










Services on Demand
Journal
Article
 text in
text in  English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkAbanico veterinario
On-line version ISSN 2448-6132Print version ISSN 2007-428X
Abanico vet vol.11 Tepic Jan./Dec. 2021 Epub Oct 11, 2021
https://doi.org/10.21929/abavet2021.13
Original Article
Genotype-by-environment interaction in white shrimp associated with White Spot Disease
*
1
 http://orcid.org/0000-0002-9439-7007
http://orcid.org/0000-0002-9439-7007
2
http://orcid.org/0000-0001-8676-7795
1Departamento de Genética y Bioestadística, Facultad de Medicina Veterinaria y Zootecnia, Universidad Nacional Autónoma de México. México.
2Departamento El Hombre y su Ambiente, Universidad Autónoma Metropolitana, Xochimilco, México.
3Departamento de Producción Agrícola y Animal, Universidad Autónoma Metropolitana, Xochimilco, México. §Fallecido, marzo 18, 2019.
This study aimed to estimate the genotype-by-environment interaction for body weight (BW) and harvest survival (HS), in the presence and absence of White Spot Disease (WSD) in two genetic lines of Penaeus vannamei (Growth -GRO- and resistance to WSD-RES-). The heritability for BW in the GRO line was 0.05 ± 0.16 in the presence of WSD and 0.35 ± 0.15 in the absence, while for the RES line it was 0.26 ± 0.07 and 0.49 ± 0.08 in the presence and absence of WSD, respectively. The genetic correlations for BW between environments were -0.17 ± 0.60 for GRO and 0.89 ± 0.09 for RES. The heritability for HS in GRO was 0.01 in both environments and the genetic correlation was not estimable, while, for RES, the heritabilities were 0.06 ± 0.04 and 0.11 ± 0.06 in the absence and presence of WSD, respectively, additionally, the genetic correlation it was not significant. Although the linear model suggests a genotype- by-environment interaction, the estimates propose independence of the same characteristic between environments, and the correlations between characteristics for the resistance line propose to independently select the characteristics when WSD is present.
Keywords: Penaeus vannamei; heritability; additive genetic correlation; body weight; survival
Este estudio tuvo como objetivo estimar la interacción genotipo por ambiente para peso corporal (PC) y supervivencia a cosecha (SC), en presencia y ausencia de Síndrome de Mancha Blanca (SMB) en dos líneas genéticas de Penaeus vannamei (Crecimiento -CRE- y resistencia a SMB -RES-). La heredabilidad para PC en la línea CRE fue 0.05 ± 0.16 en presencia de SMB y 0.35 ± 0.15 en ausencia, mientras que para la línea RES fueron 0.26 ± 0.07 y 0.49 ± 0.08 en presencia y ausencia de SMB, respectivamente. Las correlaciones genéticas para PC entre ambientes fueron -0.17 ± 0.60 para CRE y 0.89 ± 0.09 para RES. La heredabilidad para SC en CRE fue 0.01 en ambos ambientes y la correlación genética no fue estimable, mientras que, para RES, las heredabilidades fueron 0.06 ± 0.04 y 0.11 ± 0.06 en ausencia de y presencia de SMB, respectivamente, adicionalmente, la correlación genética no fue significativa. Aunque el modelo lineal sugiere una interacción genotipo por ambiente, los estimados proponen independencia de la misma característica entre ambientes y, las correlaciones entre características para la línea de resistencia proponen seleccionar de manera independiente las características cuando SMB esté presente.
Palabras clave: Penaeus vannamei; heredabilidad; correlación genética aditiva; peso corporal; supervivencia
INTRODUCTION
The world production of Pacific white shrimp (Penaeus vannamei) has been based on the production of genetic lines that have been selected for growth and general survival (Campos-Montes et al., 2009; Caballero-Zamora et al., 2015; Yuan et al., 2018). At the same time, the production units have been affected by several diseases with high rates of morbidity and mortality (Trang et al., 2019); among them, the White Spot Disease (WSD) (Hernández-Llamas et al., 2016).
The control of WSD has been a difficult goal and it has been decided to add selection criteria related to the resistance of this disease to the selection objective of Genetic Breeding Programs (GBP) in penaeids (Ødegård et al., 2011; Huang et al., 2012; Klinger and Naylor, 2012; Caballero-Zamora et al., 2015. In this idea, it is important to have adequate estimators of heritability (h2) and genetic correlation (rG) for the formulation of selection strategies. These genetic parameters, estimated under natural sprouting conditions, can provide important information to be considered in GBPs. Some authors have estimated the heritability for body weight in the presence of WSD between 0.09, using outbreak information and 0.21 from a controlled challenge (Gitterle et al., 2005b; Caballero-Zamora et al., 2015).
Regarding heritability for survival in the WSD presence, it has been estimated between 0.01 and 0.21 in controlled challenge studies based on different statistical models and infection protocols (Gitterle et al., 2005b; Gitterle et al., 2006a; Gitterle et al., 2006b), and as 0.06 in natural outbreak conditions of WSD (Caballero-Zamora et al., 2015). Instead, the estimation of these parameters for the same trait in different environments can be interpreted as a genotype-environment interaction (GEI). GEI can modify the estimation of h2 and rG between selection criteria, causing inaccurate responses to selection and alterations in the ordering of breeding candidates (Sae-Lim et al, 2016).
