelo nulo basado en permutaciones. Este modelo mantiene constante, para cada noche, tanto el número de sitios visitados por cada especie como el número de sitios con cámaras activas. Las visitas se asignaron de forma equiprobable entre los sitios activos en cada noche. Este procedimiento se repitió 1,000 veces, y el valor esperado de coocurrencia se estimó como el promedio del número de sitios compartidos entre especies en las simulaciones. La probabilidad empírica se calculó como la proporción de permutaciones en las que el número de coincidencias fue igual o superior al valor observado (Legendre & Legendre, 2012; Manly, 2018). El análisis se aplicó tanto al periodo completo (730 noches) como a cada estación (424 noches en secas y 306 noches en lluvias). El modelo nulo corresponde conceptualmente a un enfoque “suma de filas fijada - columnas equiprobable”, según la clasificación de Gotelli (2000).
A partir de las mismas permutaciones utilizadas para estimar la significancia de la coocurrencia espacial, se calculó el estadístico z, definido como la distancia (en desviaciones estándar) entre el valor observado y el esperado bajo el modelo nulo. Este estadístico indica si la asociación espacial entre dos especies es positiva (z > 0) o negativa (z < 0) y su magnitud muestra la intensidad de dicha asociación. Este parámetro se utilizó para evaluar si la diferencia de peso entre pares de especies se relaciona con su asociación espacial, aplicando una prueba de permutaciones de la pendiente de regresión (10,000 permutaciones), tanto para el periodo anual como por estación.
Finalmente, de forma análoga, se analizó mediante una prueba de permutaciones de la pendiente de regresión si el grado de solapamiento en la actividad temporal (∆ i ), como variable independiente, se relaciona con la intensidad de la asociación espacial entre especies (z), para cada periodo, anual y por estaciones.
Resultados
Frecuencia de visita diaria. La frecuencia de visitas a los aguajes fue significativamente menor durante la estación de lluvias que durante la de secas para el coyote (( = −1.16, EE = 0.11, z = − 10.10, p < 0.001), el lince (( = −0.76, EE = 0.20, z = − 3.79, p < 0.001) y la zorra (( = −0.81, EE = 0.08, z = − 9.59, p < 0.001). En términos de frecuencia diaria promedio, las visitas se redujeron de 0.15 a 0.05 en el coyote, de 0.03 a 0.01 en el lince y de 0.37 a 0.17 en la zorra (Fig. 2). Por el contrario, el mapache incrementó su frecuencia de visitas en lluvias (( = 0.19, EE = 0.06, z = 3.13, p = 0.0018), pasando su frecuencia promedio de 0.19 a 0.23. En el zorrillo no se observaron diferencias significativas entre estaciones (( = −0.16, EE = 0.15, z = − 1.07, p = 0.28), con frecuencias similares en ambas temporadas (0.05) (Fig. 2).
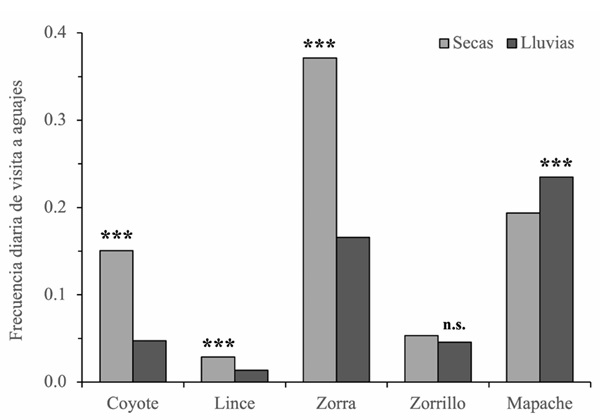
Figura 2 Frecuencia diaria promedio de visitas a aguajes por cinco especies de carnívoros (coyote, lince, zorra gris, zorrillo y mapache) en la Región del Cabo, Baja California Sur, durante las estaciones de seca y de lluvias. Las barras muestran los promedios por estación; las diferencias significativas entre estaciones se evaluaron mediante modelos lineales generalizados: (***) p < 0.001; (n.s.) diferencia no significativa.
Solapamiento temporal. A lo largo del año, todas las especies mostraron un patrón bimodal de actividad, aunque con diferencias en la distribución y amplitud de los picos horarios entre grupos funcionales. El coyote y el lince fueron más activos en horarios crepusculares (18:00-22:00 h) y matutinos (04:00-08:00 h), mientras que la zorra, el zorrillo y el mapache concentraron su actividad en horas nocturnas (20:00-23:00 h y 02:00-06:00 h) (Fig. 3). El solapamiento horario a lo largo de todo el periodo entre pares de especies varió entre ∆4 = 0.63 y ∆4 = 0.90, con intervalos de confianza al 95% de amplitud entre 0.04 y 0.15 (Fig. 4).
