











 text new page (beta)
text new page (beta) Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
Permalink
Introducción
El género Fusarium Link 1809, pertenece a la clase Ascomycota y corresponde a la fase asexual de la familia Nectriaceae. Son hongos cosmopolitas (Nelson et al., 1994) saprobios, endófitos (Leslie y Summerell, 2006) o patógenos oportunistas provocando la mortalidad de insectos y plantas (Mosqueda-Anaya et al., 2018). Las especies del género producen cantidades importantes de metabolitos secundarios como las micotoxinas; entre las más estudiadas están los tricotecenos, las zearealenonas y las fumonisinas (Jonsson, 2017), las cuales son estrogénicas, inmunomoduladoras, citotoxicas y mutagénicas (Desjardins, 2006). También producen diversos pigmentos como las naftoquinonas, antraquinonas, azonaftoquinonas y carotenoides (Frandsen et al., 2006; Goertz et al., 2010). Algunos pigmentos han sido citados con actividad biológica antiparasitaria, anticancerígena, antifúngica y antibacteriana (López et al., 2011; Kwon et al., 2007).
Los pigmentos y micotoxinas, en su mayoría, son derivados de la ruta biosintética de los policétidos. En este género de hongos, las familias de genes relacionadas a su síntesis se encuentran distribuidos en casi todas las especies de Fusarium (Frandsen et al., 2006; Lebeau et al., 2017; Malz, 2005). La producción de pigmentos en medios de cultivo específicos es un carácter usado en la taxonomía del grupo junto con caracteres macro y microscópicos (Nelson et al., 1994), particularmente, la expresión de pigmentos no solo depende del genotipo de las especies, sino también de la composición del medio o hábitat, el pH, la temperatura y la radiación (Garcia-Cela et al., 2018).
El presente trabajo tiene por objetivo determinar las especies de Fusarium que puedan ser aisladas de un Hemíptero (Bemisia aff. tabaci) asociado a una cucurbitácea ruderal (Cucurbita okeechobeensis subsp. martinezii (L.H. Bailey) Walters & Deck.-Walt.) y el tipo de pigmento que estas producen.
Materiales y métodos
Recolecta de muestras
Se recolectaron insectos muertos encontrados en hojas y frutos de una cucurbitácea ruderal conocida localmente como “morchete” o “calabacín loco” (Cucurbita okeechobeensis subsp. martinezii), en dos sitios secundarios de bosque mesófilo de montaña perturbado en los municipios de Emiliano Zapata (cerca del beneficio Puerto Rico) y Coatepec (carretera Xalapa-Coatepec). Las colectas vegetales fueron herborizadas y depositadas en el herbario XAL del Instituto de Ecología A.C. Los insectos fueron colocados en viales de vidrio con tapa para ser trasladados al laboratorio, la identificación se realizó con las claves de Caballero (1994, 1996), EPPO (2004) y Evans (2007). La literatura ha reportado que Bemisia tabaci comprende un complejo de especies, los ejemplares se identificaron como Bemsia aff. tabaci (Brown et al., 1995; Perring, 2001; ISSG, 2006).
Aislamiento de hongos
Los insectos se desinfectaron con hipoclorito de sodio al 0.5 % por 10 minutos, posteriormente fueron lavados con una disolución de etanol-agua (7:3) por un minuto y finalmente con agua destilada estéril.
El aislamiento fúngico se realizó mediante la colocación directa de los insectos en cajas Petri de 5 cm de diámetro, con medio de cultivo agar papa dextrosa (PDA, Bioxon). Se incubaron a temperatura ambiente (25 °C), durante 14 días, con un ciclo de 12h luz/12h oscuridad. Posteriormente se realizaron aislamientos, de los micelios visualmente diferentes en cajas Petri de 5 cm de diámetro con medio PDA, y fueron incubados durante 5-7 días, bajo las mismas condiciones de luz, temperatura y humedad. Mediante aislamientos consecutivos, se obtuvieron únicamente dos cultivos fúngicos axénicos con características morfológicas y velocidad de crecimiento distintas.
Identificación taxonómica
La identificación de las especies fúngicas se basó en la literatura especializada (Booth, 1971; Nelson et al., 1983, 1994; Seifert, 1996; Leslie y Summerell, 2006). Se observó la morfología de las estructuras características del género como la forma y pigmentación del micelio, conidióforos, células conidiógenas, conidios y otras estructuras especializadas, con ayuda de un microscopio estereoscópico Stemi DV4 (Carl Zeiss) y un microscopio compuesto iLED (Carl Zeiss). La velocidad de crecimiento en medio de cultivo PDA, se determinó en 7 a 21 días de incubación a 25 °C, 12h luz/12h oscuridad.