In P. vannamei, previous studies have searched GEI for body weight at harvest (BW) between locations or planting densities under commercial conditions, without finding evidence (Ibarra and Famula, 2008; Campos-Montes et al., 2009). However, Caballero- Zamora et al. (2015) observed effects of GEI for body weight at 19 weeks of age among populations that grew in the presence or in WSD absence under commercial conditions. Regarding GEI for general harvest survival (HS), no studies have reported effects of GEI.
Concerning the rG for weight and survival in shrimp, some studies have estimated the rG between BW and HS in the absence of any disease between -0.49 and 0.56 (Campos- Montes et al., 2013); while Caballero-Zamora et al. (2015) report that it was not possible to estimate this correlation in the presence of WSD, due to the loss of information structure derived from the high mortality in the population. On the other hand, there is no information on how these rG are modified for weight and survival across different environments in shrimp production. Therefore, it is important to estimate these genetic parameters (h2 and rG) in the presence or absence of WSD for the optimal design of GBPs.
Therefore, the objective of this study was to estimate the effects of GEI for BW and HS in two commercial environments (presence or absence of a natural outbreak of WSD), in two genetic lines of Pacific white shrimp (Penaeus vannamei), one selected for growth and another with a history of WSD resistance.
MATERIAL AND METHODS
The data were obtained from a Production Company of shrimp larvae located in northwestern Mexico. The records included shrimp from the 2016 production cycle. The shrimp were raised under commercial conditions in three ponds: two ponds were located on two farms established in Sonora state (Kino and Marea Alta); which presented a natural outbreak of WSD (WSD-presence), where the diagnosis was made from the symptoms and macroscopic changes typical of WSD (Stentiford and Lightner, 2011), during the production cycle and confirmed by PCR analysis. The third pond was located in Los Pozos community, Sinaloa (Pozos), where strict biosafety procedures were carried out and no symptoms of WSD were detected, nor was there a positive diagnosis by PCR (WSD- absence).
A line selected since 1998 for growth and HS (GRO), and another line with a history of resistance to WSD (RES) was used. The families considered from each line in this study had a maximum of 25% genes from the other line (Gallaga-Maldonado et al, 2020), and were analyzed independently. For the GRO line, 7,679 records from matings between 49 fathers and 69 mothers (families) were analyzed. For the RES line, 9,519 records were used with the progeny of 63 fathers and 91 mothers (families). The female to male ratio was 1.4 in both lines. The pedigree information included animals born since 2002 for GRO and since 2014 for RES.
Origin and development of genetic lines
The GRO and RES lines were formed in 1998 and 2014 respectively. The GRO line was produced using shrimp from Mexico, Venezuela, Colombia, the United States, and Ecuador. The RES line is composed of shrimp with a history of resistance to WSD from Ecuador, Panama and the United States and since 2014. A more specific description of the creation of the genetic lines can be found in Gallaga-Maldonado et al. (2020) and Campos-Montes et al. (2020).
Family management
The families were produced by artificial insemination, using a ratio of one male for every two females to form half-sib families. The inseminated females spawned in individual tanks for the counting of nauplii per family (full siblings); those spawning with less tan 25,000 nauplii being discarded for the next stage. The feeding in the larval stages was based on commercial micro-pellets with 40% to 50% protein and 8% to 10% fat, Chaetoceros microalgae, Spirulina spp., and Artemia spp., and the constitution of the diet was adapted to each stadium. The full sib families were kept in the same pond until they were marked around 60 days of age (weighing between 2 and 3 grams), using colored elastomers (Northwest Marine TechnologyTM), in the last abdominal segment of the shrimp and whose color combination worked as family identification.
Management of growing ponds
Ten days after tagging, an average of 36 shrimp per family were stocked in each pond. The ponds of Sonora (WSD-presence) were of sand of 0.20 ha, with a water column of 1.4 m, at an average temperature of 32 °C and an average salinity of 33 gL-1. The daily water exchange rate ranged from 5 to 20%. The food offered had a protein percentage between 34 and 40% at a rate of 3% of the total biomass in the pond. The stocking density in both ponds was 16 organisms/m2. In Sinaloa (WSD-absence) a 4 x 16 m concrete pond was used, with a water column of 2 m and a planting density of 70 m2. The water temperature was maintained at 30 °C with a salinity of 35 gL-1, constant aeration and a daily water exchange rate of 4 to 5%. The amount of daily food offered (35 - 40% protein); it was calculated as 6% of its biomass.
Data collection for body weight at 130 days of age and survival from 70 to 130 days
After 70 days after sowing, all the organisms were recovered from the ponds, and from each one the family of origin, sex and body weight were identified. The information of individuals with deformities, without reliable identification or undefined sex was eliminated. For the estimation of the HS, the animals recovered at the end of the period were considered as alive (1) and the animals not recovered, considering the difference between the living organisms of each family and those sown, as dead (0).
Information analysis
To compare the productive behavior between both lines in both scenarios (WSD-presence and WSD-absence), the following linear model was considered:
yijk = µ + Li + Sj + LSij + eijk
Where, yijk is the vector of observations of BW or HS, µ is the mean of the population for the variable of interest (HS or BW), Li is the effect of the i-th line (GRO, RES), Sj is the effect of the j-th environment (WSD-presence, WSD-absence), LSij is the effect of the interaction between the line and pond health status, and eijk~N (0, σ2e). Sex and the pond were also included on BW. To determine differences between line and environment combinations, a Tukey test (α = 0.05) was used.