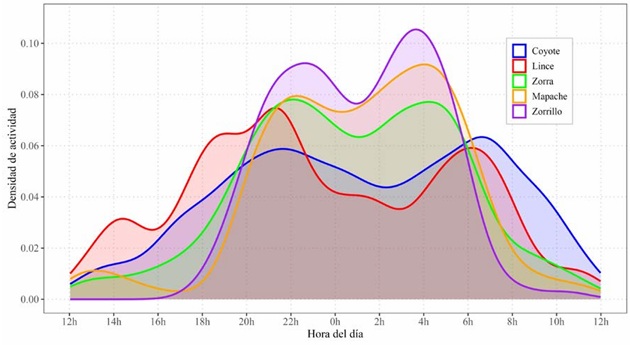
Figura 3 Distribución horaria de la actividad en aguajes permanentes de cinco especies de carnívoros (coyote, lince, zorra gris, zorrillo y mapache) en la Región del Cabo, Baja California Sur, México. Las curvas representan la densidad de actividad de cada especie normalizada a un área total igual a uno.
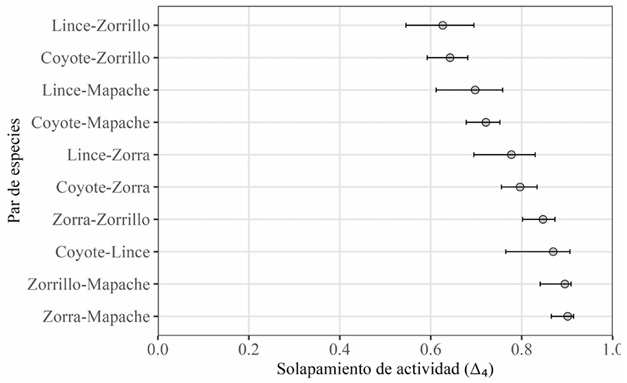
Figura 4 Solapamiento anual de la actividad horaria en la visita a aguajes permanentes entre pares de especies de carnívoros. Se muestran los valores del coeficiente de solapamiento observado (Δ₄, círculos) y los intervalos de confianza al 95% obtenidos mediante remuestreo bootstrap (líneas horizontales), bajo un escenario aleatorio.
El solapamiento de la actividad horaria entre estaciones fue similar para la mayoría de las especies, con valores de solapamiento entre 0.87 y 0.92: coyote (∆4 = 0.92, IC 95%: 0.81 - 0.93), lince (∆1 = 0.87, IC 95%: 0.72 - 0.92), zorra (∆4 = 0.89, IC 95%: 0.84 - 0.92) y zorrillo (∆4 = 0.87, IC 95%: 0.76 - 0.91). El mapache presentó el valor más bajo (∆4 = 0.78, IC 95%: 0.73 - 0.81). Las curvas de actividad horaria de mapache y zorrillo muestran un patrón más concentrado en la estación seca y más extendido durante los meses de lluvias, siendo más estables en las demás especies (Fig. 5 a, b). El solapamiento horario entre pares de especies aumentó, en la mayoría de los casos (7 de 10), en lluvias respecto a secas, sin embargo, las diferencias no fueron significativas en ningún caso (p empírica bidireccional > 0.05) (Cuadro 2).
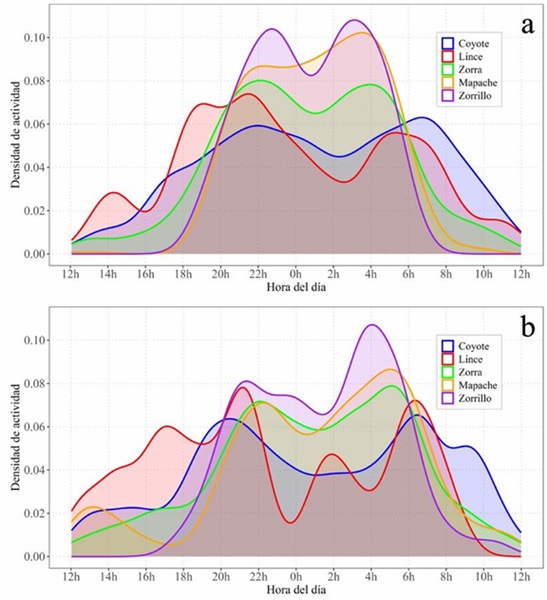
Figura 5 Distribución de la actividad horaria en la visita a aguajes permanentes por cinco especies de carnívoros (coyote, lince, zorra gris, zorrillo y mapache) en la Región del Cabo, Baja California Sur, México, durante la estación seca (a) y la estación lluviosa (b). Las curvas representan la densidad de actividad horaria de cada especie, normalizada a un área total igual a uno.
