











 text new page (beta)
text new page (beta) Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
Permalink
Introducción
Las tilapias pertenecen a la familia Cichlidae, entre sus representantes destacan la tilapia del Nilo (Oreochromis niloticus), la tilapia azul (O. aureus) y la tilapia de Mozambique (O. Mossambicus); de estas especies, la tilapia del Nilo es la especie más importante en la acuicultura, debido a su rápido crecimiento y tolerancia a un amplio rango de temperaturas y salinidades (Tran et al. 2011). Sin embargo, alcanzar la uniformidad de talla al momento de la cosecha es complicado, por su maduración precoz, ya que dirigen una gran cantidad de energía a la actividad reproductiva, lo que afecta la cantidad de energía disponible para el crecimiento (Cnaani y Hulata 2008, Tran et al. 2011). Para contrarrestar este efecto se utilizan diferentes técnicas de cultivo entre las cuales destacan los cultivos de un solo sexo (monosexo) que buscan evitar y/o reducir la maduración precoz; siendo la reversión sexual mediante esteroides sexuales una de las técnicas más utilizadas actualmente a nivel comercial (Alcántar-Vázquez et al. 2014a, Vinarukwong et al. 2018, Chávez-García 2020).
La producción de poblaciones monosexo (compuestas exclusivamente por machos) mediante la reversión sexual ha permitido incrementar la rentabilidad de las granjas acuícolas, por la rápida tasa de crecimiento de los machos en comparación con las hembras y sobre todo porque se elimina el comportamiento sexual y territorial en ambos sexos (Desprez et al. 2003, Ponzoni et al. 2005). La aplicación de esteroides sexuales para alcanzar la reversión sexual se realiza principalmente con las dietas comerciales proporcionadas a los alevines después de la eclosión, antes de la diferenciación sexual, lo cual permite obtener hasta un 100% de machos (Jiménez y Arredondo 2000, Daza et al. 2005). Sin embargo, el uso de grandes volúmenes de esteroides sexuales para obtener poblaciones monosexo ha generado una creciente preocupación por parte de grupos ambientalistas, ya que la acumulación de esteroides en los cuerpos de agua cercanos a las granjas, puede alterar las proporciones sexuales de animales silvestres que habitan dichas zonas (Murray et al. 2016). De igual forma, un creciente número de personas no desean consumir productos que han sido tratados con hormonas o substancias activas (Piferrer 2001, Müller y Hörstgen 2007, Leet et al. 2011). Lo anterior ha convertido a la masculinización en una técnica cada vez más controvertida, impulsando la búsqueda de técnicas como la producción de machos YY (Mair et al. 1997, Alcántar-Vázquez et al. 2014b). Sin embargo, la producción de machos YY no ha probado ser una solución a corto plazo como estaba planeada. Por lo que una solución de amplio alcance y de aplicación inmediata es la optimización de los protocolos utilizados para revertir el sexo, los cuales fueron diseñados sin conocer el periodo exacto donde ocurre la diferenciación sexual de la tilapia del Nilo (Guerrero 1975, Shelton et al. 1981) y probablemente sobreestiman la cantidad de tiempo necesario para completar la masculinización, especialmente en variedades altamente domesticadas, como la variedad Spring. Por lo que incorporar el conocimiento reportado en los últimos años sobre el proceso de diferenciación sexual de la tilapia del Nilo y especies relacionadas (Ijiri et al. 2008, Baroiller et al. 2009, Tao et al. 2013, Gennotte et al. 2014) al proceso de masculinización, podría resultar en una reducción del número de días necesarios para alcanzar un 100% de masculinización, lo cual permitirá reducir el uso de esteroides sexuales en el cultivo comercial, manteniendo los mismos rendimientos y asegurando un producto más amigable con el ambiente. Sin embargo, en los últimos años pocos estudios han abordado la optimización de los protocolos de masculinización, concentrándose principalmente en la dosis del esteroide (Rima et al. 2017, Singh et al. 2017, Sreenivasa y Prabhadevi 2018), la temperatura del agua (Khater et al. 2017, Teng et al. 2020), el color del acuario (Rebougas et al. 2014), el uso de complejos homeopáticos para mejorar la supervivencia (Dias-Neto et al. 2017) durante el proceso de masculinización, y el uso extractos de plantas como posibles agentes androgénicos (Yusuf et al. 2019). Solo Vinarukwong et al. (2018) reportan la evaluación del periodo de masculinización con 17ɑ-metildihidrotestosterona en dos variedades comerciales de tilapia del Nilo. Con base en lo anterior, el objetivo del presente trabajo fue evaluar el porcentaje de masculinización y el crecimiento obtenido en una variedad altamente domesticada de tilapia del Nilo, alimentada durante diferente número de días con alimento comercial adicionado con 17ɑ-metiltestosterona.