Extracción de los pigmentos
La extracción de los pigmentos se realizó mediante maceración del micelio con acetato de etilo (grado reactivo, Baker), durante 7 días, a partir de cultivos estáticos de 30 días de crecimiento. El disolvente se eliminó a presión reducida con un rotavapor Büchi a 45 °C. Para la visualización de los pigmentos coloridos, por cromatograma en capa delgada (CCD) se utilizaron cromatofolios de gel de sílice G (F254), en aluminio (Merck) y mezcla hexano-acetato de etilo (grado reactivo) 8:2, como eluyente. Los reveladores utilizados fueron luz ultravioleta onda corta, vapores de yodo sublimado (I2) y óleum (H2SO4 /ácido acético 80%, 1:1). Se obtuvieron espectros de resonancia magnética de protón (1HNMR) de los extractos crudos en un espectrómetro de NMR Bruker Avance III HD a 500 MHz (BIOMIMIC-INECOL Xalapa, Ver), utilizando CDCl3 como disolvente.
Resultados
Taxonomía
Fusarium Sección Arthrosporiella
Fusarium avenaceum (Fr.) Sacc., Syll. fung. (Abellini) 4: 713 (1886)
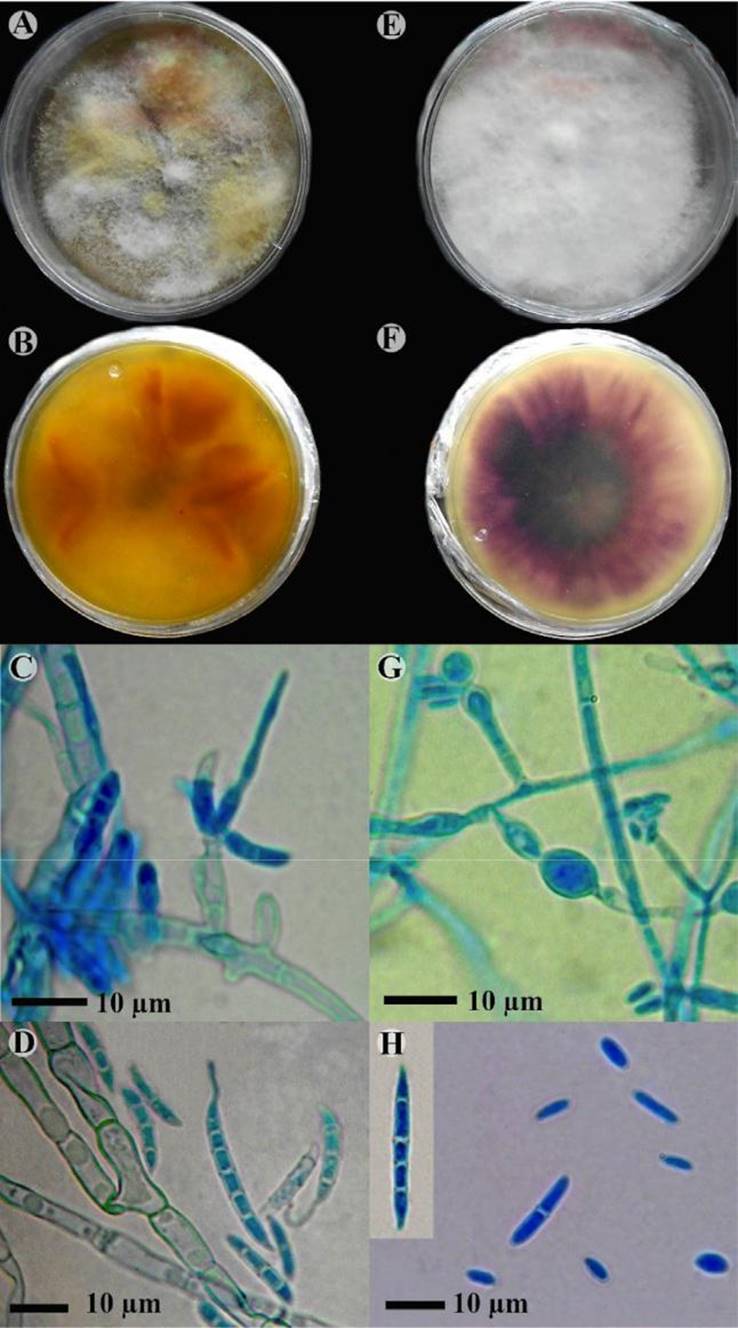
Figura 1. Especies de Fusarium aisladas de ejemplares muertos de Bemsia aff. tabaci. A-D. Fusarium avenaceum . A: micelio. B: reverso de la caja de Petri mostrando pigmentos cafés a anaranjados. C: conidióforos. D: macroconidios mostrando el apéndice largo característico de esta especie. E- H. Fusarium oxysporum. E: micelio. F: reverso de la caja de Petri mostrando los pigmentos violetas a morados. G: conidióforos y clamidosporas. H: macro y microconidios.