Cuadro 2 Solapamiento horario de la actividad (Δ i ) entre pares de especies de mesocarnívoros entre estación de lluvia y la estación de seca, y su diferencia observada (ΔL-ΔS)obs. El estimador fue (4, excepto en el caso del lince en lluvias, para el cual se utilizó (1 debido al bajo número de registros (36). La diferencia de solapamiento esperada, (ΔL-ΔS)esp, el intervalo de confianza del 95% y la p empírica bilateral se obtuvieron mediante remuestreo bootstrap (1,000 iteraciones).
| Especies | ∆i lluvia | ∆i seca | (∆L – ∆S)obs | (∆L– ∆S)esp | IC 95% dif. esp. | p |
| Coyote-Lince | 0.854 | 0.875 | –0.020 | –0.078 | (–0.207, 0.045) | 0.869 |
| Coyote-Zorra | 0.797 | 0.787 | 0.010 | –0.004 | (–0.085, 0.080) | 0.839 |
| Coyote-Zorrillo | 0.656 | 0.617 | 0.039 | 0.031 | (–0.071, 0.131) | 0.532 |
| Coyote-Mapache | 0.759 | 0.643 | 0.117 | 0.101 | (0.012, 0.186) | 0.367 |
| Lince-Zorra | 0.748 | 0.789 | –0.041 | –0.063 | (–0.215, 0.072) | 0.692 |
| Lince-Zorrillo | 0.621 | 0.614 | 0.007 | –0.014 | (–0.162, 0.139) | 0.919 |
| Lince-Mapache | 0.711 | 0.636 | 0.075 | 0.041 | (–0.101, 0.174) | 0.388 |
| Zorra-Zorrillo | 0.842 | 0.824 | 0.018 | –0.001 | (–0.075, 0.072) | 0.630 |
| Zorra-Mapache | 0.908 | 0.849 | 0.059 | 0.040 | (–0.008, 0.085) | 0.230 |
| Zorrillo-Mapache | 0.833 | 0.942 | –0.109 | –0.105 | (–0.176, –0.035) | 0.456 |
Segregación espacial. La coocurrencia en un mismo aguaje durante una misma noche fue menor a la esperada bajo un modelo de independencia en todos los pares analizados (z < 0). Esta evitación espacial se detectó tanto en el periodo anual como en cada estación, y fue estadísticamente significativa en todos los casos (p empírica unidireccional = 1). La magnitud de la evitación espacial tendió a ser mayor durante la temporada de secas que en la de lluvias.
Diferencia de tamaño. El solapamiento temporal entre pares de especies a lo largo de todo el año disminuyó significativamente a medida que aumentó la diferencia de peso entre ellas, tanto a lo largo del año (pendiente = −0.023; p = 0.037) como durante la estación de lluvias (pendiente = −0.024; p = 0.036), aunque no en la estación seca (pendiente = −0.025; p = 0.080) (Fig. 6 a-c). En contraste, no se detectó una relación significativa entre la diferencia de peso y la coocurrencia espacial en ningún periodo (año: pendiente = −0.129, p = 0.435; lluvias: pendiente = 0.191, p = 0.667; secas: pendiente = −0.251, p = 0.319) (Fig. 6 d-f). Finalmente, el solapamiento temporal se asoció negativamente con la coocurrencia espacial en la estación de lluvias (pendiente = −19.473; p empírica = 0.027), pero no en el año completo (pendiente = −16.198; p = 0.136) ni en la estación seca (pendiente = −4.417; p = 0.340) (Fig. 6 g-i).