Materiales y métodos
Ubicación del experimento
El trabajo de investigación se llevó a cabo en las instalaciones del Laboratorio acuícola y de la Unidad Experimental de Producción Acuícola (UEPA), ambos de la Universidad del Papaloapan (UNPA) campus Loma Bonita, Oaxaca, localizado en las coordenadas 18o06’ LN y 95o53’ LO, a una altura de 30 msnm.
Alevines
Se utilizaron alevines de tilapia del Nilo variedad Spring de cinco días de edad (dde), provenientes de un laboratorio comercial productor de alevín, ubicado en el distrito de Tuxtepec, Oaxaca. Los alevines se trasladaron al Laboratorio de Acuicultura de la UNPA en una bolsa de plástico transparente con oxigenación. A su llegada se les aplicó un tratamiento con sal (> 35 ups) durante 10 min para evitar la introducción de parásitos externos al sistema. De igual forma, se evitaron cambios bruscos de temperatura (> 1 oC).
Alimento hormonado
Para realizar la masculinización se utilizó alimento hormonado comercial tipo harina (El Pedregal® Silver Cup, 60 mg 17ɑ-metiltestosterona), mientras que para el grupo control se utilizó alimento tipo harina del mismo diámetro pero sin hormona adicionada.
Diseño experimental
Se evaluaron cuatro tiempos de aplicación (10, 15, 20 y 25 días) del alimento hormonado más un control (0), en una población mixta (hembras y machos) de tilapia del Nilo variedad Spring. Los tratamientos fueron designados H0, H10, H15, H20 y H25, indicando el tiempo que fueron alimentados con la hormona (H). Se utilizaron tres réplicas por tratamiento distribuidas de manera aleatoria dentro del sistema experimental. Para el experimento se empleó un sistema de recirculación compuesto por 15 acuarios de acrílico (tres acuarios por cada tratamiento) con una capacidad de 85 L cada uno, con dos filtros, uno mecánico y un bio-filtro compuesto por bio-bolas. El experimento consistió de 30 días de cultivo en acuarios y otros 20 días de cultivo en estanques exteriores de 1000 L de capacidad.
Siembra
Los alevines se sembraron a una densidad inicial de 0.5 alevines L-1 (43 alevines por acuario), con un fotoperiodo de 12 luz: 12 oscuridad. Se registró la temperatura del agua tres veces al día utilizando un termómetro analógico (± 2 oC, Taylor®) para determinar la temperatura promedio diaria, con el fin de mantenerla en un rango de 26-27 oC.
Alimentación de los alevines
Los alevines del grupo control y los tratamientos (H10, H15, H20 y H25) se alimentaron inicialmente con 0.15 g de alimento cada dos horas (8:00, 10:00, 12:00, 14:00, 16:00 y 18:00). A partir de la segunda biometría, se hizo un ajuste con base en la biomasa obtenida y se proporcionó una ración del 10% de la biomasa total por réplica repartida en seis dosis diarias. El flujo de agua y aire dentro de los acuarios se cerró 10 min antes de alimentar a los alevines y 15 minutos después de alimentarlos, para facilitar el consumo de alimento. Se realizaron sifoneos y recambios de agua (~20%) diarios para extraer los residuos del alimento no ingerido, las heces y los alevines muertos. La concentración de amonio se evaluó una vez por semana.
Para realizar las biometrías se colectó una muestra al azar del 25% de los peces por réplica de cada tratamiento y se registró el peso húmedo (PH) individual con una balanza digital (Ohaus® ± 0.01). La longitud total (LT) se obtuvo mediante una imagen digital usando el software ImageJ® (versión 1.36). Las biometrías se realizaron cada 10 días a partir del día de siembra hasta terminar el experimento (50 días). Al finalizar el periodo de alevín (30 días) se procedió a realizar el cambio de alimento. A ambos grupos se les suministró por el resto del experimento (20 días) alimento tipo migaja (53% proteína, Purina® Agribrands).