The genetic parameters for BW and HS were estimated for each line, using an animal model and restricted maximum likelihood, with the ASReml software. Considering the criteria for approximation of a binomial distribution to a normal distribution (Schader and Schmid, 1989; Emura and Yu-Ting, 2018), normality was assumed in the HS analysis; the model used was:
y = Xβ + Zu +Wf + ε
Where, y is the vector of observations of (BW or HS) of both environments, β is the vector of fixed effects of each characteristic, u is the vector of random additive genetic effects of the animal, u~MVN (0, G), where G = V ⊗ A, where V is a symmetric matrix containing the (co) variances between the effects of same family animals for the characteristics in the two environments, and A is a matrix of additive relationships; ⊗ is the Kronecker product, environmental residuals, f is the unknown vector of the common family effect for all characteristics, f~MVN (0, F); where, F=C ⊗ I, where, C is a matrix of co (variances) of common family environment effects, only for BW, and I is an identity matrix of appropriate order, and ε is the vector of effects random, ε~MVN (0, R), where, R = E ⊗ I, where, E is the matrix of co (variances) of the residual effects that contains the covariances between the two characteristics, and I is an identity matrix of appropriate order, with σ2e as the residual variance.
Finally, X, Z, and W are known incidence matrices that relate the observations to the fixed effects (which varied depending on the trait analyzed), the genetic effects of the animal, and common family environment effects, respectively. The genetic correlations between both characteristics in the line-pond combination were estimated with ASReml, using bivariate models and with the same model, but considering the y vector of information from BW and HS. No restrictions were used in the covariance structure and common family environment effects were considered independent.
In estimating the genetic parameters for BW, the fixed effects included in the model were: sex, age at harvest linear and quadratic, additionally in the case of WSD-presence; the pond effect was included (Kino and Marea Alta). Regarding the HS for the affected ponds, the only fixed effect considered was that of the pond in WSD-presence; while in the WSD- absence environment, no fixed effect was considered.
The phenotypic variance for each characteristic was estimated as the sum of the variance components of the random effects (animal genetic and common family). The h2 was estimated as phenotypic variance proportion that is due to the additive genetic variance, and the rG was estimated as the covariance divided by the product of the corresponding standard deviations. The statistical significance of the estimated parameters was based on the confidence intervals (95%), constructed with their standard errors, assuming normality. The existence of GEI was determined when rG between environments was less than 0.80 (Sae-Lim et al., 2016).
Finally, to analyze whether the behavior of the characteristics between genetic lines was similar. A comparison of the estimated rG was made in each line (Nguyen et al., 2016), such comparison was made by means of a Fisher's Z transformation (Rosenthal et al., 1992) implemented in the “Cocor” package in R (Diedenhofen and Musch, 2014), a significance test for the difference between 2 correlations, based on dependent groups with 1 variable in common. For a better understanding, a diagram of the estimated genetic correlations per line is presented in Figure 1.
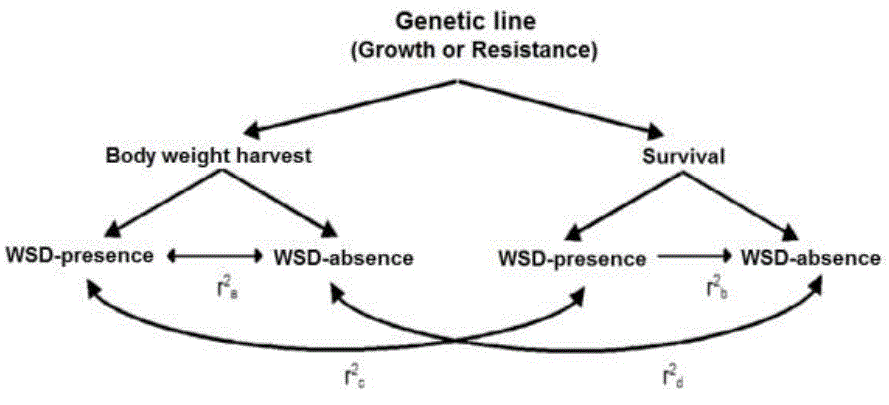
Figure 1 Scheme of the genetic correlations estimated in the study by genetic line r2a and r2b = Genetic correlation between environments (WSD-presence and WSD-absence) within each characteristic. r2c = Genetic correlation between WSD environments-presence of the two characteristics. r2d = Genetic correlation between WSD environments-absence of the two characteristics
RESULTS AND DISCUSSION
Comparison of productive behavior between lines
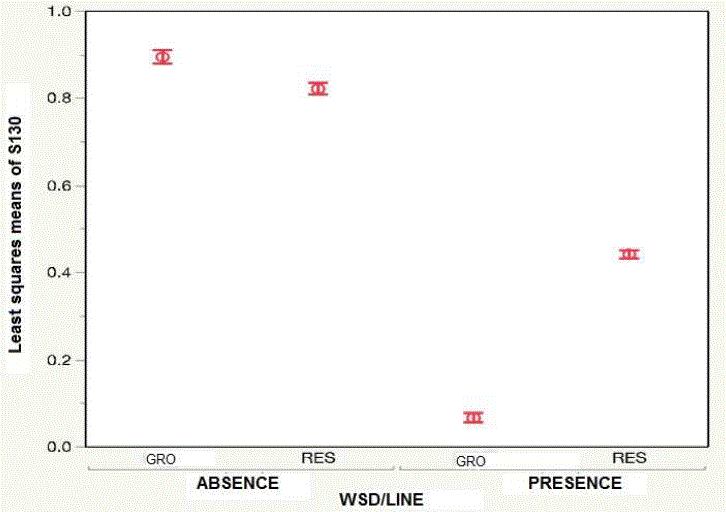
Descriptive statistics for BW and HS in each genetic line (GRO and RES), within environment (WSD-presence, WSD-absence) are shown in Table 1. At the same time, Figures 2 and 3 show the LSM of both lines (GRO and RES), through the environments for BW and HS. These results show differences in HS, where the GRO line has a low HS in WSD-presence; while the shrimp of the RES line have a lower HS in WSD-presence. Additionally, there is line-by-environment interaction (P <0.0001) in the two characteristics. These line-by-environment interactions highlight the importance of considering the probability of occurrence of WSD disease, when the line is chosen in the genetic improvement program (Sae-Lim et al., 2016).