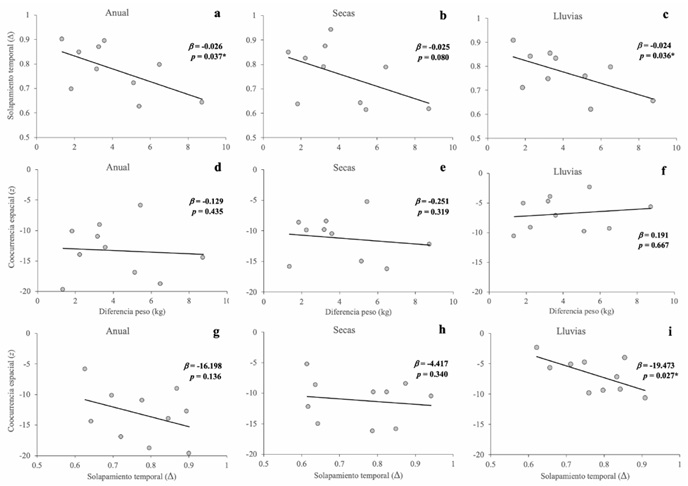
Figura 6 Relación entre diferencia de peso y solapamiento temporal (a-c), diferencia de peso y coocurrencia espacial (d-f) y entre solapamiento temporal y coocurrencia espacial (g-i) entre pares de especies de carnívoros. Las columnas muestran los resultados para el periodo anual (izquierda), estación seca (centro) y estación de lluvias (derecha). Los puntos representan los valores observados para los pares de especies y la línea indica únicamente la tendencia visual de la relación. (( corresponde a la pendiente de regresión observada y p es la probabilidad empírica estimada mediante una prueba de permutaciones (10,000 iteraciones). Se indica con asterisco (*) cuando p < 0.05.
Discusión
En este estudio analizamos cómo cinco especies de mesocarnívoros utilizan aguajes permanentes en un ecosistema árido del noroeste de México, evaluando el papel de la partición espacio-temporal en su coexistencia. Nuestros resultados respaldan la hipótesis de que el uso simultáneo de estos sitios se produce mediante mecanismos complementarios: todas las especies mostraron una clara evitación espacial y patrones diferenciados de actividad horaria, con una segregación funcional entre especies crepusculares y nocturnas. En contraste, los efectos de la estacionalidad y del tamaño corporal fueron más heterogéneos. El solapamiento temporal no varió significativamente entre estaciones. Sin embargo, en la estación de lluvias, se observó una asociación positiva entre solapamiento temporal y evitación espacial, relación que no se detectó en secas. La diferencia de tamaño corporal se asoció negativamente con el solapamiento temporal sólo en lluvias, sin relación con la evitación espacial en ningún periodo.
Durante la temporada seca, el coyote, el lince y la zorra aumentaron su frecuencia de visitas, lo que sugiere una mayor dependencia del agua libre cuando su disponibilidad es limitada, en línea con estudios previos en ambientes áridos (Cain et al., 2006; Hall et al., 2013; Harris et al., 2015). En contraste, el zorrillo mantuvo y el mapache incluso aumentó la frecuencia de visita en lluvias, lo que sugiere diferencias fisiológicas o conductuales en el acceso al agua. El zorrillo, por su pequeño tamaño, podría obtener una mayor proporción de agua a través de sus presas (Gittleman & Harvey, 1982; McNab, 2002). En cambio, especies de mayor tamaño como el coyote, el lince o la zorra tienen requerimientos absolutos más altos, difíciles de cubrir con agua metabólica, lo que incrementa su dependencia del agua libre (Golightly & Ohmart, 1984). En el caso del mapache, el aumento en la frecuencia de visita durante la temporada de lluvias podría deberse a que los aguajes no sólo le proporcionan agua, sino que también constituyen un componente clave del hábitat donde obtiene alimento (Heske & Ahlers, 2016), cuya disponibilidad podría aumentar en esta estación.
En términos temporales, las especies mostraron patrones de actividad diferenciados entre sí, pero consistentes y relativamente estables entre estaciones. El coyote y el lince fueron más crepusculares, mientras que la zorra, el zorrillo y el mapache concentraron su actividad en horas nocturnas, un patrón también reportado en otros estudios (George & Crooks, 2006; Atwood et al., 2011). Esta organización horaria puede reflejar tanto diferencias funcionales como presiones de competencia o riesgos de agresión o predación (Kronfeld-Schor & Dayan, 2003). La mayor similitud en los patrones de actividad entre especies del mismo grupo funcional (crepusculares o nocturnos) se reflejaría en el mayor solapamiento temporal de especies dentro de cada grupo y menor entre ellos, lo que sugiere cierto grado de partición temporal del recurso (Di Bitetti et al., 2009).