Análisis de proporción de sexos
Para la determinación de sexos en cada uno de los tratamientos se empleó la técnica de squash propuesta por Guerrero y Shelton (1974), la cual consiste en extraer la gónada del juvenil para hacer una fijación en un porta-objetos en el cual se coloca la gónada, se le agregan un par de gotas de acetocarmin (HYCEL®), para posteriormente macerar con ayuda de un cubre-objetos de tal modo que la gónada se rompe permitiendo observar su estructura interna.
Evaluación del crecimiento
La evaluación del crecimiento se realizó a partir del peso húmedo y longitud total obtenidas a partir de cada biometría. Los índices obtenidos en el presente experimento fueron:
Biomasa ganada en g:
Tasa de crecimiento diario:
Factor condición:
Evaluación del factor de conversión alimenticia
El factor de conversión alimenticia (FCA) se obtuvo con la siguiente formula:
Supervivencia
El porcentaje de supervivencia (S) se obtuvo al final del experimento mediante la fórmula:
Análisis estadísticos
Se verificaron los supuestos de normalidad (Kolmogorov-Smirnov) y homocedasticidad (Levene). Las diferencias en peso húmedo, longitud total, así como los índices obtenidos (BG, TCE, FCA, FC) se evaluaron mediante un análisis de varianza de una vía. Las diferencias entre tratamientos se compararon mediante la prueba de Tukey a un nivel de significancia del 95%. La proporción de machos identificados en cada tratamiento se compararon contra la proporción de machos (1 macho: 1 hembra) esperada en desoves normales para tilapia del Nilo mediante una prueba de chi-cuadrada a un nivel de significancia del 99%. El porcentaje de supervivencia final entre tratamientos se transformó mediante la función arco-seno y se comparó mediante un análisis de varianza de una vía. Todos los análisis se realizaron con el programa STATISTICA V. 8.
Resultados
Los resultados obtenidos para el PH y LT durante el periodo de alevín se encuentran en la Figura 1 (A peso húmedo, B longitud total). Se observaron diferencias significativas en el PH a partir del día 20, con los grupos H20 y H25, presentando los valores significativamente más altos (p < 0.05) en comparación con los grupos H10 y H0. Para el día 30 el grupo H25 registró los valores significativamente más altos (p < 0.05) en comparación con los valores observados en los grupos H15 y H0. No se observaron diferencias significativas (p > 0.05) entre los grupos H10 y H20. Con respecto a la LT, para el día 20 se encontraron diferencias significativas (p < 0.05) entre los grupos H20 y H25 con respecto a los grupos H10 y H0, registrando los primeros los mayores valores. Al día 30 se registraron los valores de longitud total significativamente mayores (p < 0.05) en el grupo H25 en comparación con los grupos H0 y H15. Por último, tanto a los 20 como a los 30 días el grupo H0 mostró los valores de PH y LT significativamente más bajos (p < 0.05) en comparación con los grupos que consumieron el esteroide.
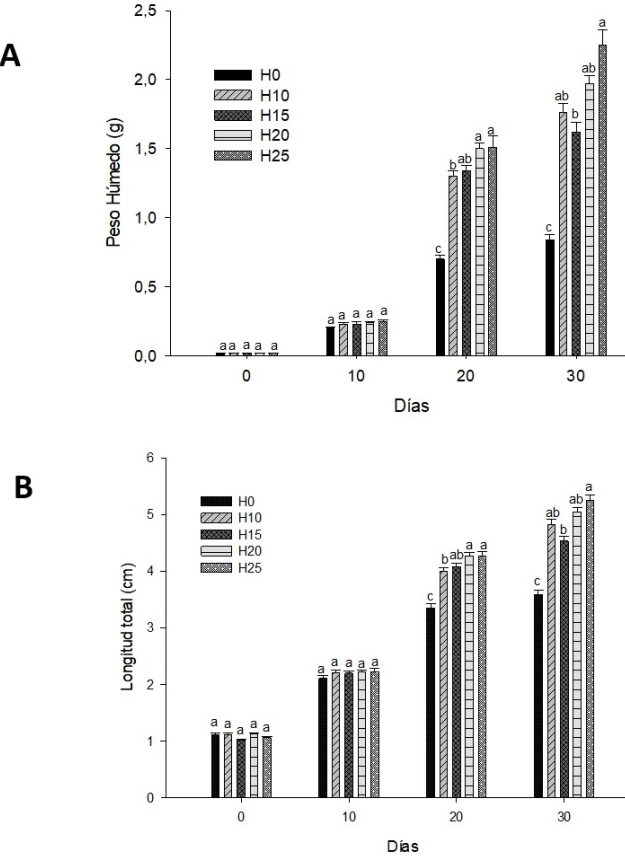
Figura 1: A.-Peso húmedo, B.- Longitud total obtenida durante el periodo de alevín en la tilapia del Nilo. La presencia de superíndices con diferente letra sobre las columnas indica diferencias significativas entre tratamientos para cada biometría (p < 0.05)
Los resultados del PH y LT durante el periodo juvenil se presentan en la Figura 2 (A Peso húmedo, B Longitud total). El incremento en PH registró diferencias significativas (p < 0.05) a los 40 y 50 días, con el grupo H25 registrando los mayores valores en comparación con el grupo H0, H10 y H15. No se observaron diferencias significativas (p > 0.05) entre el grupo H25 y el grupo H20. El grupo H0 registró los valores significativamente (p > 0.05) más bajos de PH durante el periodo en comparación con los grupos alimentados con el esteroide.