Table 1 Number of individuals (n) and least squares means for body weight and survival rate to harvest in the growth line and in the Resistance line, in the presence and absence of White Spot Disease
| Environment | Line | Body Weigth(g) | Survival Rate | ||
|---|---|---|---|---|---|
| n | LSM±se | n | LSM±se | ||
| WSD-presence | RES | 2,524 | 12.80±0.07a | 6,414 | 0.45±0.01a |
| GRO | 294 | 8.75±0.06b | 5,494 | 0.06±0.01b | |
| WSD-absence | RES | 2,838 | 11.91±0.17c | 3,105 | 0.82±0.01c |
| GRO | 1,926 | 13.75±0.05d | 2,185 | 0.88±0.01d | |
LSM: least squares means, se: standard error.
*The different literals within the columns indicate statistically significant differences TUKEY (α = 0.05).
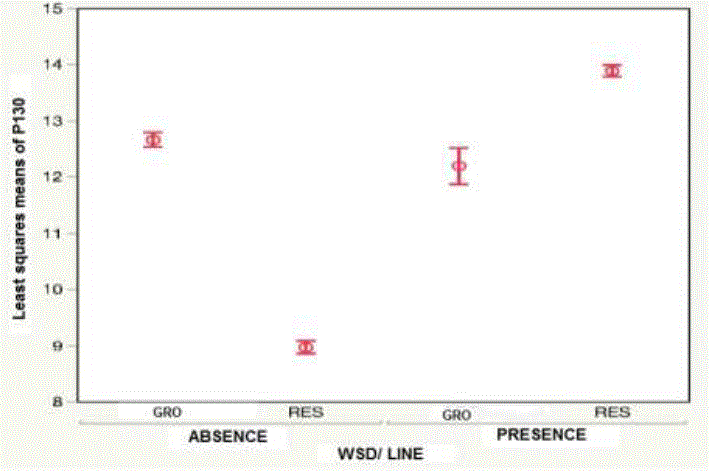
Figure 2 Least squares means for body weight at harvest in the growth lines and resistance to WSD, in the presence and absence of SMB in P. vannamei
Heritability for body weight at harvest
The heritabilities for BW in both lines are shown in Table 2. The common family environment effects were 0.05 in both lines. The inclusion of these effects in all the models reduced the estimation of the additive variance, according to what was presented by other authors (Campos-Montes et al., 2013; Montaldo et al., 2013). The heritabilities in WSD- absence were consistent with those reported by authors such as Tan et al. (2017), Trang et al. (2019) and Ren et al. (2020), despite the fact that the latter did not consider common family environment effects in its model; however, these heritability estimates were higher than those reported by other authors (Li et al., 2015; Zhang et al., 2017; Yuan et al., 2018). On the other hand, the heritabilities for WSD-presence are similar to those estimated by Caballero-Zamora et al. (2015), who also used data from a natural outbreak of WSD.
Table 2 Heritability and additive genetic correlations for body weight in growth and resistance lines by environment *
| Enviroment | WSD-presence | WSD-absence | |
|---|---|---|---|
| WSD-presence | 0.05±0.16 | -0-17±0.60 | |
| Growth line | WSD-absence | 0.35±0.15 | |
| WSD-presence | 0.26±0.07 | 0.89±0.09 | |
| Resistances Line | WDS-absence | 0.49±0.08 |
*Estimated by bivariate analysis. Genetic correlations are shown in bold
In the GRO line, the difference between the heritabilities for WSD-presence (0.05 ± 0.16) and WSD-absence (0.35 ± 0.15), can be an indicator of heterogeneity of variances (Sae- Lim et al, 2016). In this case, in addition to changes in the additive genetic variance, the source of this heteroscedasticity may be contained in changes in the environmental variance due to the microenvironmental sensitivity of the individuals. In the case of WSD- presence, it is important to mention that the low precision in the estimate of h2 may be due to the data structure resulting from the high mortality. In the RES line, the heritability estimates for BW were higher than for GRO; in addition to having better precision. On the other hand, the estimate of h2 in WSD-presence (0.26 ± 0.07) is lower than in WSD- absence (0.49 ± 0.08), as in the GRO line, and the estimators of this line were consistent with other authors (Luan et al., 2015; Sui et al., 2016; Zhang et al., 2017 and Yuan et al., 2018).