Contrario a lo esperado y a lo observado en otros estudios (Palomares & Caro, 1999; Vanak et al., 2013), no se encontró un incremento significativo en la partición temporal del uso de aguajes durante la temporada seca, pese a la mayor limitación del recurso hídrico. En cambio, la diferencia de tamaño corporal mostró una relación negativa con el solapamiento temporal durante la temporada de lluvias, cuando el recurso es más abundante espacialmente. La modulación de la actividad horaria en función de diferencias de tamaño, especialmente cuando implica asimetrías de dominancia o riesgo, ha sido documentada en diversos estudios (Vanak et al., 2013; Xue et al., 2018; Vernes et al., 2022). En este caso, sin embargo, la asociación apareció sólo en temporada de lluvias. Esto sugiere que, en condiciones menos restrictivas, la diferenciación en el uso temporal de los aguajes, parcialmente condicionada por la diferencia de tamaño entre especies, puede ser suficiente para permitir su uso compartido. En ecosistemas secos similares, como la caatinga brasileña, se ha observado que incluso sin una segregación marcada, una separación en los picos de actividad puede ser suficiente para disminuir la probabilidad de encuentros agonísticos y facilitar la coexistencia (Marinho et al., 2020). De forma similar, Smith et al. (2023) proponen que la evitación interespecífica no requiere una separación total en el ciclo diario, sino que puede manifestarse como una asincronía sutil en los periodos de actividad núcleo, lo que permite cierto solapamiento general con mecanismos finos de partición temporal.
A nivel espacial, los resultados muestran que, cuando dos especies están activas la misma noche, coinciden en el mismo aguaje con menor frecuencia que la esperada bajo un uso independiente. Este patrón sugiere una modulación en la presencia de las especies para evitar encuentros directos, posiblemente como estrategia para reducir conflictos y prevenir agresión o depredación intra-gremial (Palomares & Caro, 1999; Donadio & Buskirk, 2006). La persistencia de esta evitación a lo largo del año resalta su papel como estrategia espacial consistente, más allá de las variaciones estacionales (Marinho et al., 2020). En estudios similares, se ha observado que las especies más pequeñas tienden a evitar espacialmente a carnívoros más grandes y potencialmente dominantes, lo que indica que la diferencia de tamaño corporal puede promover la segregación espacial como mecanismo de evitación interespecífica (de Satgé et al., 2017). En nuestro caso, sin embargo, no se encontró relación entre la diferencia de tamaño y la intensidad de la evitación. Esta ausencia de relación podría indicar que las asimetrías de dominancia asociadas al tamaño corporal son débiles o no generan interacciones agresivas directas. De ser así, podrían modular la partición temporal, pero no influir claramente en la segregación espacial. No obstante, más de la mitad de los pares de especies presentan diferencias de tamaño intermedias según la clasificación de Donadio y Buskirk (2006), quienes señalan que estos pares son los más propensos a involucrarse en interacciones letales entre carnívoros.
Como último patrón relevante, encontramos que el solapamiento horario entre especies se relacionó positivamente con la intensidad de la evitación espacial, pero sólo durante la temporada de lluvias. En esta estación, también observamos que una mayor diferencia de tamaño se asociaba con menor solapamiento temporal. Esto podría indicar que, cuando la diferenciación temporal es insuficiente durante lluvias, las especies pueden recurrir a la segregación espacial para reducir el riesgo de encuentros. La ausencia de estas relaciones en secas no implica necesariamente una menor presión por evitar encuentros, sino que podría reflejar una limitación física impuesta por la concentración del recurso en pocos aguajes, lo que reduce las posibilidades de espaciamiento o alternancia temporal. Además, la resolución espacial y el tamaño reducido de muestra de este estudio podrían limitar nuestra capacidad para detectar con precisión estas dinámicas, o introducir sesgos relacionados con una posible preferencia o evitación de los aguajes por factores no relacionados con la oferta de agua, como la disponibilidad de alimento o la competencia trófica (Neale & Sacks, 2001; Arjo et al., 2007).
En conjunto, nuestros resultados muestran que, en este ecosistema árido, los mesocarnívoros reducen la coincidencia en el uso de aguajes mediante una combinación de segregación horaria y evitación espacial. Esta organización fue consistente entre estaciones, aunque con diferencias en su expresión. Durante la temporada de lluvias, la diferencia de tamaño corporal se asoció con menor solapamiento horario y los pares de especies con mayor solapamiento temporal mostraron mayor evitación espacial, lo que sugiere una compensación entre ambos ejes de partición. En cambio, estos patrones no se observaron en la temporada seca, posiblemente debido a la contracción en el número de aguajes disponibles. Estos resultados, obtenidos a partir de datos en un número limitado de sitios, deben interpretarse como evidencia exploratoria sobre la estructuración espacio-temporal de la coexistencia en ambientes de alta estacionalidad hídrica.