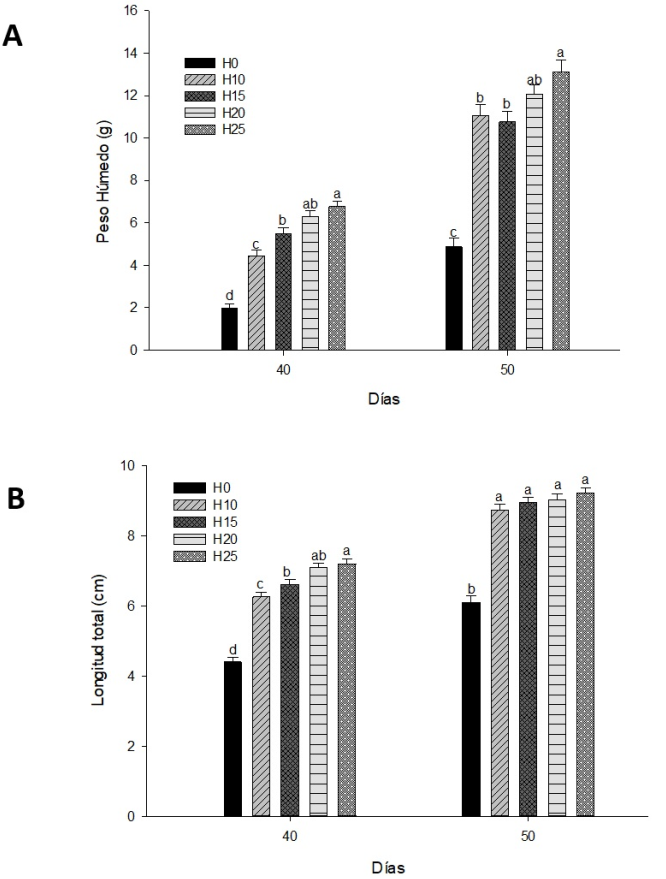
Figura 2: A.-Peso húmedo, B.-Longitud total obtenida durante el periodo de juvenil en la tilapia del Nilo. La presencia de superíndices con diferente letra sobre las columnas indica diferencias significativas entre tratamientos para cada biometría (p < 0.05).
La LT, al igual que el PH, registró diferencias significativas (p < 0.05) a los 40 y 50 días. Entre los grupos masculinizados, a los 40 días el grupo H25 registró los valores significativamente (p < 0.05) más elevados en comparación con grupos H10 y H15, mientras que a los 50 días no se observaron diferencias significativas (p > 0.05) entre los grupos alimentados con el esteroide (Figura 2B Longitud total). El grupo H0 registró los valores significativamente (p > 0.05) más bajos de LT durante el periodo en comparación con los grupos alimentados con el esteroide.
Los resultados obtenidos para los índices de crecimiento se presentan en la Tabla 1. No se observaron diferencias significativas (p > 0.05) entre grupos para ninguno de los cuatro índices a los 10 días.