Morfología en PDA: micelio aéreo presente, de aspecto algodonoso, de color blanquecino con algunas zonas de color amarillento, masas de esporas (esporodoquios) de color café anaranjado que se observan como gútulas. Reverso de la caja de color café anaranjado, beige amarillento o beige anaranjado, en las orillas pigmentos concéntricos de color naranja. Con la edad, el pigmento se torna de color café oscuro. Conidióforos de (17.4-) 22.7- 33.8 µm de longitud, creciendo en esporodoquios, monofiálides no ramificadas alargadas. Macroconidios de (12-) 13-30.5 x (1.79) 2.5-3 µm, curvados dorsiventralmente, abundantes, con 0-5 septos, con pared delgada. Microconidios cilíndricos, hialinos, 5.85-9.7 × 1.9-2.3 µm, escasos. Clamidosporas ausentes. Velocidad de crecimiento en PDA 0.22 cm/24 h.
Hábitat: sobre cadáveres de Bemisia aff. tabaci en frutos y hojas de Cucurbita okeechobeensis sub. martinezii, bosque mesófilo de montaña secundario.
Material examinado: Veracruz. Municipio de Coatepec, carretera antigua Xalapa-Coatepec. Cerca del Retén (cepas CO2, CO2-1); municipio de Emiliano Zapata, cerca del Beneficio Puerto Rico (cepas CO2-2, CO2-3).
Notas: Las características morfológicas concuerdan con Fusarium avenaceum, observándose al reverso de la caja pigmentos de color café anaranjado, célula basal de los macroconidios alargada, a manera de un apéndice delgado o ganchudo, ausencia de clamidosporas y microconidios muy escasos (Booth, 1971; Nelson et al., 1983, 1994; Seifert, 1996; Leslie y Summerell, 2006).
Fusarium Sección Liseola
Fusarium oxysporum Schltdl., Fl. berol. (Berlin) 2: 139 (1824)
Morfología en PDA: micelio aéreo presente, de aspecto algodonoso y de color blanquecino. Reverso de la caja de color morado oscuro al centro, de color morado claro hacia las orillas de la caja o hialino. Con la edad, el pigmento originalmente de color violeta se torna violeta oscuro a casi negro.
Conidióforos de (5.9-) 8.4-11.9 x (2-) 2.2-4.5 µm de longitud, creciendo a manera de esporodoquios. Macroconidios de 11-30 x 2.5-3 µm, escasos, con 1-4, septos, con pared gruesa, célula basal en forma de pie o gancho, larga. Microconidios de 10-11 x 2-3 µm, abundantes, sin septos, ovoides a ligeramente reniformes, con pared gruesa. Clamidosporas. Presentes, elípticas, de pared gruesa, catenuladas, hialinas, de 8-9 x 6-7 µm. Velocidad de crecimiento en PDA 0.3 cm/24 hrs.
Hábitat: sobre cadáveres de Bemisia aff. tabaci. Sobre frutos y hojas de Cucurbita okeechobeensis sub. martinezii. Bosque mesófilo de montaña secundario.
Material examinado: Veracruz. Municipio de Coatepec, carretera antigua Xalapa-Coatepec. Cerca del Retén (cepas CO1, CO1-1); municipio de Emiliano Zapata, cerca del Beneficio Puerto Rico (cepas CO1-2, CO1-3).
Notas: Fusarium oxysporum es una especie de amplia distribución que se encuentra asociada a una diversidad de hospederos, en muchas ocasiones es fitopatógeno, Dentro de las características principales para su identificación se considera la capacidad de producir compuestos de color violeta o morado en el reverso de la colonia, y presentar macroconidios con un apéndice pequeño a manera de pie en la célula basal del macroconidio, característica distintiva de esta especie.