Variations in heritability estimators for BW may be the result of the microenvironmental sensitivity of individuals (Sae-Lim et al, 2016). Given the above, it is important to note that these changes in heritability can alter the response prediction accuracy to selection.
Genetic correlations for BW
The estimated rG for the BWs between environments in the GRO line was negative and not significant (P> 0.05) (-0.17 ± 0.60); although in estimates with preliminary models (data not shown) it was consistently negative. The imprecise value of the estimate may be a low value consequence of the additive genetic variance of WB in WSD-presence. It complicates the evaluation of GEI effects in this line; while in the RES line, this correlation was not different from 1 (P> 0.05) (Table 2); noting that the additive genetic effects for BW are very similar in the two environments (Sae-Lim et al., 2016). In other words, there is no GEI effect on RES for BW. Considering that both lines were under the same environmental management conditions and exposed to the same pathogen (WSD), it is possible to consider that the differences in the estimators of both lines are the result of the low HS rate of the GRO line. In several studies GEI was not detected when considering environmental conditions, such as planting density (Campos-Montes et al., 2009; Tan et al., 2017) or cultivation location (Sui et al., 2016). Other works report possible effects of GEI for BW, considering the point values of the rG estimators. However, their standard errors do not allow defining them as significantly different from one or zero (Caballero- Zamora et al., 2015; Li et al., 2015; Nguyen et al., 2020).
Heritability for survival to harvest in both genetic lines
The heritability and additive genetic correlations for HS in the genetic lines are shown in Table 3. Regarding the common family effects, these were considered independent between environments for each characteristic, and varied between 0.02 and 0.04. The estimated heritabilities for HS in WSD-presence were 0.01 and 0.11 for GRO and RES respectively; the previous ones are similar to those reported by other authors in the presence of WSD (Gitterle et al., 2005a; Caballero-Zamora et al., 2015); however, they were lower than those estimated by Li et al. (2015) and Trang et al. (2019); both in WSD controlled challenges. The results of the HS model were consistent with estimates, using univariate models, considering a binomial distribution (results not shown).
Table 3 Heritability and additive genetic correlations for survival in growth lines and resistance by environment *
| Enviroment | WSD-presence | WSD-absence | |
|---|---|---|---|
| WSD-precense | 0.01±0.02 | 0.00 | |
| Growth line | WSD-absence | 0.01±0.03 | |
| WSD-presence | 0.11±0.06 | 0.10±0.40 | |
| Resistance line | WSD-absence | 0.06±0.04 |
*Estimated using bivariate models Genetic correlations are shown in bold
The heritabilities for HS in WSD-absence were 0.01 ± 0.03 in GRO and 0.06 ± 0.04 in RES concordant with other estimators for overall survival (Campos-Montes et al., 2013; Zhang et al., 2017; Ren et al., 2020), but lower than those presented by Lu et al. (2017), Luan et al. (2020), and Tan et al. (2017), where the effect of a common family environment was not included, which could have generated an overestimation of the genetic parameters.
The heritabilities in GRO were essentially zero in both environments, 0.01 ± 0.02 in SMB- presence and 0.02 ± 0.03 in WSD-absence, which represents minimal possibilities of genetic advancement by selection for this characteristic in both scenarios, which is consistent with other authors (Gitterle et al., 2005a; Campos-Montes et al., 2013; Li et al., 2015; Lu et al., 2017). The minimum advance by selection could be related to the difficulty in the estimation due to the mortality rate by the statistical models (Vehviläinen et al., 2008), a very low genetic proportion in the survival expression, or possibly, the damage in the structure of genetic family relationships when WSD was present.
The estimated heritabilities for HS in RES were 0.11 ± 0.06 and 0.06 ± 0.04, for WSD- presence and WSD-absence, respectively (Table 3); which were not statistically different from those estimated for the GRO line. The h2 for HS in WSD-presence was higher than that estimated by Caballero-Zamora et al. (2015) 0.06 and that reported by Gitterle et al. (2005b), who calculated values between 0.03 and 0.07. The h2 for WSD-absence was concordant with other estimators for general survival in the same species (Campos- Montes et al., 2013; Zhang et al., 2017; Ren et al., 2020).
The heritabilities of HS for both environments were consistent in both lines (Table 3), suggesting that there is no compression of the additive variance, in the lines, associated with the environment (Sae-Lim et al., 2016). The genetic correlations between HS in WSD- presence and WSD-absence were not different from zero (P> 0.05), coinciding with that reported by Vehviläinen et al. (2010) in another aquatic species, which may suggest that survivals in both environments are independent characteristics, as suggested by Thoa et al. (2015).
Genetic correlations between body weight and harvest survival
The genetic correlations by environment between BW and HS are shown in Table 4. For the estimation of these correlations, the common family effect was considered independent between environments.
Table 4 Genetic correlations within lines (Growth and Resistance) between body weight to harvest (BW) and harvest survival for environments in the presence or absence of WSD
| WSD-presence | WSD-absenc | |
|---|---|---|
| Growth line | Ne* | -0.09±0.92 |
| Resistance line | 0.04±0.16 | 0.57±0.20 |
*Not estimable
The genetic correlation between the two characteristics could not be estimated in WSD- presence in the GRO line, possibly due to the affectation in the structure of the information associated with the high mortality presented in that line; whereas, for the RES line, it was not different from zero (0.04 ± 0.16), agreeing with the results of Caballero-Zamora et al. (2015). In the case of WSD-absence for the GRO line, the rG was not different from zero (-0.09 ± 0.92), unlike the one estimated in the RES line which was 0.57 ± 0.20, which was consistent with that estimated by Campos-Montes et al. (2013), in commercial ponds, in the absence of SMB (0.56 ± 0.10). The rG estimated in the RES line was higher than that reported by Yuan et al. (2018), Gitterle et al. (2005a) , and Zhang et al. (2017). The differences between the genetic correlations in RES may be an indicator of changes in the variance components possibly associated with GEI, in turn related to the corresponding covariance, which would have implications in the response to correlated selection (Sae-Lim et al., 2016).