Tabla 1: Biomasa ganada (BG), tasa de crecimiento diario (TCD), factor de conversión alimenticia (FCA) y factor de condición (FC) obtenidos en la tilapia del Nilo (Oreochromis niloticus) alimentada durante diferentes tiempos con alimento comercial adicionado con el esteroide 17ɑ-metiltestosterona durante el periodo de alevín. Promedio ± error estándar.
| Tratamientos | |||||
| Índice | H0 | H10 | H15 | H20 | H25 |
| Día 10 | |||||
| BG | 7.90 ± 0.55a | 10.35 ± 0.90a | 9.88 ± 1.13a | 10.82 ± 0.16a | 10.75 ± 0.46a |
| TCD | 0.008 ± 0.001a | 0.009 ± 0.001a | 0.009 ± 0.001a | 0.009 ± 0.001a | 0.009 ± 0.001a |
| FCA | 1.06 ± 0.03a | 1.05 ± 0.02a | 1.05 ± 0.02a | 1.02 ± 0.03a | 1.04 ± 0.02a |
| FC | 2.29 ± 0.17a | 2.21 ± 0.15a | 2.20 ± 0.17a | 2.37 ± 0.19a | 2.46 ± 0.23a |
| Día 20 | |||||
| BG | 21.84 ± 1.77b | 51.54 ± 1.46a | 49.69 ± 3.26a | 57.25 ± 2.92a | 58.15 ± 7.33a |
| TCD | 0.020 ± 0.001b | 0.043 ± 0.002a | 0.046 ± 0.002a | 0.050 ± 0.002a | 0.051 ± 0.003a |
| FCA | 1.42 ± 0.04a | 1.20 ± 0.01b | 1.21 ± 0.03b | 1.19 ± 0.01b | 1.20 ± 0.04b |
| FC | 2.01 ± 0.19a | 2.12 ± 0.12a | 2.10 ± 0.14a | 2.03 ± 0.13a | 2.07 ± 0.17a |
| Día 30 | |||||
| BG | 31.11 ± 4.91c | 79.46 ± 6.51b | 88.77 ± 4.02b | 102.92 ± 6.97b | 151.34 ± 5.76a |
| TCD | 0.028 ± 0.004c | 0.071 ± 0.005b | 0.080 ± 0.006b | 0.092 ± 0.006b | 0.135 ± 0.010a |
| FCA | 1.91 ± 0.13a | 1.61 ± 0.09ab | 1.44 ± 0.03bc | 1.45 ± 0.05b | 1.23 ± 0.06c |
| FC | 2.63 ± 0.25a | 1.93 ± 0.14b | 1.96 ± 0.15b | 1.81 ± 0.07b | 1.96 ± 0.15b |
| Día 40 | |||||
| BG | 12.24 ± 4.93b | 43.79 ± 11.79ab | 65.61 ± 19.16ab | 84.2 ± 7.86a | 60.42 ± 15.38ab |
| TCD | 0.023 ± 0.007b | 0.055 ± 0.012ab | 0.086 ± 0.014a | 0.100 ± 0.011a | 0.076 ± 0.013a |
| FCA | 3.38 ± 1.53a | 2.36 ± 0.55a | 1.97 ± 0.48a | 1.62 ± 0.05a | 2.53 ± 0.56a |
| FC | 2.76 ± 0.39a | 1.95 ± 0.16ab | 2.06 ± 0.16ab | 1.84 ± 0.12b | 1.91 ± 0.14ab |
| Día 50 | |||||
| BG | 60.94 ± 11.79b | 212.96 ± 37.63a | 166.81 ± 17.79ab | 194.74 ± 29.41a | 200.87 ± 34.27a |
| TCD | 0.116 ± 0.016b | 0.265 ± 0.021a | 0.210 ± 0.02a | 0.230 ± 0.019a | 0.254 ± 0.023a |
| FCA | 1.35 ± 0.08a | 1.39 ± 0.15a | 1.65 ± 0.09a | 1.72 ± 0.20a | 1.71 ± 0.18a |
| FC | 2.58 ± 0.33a | 1.76 ± 0.14b | 1.56 ± 0.10b | 1.78 ± 0.16b | 1.72 ± 0.09b |
H0 = control, H10 = alimentado 10 días con el esteroide, H15 = alimentado 15 días con el esteroide, H20 = alimentado 20 días con el esteroide, H25 = Alimentado 25 días con el esteroide. Valores marcados con su- períndices con distinta letra dentro del mismo renglón indican diferencias significativas (p < 0.05).