Observación de los pigmentos
Cada una de las cepas produjo coloraciones distintas observadas en medio PDA, sin embargo, en la cromatografía en capa delgada de las extracciones con acetato de etilo se observó que comparten al menos dos componentes iguales, Fusarium avenaceum produce además un compuesto con mayor polaridad al de F. oxysporum en las mismas condiciones de cultivo (Figura 2). En el espectro de 1HNMR, de los extractos completos se advierten algunas diferencias en la zona de los grupos vinilo, ambas cepas presentan señales protones aromáticos y protones provenientes de grupos metoxilo y señales adyacentes a grupos carbonilo y amino. Con base en su coloración al ser revelados en CCD y patrones de desplazamientos en 1HNMR y lo reportado en la literatura, los pigmentos producidos por las cepas en este estudio podrían corresponder a pigmentos de tipo xanthona y antraquinona (Tatum et al., 1985; Kundu et al., 2016; Lebeau et al., 2017).
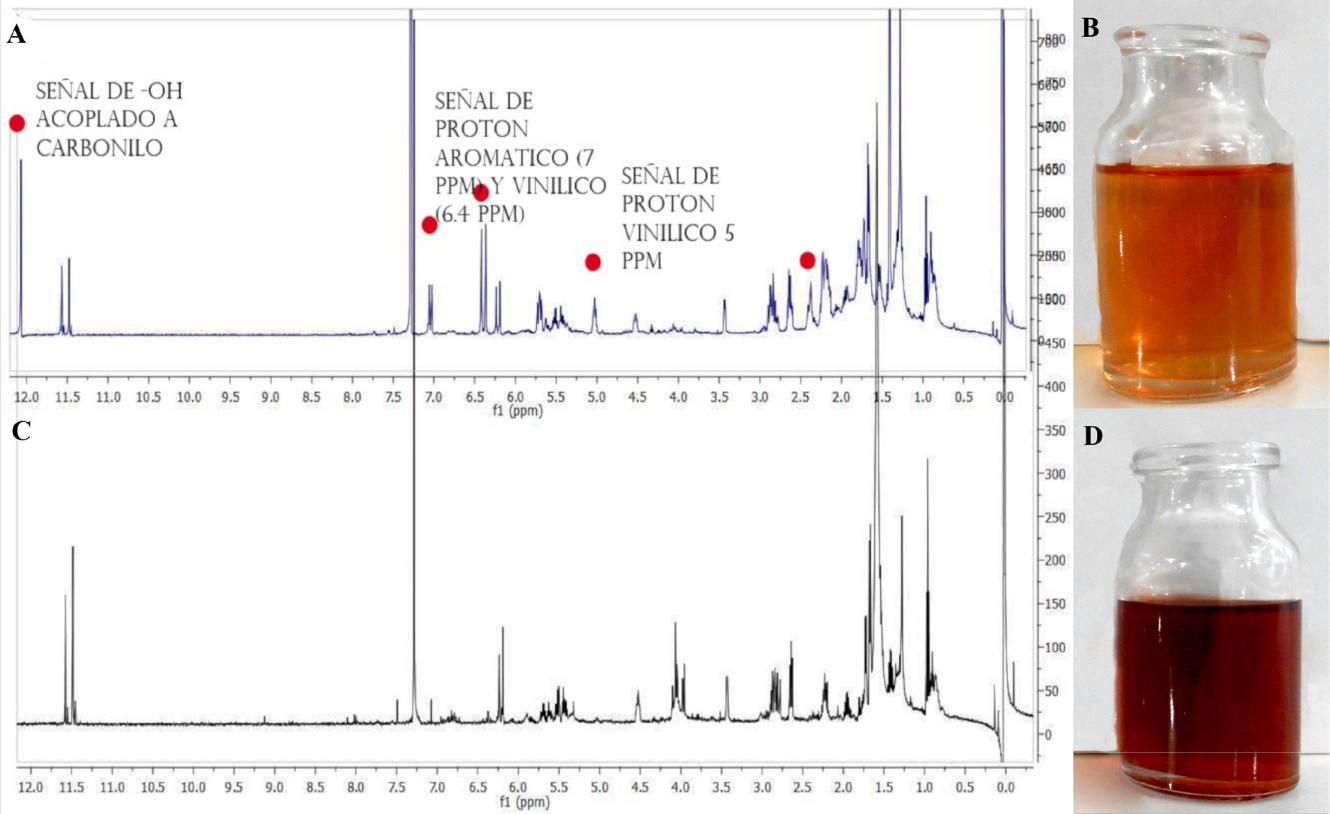
Figura 2. Análisis de los pigmentos obtenidos a partir de cepas de Fusarium avenaceum y F. oxysporum. A: espectro de Resonancia Magnética Nuclear de Protón (1HNMR) a 500 MHz de F. avenaceum. Se observan señales de protones aromáticos en 7 ppm y vinílicos en 6.5, además de señales de protones de hidroxilos cercanos a grupo carbonilo en 12 ppm. B: extracción con acetato de etilo de F. avenaceum. C: espectro de Resonancia Magnética Nuclear de Protón (1HNMR) a 500 MHz de F. oxysporum. D: extracción con acetato de etilo de F. oxysporum.