Some authors highlight the importance of the creation of genetic lines in aquaculture (Nguyen et al., 2016), the results of this study suggest that the selection indices for BW should take into account the genetic line used in the genetic improvement program. On the other hand, the estimation of genetic parameters related to BW must consider the presence of endemic diseases, such as WSD in shrimp culture and visualize HS in the presence and absence of WSD as independent characteristics, in both genetic lines.
In addition to changes in heritabilities and genetic correlations in both lines, their productivity was different in the environments studied, which could be interpreted as an indicator of phenotypic plasticity, which may be common in marine organisms as suggested by Munday, (2013). It can be understood as the expression of different phenotypes in individuals with the same genotype, but under different environmental conditions (Munasinghe and Seneviratha, 2015). Consequently, this phenotypic plasticity effect should be considered in shrimp genetic improvement programs, ensuring that the performance environments to be analyzed are as close to the production conditions as possible (Nguyen et al., 2016).
CONCLUSIONS
The results of the linear model suggest differences between the lines, both for body weight and for survival across environments; however, the estimates of the genetic correlations do not allow considering GEI effects within the line in both characteristics, which would indicate that they are independent. Furthermore, the genetic correlations between the characteristics of the resistance line propose to treat them as independent variables, when WSD is present in the environment.
LITERATURA CITADA
Caballero-Zamora A, Montaldo HH, Campos-Montes GR, Cienfuegos-Rivas EG, Martínez-Ortega A, Castillo-Juárez H. 2015. Genetic parameters for body weight and survival in the Pacific White Shrimp Penaeus (Litopenaeus) vannamei affected by a White Spot Syndrome Virus (WSSV) natural outbreak. Aquaculture. 447:102-107. ISSN: 0044- 8486. https://doi.org/10.1016/j.aquaculture.2014.08.028 [ Links ]
Campos-Montes GR, Montaldo HH, Martínez-Ortega A, Castillo-Juárez H. 2009. Genotype by environment interaction effects for body weight at 130 days of age in the Pacific white shrimp [Penaeus (Litopenaeus) vannamei]. Veterinaria México. 40(3):255-267. ISSN: 2448-6760. https://www.medigraphic.com/cgi-bin/new/resumenI.cgi?IDARTICULO=22647 [ Links ]
Campos-Montes GR, Montaldo HH, Martínez-Ortega A, Jiménez AM, Castillo-Juárez H. 2013. Genetic parameters for growth and survival traits in Pacific white shrimp Penaeus (Litopenaeus) vannamei from a nucleus population undergoing a two-stage selection program. Aquaculture International. 21:299-310. ISSN: 1573-143X. https://doi.org/10.1007/s10499-012-9553-1 [ Links ]
Campos-Montes GR, Caballero-Zamora A, Montaldo HH, Montoya-Rodríguez L, Rodríguez-Sala BGG, Rodríguez SAS, Martínez-Ortega A, Quintana-Casares JC, Castillo-Juárez H. 2020. Genetic (co) variation in resistance of Pacific white shrimp Litopenaeus vannamei to acute hepatopancreatic necrosis disease (AHPND) and white spot syndrome virus (WSSV) in challenge tests. Aquaculture. 520:734994. ISSN: 0044-8486. https://doi.org/10.1016/j.aquaculture.2020.734994 [ Links ]
Diedenhofen B, Musch J. 2015. cocor: A comprehensive solution for the statistical comparison of correlations. PloS one. 10(4):e0121945. ISSN: 1932-6203. https://doi.org/10.1371/journal.pone.0121945 [ Links ]
Emura T, Liao YT. 2018. Critical review and comparison of continuity correction methods: The normal approximation to the binomial distribution. Communications in Statistics-Simulation and Computation. 47(8):2266-2285. ISSN: 0361-0918. https://doi.org/10.1080/03610918.2017.1341527 [ Links ]
Gallaga-Maldonado EP, Montaldo HH, Castillo-Juárez H, Campos-Montes GR, Martínez-Ortega A, Quintana-Casares JC, Montoya-Rodríguez L, Betancourt-Lozano M, Lozano-Olvera R, Vázquez-Peláez C. 2020. Crossbreeding effects for White Spot Disease resistance in challenge tests and field pond performance in Pacific white shrimp Litopenaeus vannamei involving susceptible and resistance lines. Aquaculture. 516:734-527. ISSN: 0044-8486. https://doi.org/10.1016/j.aquaculture.2019.734527 [ Links ]
Gitterle T, Rye M, Saltec R, Cock J, Johansen H, Lozano C, Suárez J, Gjerde B. 2005a. Genetic (co)variation in harvest body weight and survival in (Penaeus (Litopenaeus) vannamei) under standard commercial conditions. Aquaculture. 243:83-92. ISSN: 0044-8486. https://doi.org/10.1016/j.aquaculture.2004.10.015 [ Links ]