Los resultados obtenidos para BG y TCD registraron diferencias significativas (p < 0.05) a los 20 y 30 días, el grupo H0 mostró los valores el más bajos en comparación con los grupos tratados con el esteroide. Dentro de los grupos tratados no se registraron diferencias significativas (p > 0.05) en ninguno de los grupos a los 20 días, mientras que a los 30 días el grupo H25 mostró los valores significativamente más altos en comparación con el resto de los grupos tratados. En lo que respecta al FCA, el grupo H0 registró los valores significativamente (p < 0.05) mayores a los 20 y 30 días en comparación con los grupos tratados con el esteroide. Entre los grupos tratados, no se registraron diferencias significativas a los 20 días, mientras que a los 30 días el grupo H25 registró un valor significativamente (p < 0.05) más bajo en comparación con los grupos H10 y H20. Por último, el FC no mostró diferencias significativas a los 20 días entre el grupo H0 y los grupos tratados con el esteroide. Pero a los 30 días el valor observado en el grupo H0 fue significativamente (p < 0.05) mayor en comparación con los grupos alimentados con el esteroide. No se observaron diferencias significativas (p > 0.05) entre ninguno de los grupos alimentados con el esteroide a 20 o 30 días (Tabla 1).
A los 40 días, el grupo H0 mostró un valor de BG significativamente (p < 0.05) más bajo en comparación con el observado en el grupo H20, mientras que para el final del experimento (50 días), fue significativamente (p < 0.05) más bajo en comparación a los grupos H20 y H25. Al igual que la BG, la TCD mostró valores significativamente (p < 0.05) más bajos para el grupo H0, tanto a 40 como a 50 días, en comparación con los grupos alimentados con el esteroide. Entre los grupos tratados no se observaron diferencias significativas (p < 0.05) en la TCD a 40 o 50 días.
En lo que respecta al FCA no se observaron diferencias significativas entre ninguno de los grupos analizados a 40 o 50 días. Por último, el valor del FC observado para el grupo H0 fue significativamente (p < 0.05) más alto a los 40 días en comparación con el grupo H20, mientras que para el final del experimento, el FC del grupo H0 mostró un valor significativamente (p < 0.05) más alto en comparación con los grupos alimentados con el esteroide. No se registraron diferencias significativas (p > 0.05) entre los grupos tratados a 40 o 50 días (Tabla 1).
Los resultados obtenidos de la proporción de sexos y supervivencia final se presentan en el Tabla 2. Se observó una proporción de machos significativamente (p < 0.001) más alta en todos los grupos que recibieron el esteroide, en comparación con la proporción de machos esperada para un desove de tilapia del Nilo sin masculinizar (50% de machos). En lo que respecta a la supervivencia final, se obtuvo un valor significativamente (p < 0.05) más alto para el grupo H20 en comparación con el grupo control. No se observaron diferencias significativas (p > 0.05) entre los grupos alimentados con el esteroide.
Tabla 2: Proporción de sexos y porcentaje de supervivencia final (SF) en juveniles de tilapia del Nilo (Oreochromis niloticus) alimentados durante diferentes tiempos con alimento comercial adicionado con el esteroide 17ɑ-metiltestosterona durante el periodo de alevín. Promedio ± error estándar. n = 3.
| Tratamiento | NPA* | Proporción de sexos (%) | SF (%) | |
| Machos | Hembras | |||
| H0 | 30 | 60 | 40 | 81.1 ± 4.20b |
| H10 | 30 | 1001 | 0 | 93.3 ± 1.28ab |
| H15 | 30 | 1001 | 0 | 93.2 ± 3.94ab |
| H20 | 30 | 971 | 3 | 97.7 ± 2.27a |
| H25 | 30 | 1001 | 0 | 92.6 ± 4.12ab |
*NPA = número de peces analizados. 1 Significativamente diferente (p < 0.001) de la proporción de machos esperada. Valores marcados con superíndices con distinta letra dentro de la misma columna indican diferencias significativas (p < 0.05).