Discusión
Fusarium oxysporum es una especie cosmopolita y común en el suelo, contiene formas patogénicas, conocidas como formae speciales que pueden ocasionar enfermedades en más de 100 especies de plantas (Gordon y Martyn, 1997; Bosland, 1988). También existen cepas que no dañan al hospedero (no patógenicas) y que pueden actuar como posibles agentes de control biológico (Benhamou y Garand, 2001; Shishido et al., 2005). Algunos compuestos de tipo secundario de especies de Fusarium, contribuyen a vencer los mecanismos de defensa de las plantas hospederas y pueden matar a los insectos a través de la actividad de toxinas producidas por las hifas penetrantes (Gupta et al., 1991). Algunos de los metabolitos secundarios, reportados en cultivos in vitro de F. oxysporum formae speciales, son ácido fusárico, beauvaericina y pigmentos de color naranja a rojo como bikaverina y otros (Bell et al., 2003). La bikaverina, puede actúar como biocida contra diferentes organismos, como Leishmania brasiliensis, Phytophtora infestans y Bursaphelenchus xylophilus (nematodo del pino) (Balan et al., 1970; Kwon et al., 2007; Son et al., 2008).
En relación a Fusarium avenaceum, este habita cereales y maíz en zonas de clima templado y presenta capacidad patógenica en cultivos como manzana y papa (Booth, 1971). Puede producir in vitro micotoxinas como moniliformina, enniatina y fusarina por mencionar algunas (Sørensen y Giese, 2013). Existen estudios en donde se demuestra que F. avenaceum es entomo-patogénica en diversos insectos como gusano de la yema del abeto (Choristoneura fumiferana), diversas especies de mosquitas blancas (Trialeurodes sp., Bemisia sp.) y gorgojo del arroz (Sitophilus oryzae) (Strunz y Strongman, 1988; Rojas et al., 2003; Sun, 2008; Batta, 2012); además de mediano nivel de patogenicidad en algunos insectos del orden Lepidoptera (Teetor-Barsch y Roberts, 1983).
La mayoría de las especies de Fusarium producen pigmentos que van del color amarillo, naranja, rojo, violeta, azul y negro, que se han utilizado como carácter en las claves taxonómicas que corresponden a una variedad de moléculas de tipo xantona y antraquinona (Ashley et al., 1937; Goertz et al., 2010). Algunos de estos policétidos fúngicos son tóxicos para animales y humanos, pueden ser fitotóxicos, contribuyendo a la capacidad de los hongos para invadir y ocasionar daño a las plantas, otros presentan propiedades farmacéuticas benéficas como antibióticos, antiparasitarios, e insecticidas (Keller et al., 2005). Aunque no se pueden describir con exactitud las funciones de los metabolitos secundarios, generalmente se reconoce que los pigmentos protegen a los hongos de la exposición al estrés ambiental como la luz UV y la oxidación. (Duran et al., 2002; Yu. et al., 2004; Medentsev et al., 2005).
Conclusiones
Los extractos obtenidos de cultivos estacionarios de ambas especies, son una mezcla de metabolitos secundarios de tipo policetídico, los cuales se evidenciaron mediante señales características en 1HNMR y en CCD, comparando con lo reportado en la bibliografía, que pueden corresponder a pigmentos de tipo xantona y atraquinona, y micotoxinas, compuestos que producen una coloración anaranjada-rojiza en el medio. Aunque no fue el objetivo del presente trabajo, la presencia de estos compuestos sugiere algún tipo de patogenicidad en insectos como la mosquita blanca (Bemisia sp.), no así en la planta huésped; asimismo estas dos especies de Fusarium pueden ser entomo-patógenos débiles o pueden vivir como saprobios en insectos muertos.
Por lo que resultará interesante determinar la estructura molecular de los tres compuestos mayoritarios, observados en cromatografía en capa fina, pues las cepas identificadas que se obtuvieron a partir de cadáveres de Bemisia aff. tabaci, podrían presentar actividad biológica insecticida.