Gitterle T, Saltec R, Gjerde B, Cock J, Johansen H, Salazar M, Lozano C, Rye M. 2005b. Genetic (co)variation in resistance to White Spot Syndrome Virus (WSSV) and harvest weight in Penaeus (Litopenaeus) vannamei. Aquaculture. 246:139-149. ISSN: 0044-8486. https://doi.org/10.1016/j.aquaculture.2005.02.011 [ Links ]
Gitterle T, Odegard J, Gjerde B, Rye M, Salte R. 2006a. Genetic parameters and accuracy of selection for resistance to white spot syndrome virus (WSSV) in Penaeus (Litopenaeus) vannamei using different statistical models. Aquaculture. 251:210-218. ISSN: 0044-8486. https://doi.org/10.1016/j.aquaculture.2005.06.008 [ Links ]
Gitterle T, Gjerde B, Cock J, Salazar M, Rye M, Vidal O, Lozano C, Erazo C, Salte R. 2006b. Optimization of experimental infection protocols for the estimation of genetic parameters of resistance to White Spot Syndrome Virus (WSSV) in Penaeus (Litopenaeus) vannamei. Aquaculture. 261:501-509. ISSN: 0044-8486. https://doi.org/10.1016/j.aquaculture.2006.07.017 [ Links ]
Hernández-LLamas A, Cabanillas-Ramos J, Magallon-Barajas FJ. 2016. Estimating impact of white spot disease on economic risk in semi-intensive shrimp farms in Mexico: the case of the State of Sinaloa. Reviews in Aquaculture. 8(2):111-120. ISSN: 1753-5123. https://doi.org/10.1111/raq.12084 [ Links ]
Huang Y, Yin Z, Weng S, He J, Li S. 2012. Selective breeding and preliminary comercial performance of Penaeus vannamei for resistance to resistance to White spot síndrome virus (WSSV). Aquaculture. 364-365:111-117. ISSN: 0044-8486. https://doi.org/10.1016/j.aquaculture.2012.08.002 [ Links ]
Ibarra AM, Famula TR. 2008. Genotype by environment interaction for adult body weights of shrimp Penaeus vannamei when grown at low and high densitie. Genetics Selection Evolution. 40(5):541. ISSN: 0999-193X. https://doi.org/10.1186/1297-9686-40-5-541 [ Links ]
Klinger D, Naylor R. 2012. Searching for solutions in aquaculture: charting a sustainable course. Annual Review of Environment and Resources. 37:247-276. ISSN: 1543-5938. https://doi.org/10.1146/annurev-environ-021111-161531 [ Links ]
Li W, Luan S, Luo K, Sui J, Xu X, Tan J, Kong J. 2015. Genetic parameters and genotype by environment interaction for cold tolerance, body weight, and survival of the Pacific white shrimp Penaeus vannamei at different temperatures. Aquaculture. 441:8-15. ISSN: 0044-8486. https://doi.org/10.1016/j.aquaculture.2015.02.013 [ Links ]
Lu X, Luan S, Cao B, Sui J, Dai P, Meng X, Luo K, Kong J. 2017. Heterosis and heritability estimates for the survival of the Pacific white shrimp (Litopenaeus vannamei) under the commercial scale ponds. Acta Oceanologica Sinica. 36(2):62-68. ISSN: 0253-505X. https://doi.org/10.1007/s13131-016-0942-6 [ Links ]
Luan S, Luo K, Chai Z, Cao B, Meng X, Lu X, Liu N, Xu S, Kong J. 2015. An analysis of indirect genetic effects on adult body weight of the Pacific white shrimp Litopenaeus vannamei at low rearing density. Genetics Selection Evolution. 47:95. ISSN: 0999-193X. https://doi.org/10.1186/s12711-015-0164-y [ Links ]
Luan S, Qiang G, Cao B, Luo K, Meng X, Chen B, Kong J. 2020. Feed competition reduces heritable variation for body weight in Litopenaeus vannamei. Genetics Selection Evolution. 52:45. ISSN: 0999-193X. https://doi.org/10.1186/s12711-020-00565-3 [ Links ]
Montaldo HH, Castillo‐Juárez H, Campos‐Montes G, Pérez‐Enciso M. 2013. Effect of the data family structure, tank replication and the statistical model, on the estimation of genetic parameters for body weight at 28 days of age in the Pacific white shrimp (Penaeus (Litopenaeus) vannamei Boone, 1931). Aquaculture Research. 44(11):1715-1723. ISSN: 1355-557X. https://doi.org/10.1111/j.1365-2109.2012.03176.x [ Links ]
Munasinghe DHN, Senevirathna JDM. 2015. Phenotypic plasticity and genetic variation of two wild populations of green tiger shrimp (Penaeus semisulcatus - De Haan, 1844). International Journal of Marine Science. 5:1-8. ISSN: 1927-6648. http://www.aquapublisher.com/index.php/ijms/article/view/1561 [ Links ]
Munday PL, Warner RR, Monro K, Pandolfi JM, Marshall DJ. 2013. Predicting evolutionary responses to climate change in the sea. Ecology letters. 16(12):1488-1500. ISSN: 1461-023X. https://doi.org/10.1111/ele.12185 [ Links ]
Nguyen NH. 2016. Genetic improvement for important farmed aquaculture species with a reference to carp, tilapia and prawns in Asia: achievements, lessons and challenges. Fish and Fisheries. 17(2):483-506. ISSN: 1467-2979. https://doi.org/10.1111/faf.12122 [ Links ]
Nguyen NH, Ninh NH, Hung NH. 2020. Evaluation of two genetic lines of Pacific White leg shrimp Liptopenaeus vannamei selected in tank and pond environments. Aquaculture. 516:734522. ISSN: 0044-8486. https://doi.org/10.1016/j.aquaculture.2019.734522 [ Links ]
Ødegård J, Baranski M, Gjerde B, Gjedrem T. 2011. Methodology for genetic evaluation of disease resistance in aquaculture species: challenges and future prospects. Aquaculture research. 42:103-114. ISSN: 1355-557X. https://doi.org/10.1111/j.1365-2109.2010.02669.x [ Links ]
Ren S, Prentis P, Mather PB, Li Y, Tang B, Hurwood DA. 2020. Genetic parameters for growth and survival traits in a base population of Pacific white shrimp (Litopenaeus vannamei) developed from domesticated strains in China. Aquaculture. 523:735-148. ISSN: 0044-8486. https://doi.org/10.1016/j.aquaculture.2020.735148 [ Links ]
Rosenthal R, Rubin DB, Meng XL. 1992. Comparing correlated correlation coefficients. Psychological bulletin. 111(1):172-175. ISSN: 0033-2909. APA Psycnet - http://nrs.harvard.edu/urn-3:HUL.InstRepos:11718225 [ Links ]
Sae‐Lim P, Gjerde B, Nielsen HM, Mulder H, Kause A. 2016. A review of genotype by‐ environment interaction and micro‐environmental sensitivity in aquaculture species. Reviews in Aquaculture. 8(4):369-393. ISSN: 1753-5123. https://doi.org/10.1111/raq.12098 [ Links ]
Schader M, Schmid F. 1989. Two rules of thumb for the approximation of the binomial distribution by the normal distribution. The American Statistician. 43(1):23-24. ISSN: 0003-1305. https://doi.org/10.1080/00031305.1989.10475601 [ Links ]
Stentiford GD, Lightner DV. (2011). Cases of white spot disease (WSD) in European shrimp farms. Aquaculture. 319(1-2):302-306. ISSN: 0044-8486. https://doi.org/10.1016/j.aquaculture.2011.06.032 [ Links ]
Sui J, Luan S, Luo K, Meng X, Lu X, Cao B, Li W, Chai Z, Liu N, Xu S, Kong J. 2016. Genetic parameters and response to selection for harvest body weight of Pacific white shrimp, Litopenaeus vannamei. Aquaculture Research. 47(9):2795-2803. ISSN: 1355- 557X. https://doi.org/10.1111/are.12729 [ Links ]
Tan J, Luan S, Luo K, Guan J, Li W, Sui J, Guo Z, Xu S, Kong J. 2017. Heritability and genotype by environment interactions for growth and survival in Litopenaeus vannamei at low and high densities. Aquaculture Research. 48(4):1430-1438. ISSN: 1355-557X. https://doi.org/10.1111/are.12978 [ Links ]
Thoa NP, Knibb W, Ninh NH, Van Dai N, Nhat PH, Nguyen NH. 2015. Genetic variation in survival of tilapia (Oreochromis niloticus, Linnaeus, 1758) fry during the early phase of rearing in brackish water environment (5-10 ppt). Aquaculture. 442:112-118. ISSN: 0044- 8486. https://doi.org/10.1016/j.aquaculture.2015.02.040 [ Links ]
Trang TT, Hung NH, Ninh NH, Knibb W, Nguyen NH. 2019. Genetic Variation in Disease Resistance Against White Spot Syndrome Virus (WSSV) in Liptopenaeus vannamei. Frontiers in Genetics. 10:264. ISSN: 1664-8021. https://doi.org/10.3389/fgene.2019.00264 [ Links ]
Vehviläinen H, Kause A, Koskinen H, Paananen T. 2010. Genetic architecture of rainbow trout survival from egg to adult. Genetics research. 92(1):1-11. ISSN: 0016-6723. https://doi.org/10.1017/s0016672310000017 [ Links ]
Vehviläinen H, Kause A, Quinton C, Koskinen H, Paananen T. 2008. Survival of the currently fittest: genetics of rainbow trout survival across time and space. Genetics. 180(1):507-516. ISSN: 0016-6731. https://doi.org/10.1534/genetics.108.089896 [ Links ]
Yuan R, Hu Z, Liu J, Zhang J. 2018. Genetic parameters for growth-related traits and survival in Pacific white shrimp, Litopenaeus vannamei under conditions of high ammonia- N concentrations. Turkish Journal of Fisheries and Aquatic Sciences. 18(1):37-47. ISSN: 1303-2712. https://doi.org/10.4194/1303-2712-v18_1_05 [ Links ]
Zhang J, Cao F, Liu J, Yuan R. 2017. Genetic parameters for growth and survival traits in Litopenaeus vannamei at different ages. Aquaculture International. 25(5):1901-1911. ISSN: 0967-6120. https://doi.org/10.1007/s10499-017-0163-9 [ Links ]
Received: December 02, 2020; Accepted: March 20, 2021
 Este es un artículo publicado en acceso abierto bajo una licencia
Creative Commons
Este es un artículo publicado en acceso abierto bajo una licencia
Creative Commons