Discusión
La edad de los alevines utilizados garantiza que la aplicación del esteroide tuvo un nivel óptimo en la sangre antes de llegar a los 10 días de edad. Al respecto, Vera-Cruz et al. (1996) recomiendan que los alevines deben tener un tamaño menor a 10 mm antes de iniciar el proceso de masculinización, para asegurar porcentajes cercanos al 100%, mientras que Melard y Philippart (1995) sugieren alimentar a los alevines por 12 horas para mejorar la asimilación del esteroide. En el caso de la tilapia del Nilo, Hiott y Phelps (1993) evaluaron aspectos relacionados con la talla de inicio y el éxito en la reversión sexual, llegando a la conclusión que la talla óptima para asegurar un porcentaje cercano al 100% de masculinización es menor a los 11 mm. En el presente trabajo los alevines utilizados tuvieron una talla inicial de 10 mm y se alimentaron seis veces al día por 12 horas, lo que influyó en una masculinización del 100% en el tratamiento H10.
El andrógeno utilizado mostró efectos significativos sobre la composición sexual de las poblaciones de peces tratados. El porcentaje de eficiencia logrado con el método de reversión oral en los grupos alimentados a diferentes tiempos se puede considerar exitoso, ya que de acuerdo a Phelps (2001) una masculinización exitosa mediante andrógenos debe tener un porcentaje de machos mínimo del 97%. Lo que confirma que para tener una reversión sexual exitosa se debe abarcar solo el periodo lábil de la especie, el cual en el caso de la tilapia del Nilo se presenta alrededor de los cinco (en hembras) a seis (en machos) días después de la eclosión (Ijiri et al. 2008, Tao et al. 2013). Los elevados niveles de 17a-metiltestosterona (MT) mantenidos durante los primeros 10 días de permitieron obtener 100% de machos, lo que indica que esta duración (para el tratamiento hormonal) es suficiente para suprimir la acción el gen cyp19a (responsable de producir la enzima aromatasa, la cual permite la diferenciación ovárica), mediante la estimulación del gen dmrt1 que está involucrado en la diferenciación testicular (Bhandari et al. 2006, Ijiri et al. 2008). En este sentido, Tao et al. (2013) reportan a los cinco días de la presencia en gónadas XX (hembras) de receptores de andrógenos (ar1 y ar2), los cuales se unen al esteroide suministrado oralmente, favoreciendo el proceso de masculinización. Los resultados encontrados concuerdan con lo reportado por Vinarukwong et al. (2018)) en dos variedades domesticadas de tilapia del Nilo cultivadas en Tailandia, donde con solo 15 días de tratamiento tuvieron 100% de masculinización. En este caso, la duración de 5 y 10 días, tuvieron entre el 82 y 90% de masculinización, respectivamente. La variación en los porcentajes obtenidos probablemente obedece a las variedades utilizadas y al esteroide empleado, ya que en este caso se utilizó 17ɑ-metildihidrotestosterona a concentración de 80 mg por kg de alimento. Los resultados obtenidos indican que hormonar de 28 a 40 días no tiene efecto significativo en la proporción de machos obtenidos. El 3% de hembras observado en el tratamiento con la aplicación de MT por 20 días (H20), probablemente fue causado por factores genéticos parentales (Alcántar-Vázquez et al. 2015), como por ejemplo, una insensibilidad genética al efecto de la hormona, o bien al muestreo realizado.
La razón de un periodo de masculinización largo en el cultivo comercial de la tilapia del Nilo, se puede deber a que por lo general los desoves provienen de decenas o cientos de hembras, lo cual puede representar una alta variación en edades (de horas a días) de los alevines que entran al pro ceso de reversión sexual. Al respecto, la mayoría de los estudios donde se establece el periodo de masculinización óptimo para esta especie son mayores a 25 años (Shelton et al. 1981, Hiott y Phelps 1993), lo cual no toma en cuenta la caracterización precisa a nivel genético del desarrollo embrionario y larval de la tilapia del Nilo (Ijiri et al. 2008, Baroiller et al. 2009, Tao et al. 2013, Gennotte et al. 2014), lo que ha permitido determinar los periodos de desarrollo claves para una reversión sexual exitosa, y la extensiva selección artificial y domesticación a la que ha sido sometida la especie en las últimas décadas, lo que ha permitido la aparición de un gran número de variedades genéticas altamente específicas para diferentes requerimientos de cultivo (Fitzsimmons et al. 2011). Por lo general, estas nuevas variedades o líneas genéticas son desarrolladas a partir de pocas hembras reproductoras, lo que permite uniformizar su comportamiento social y reproductivo. Si a lo anterior, se le suma la optimización a nivel comercial de los procesos de manejo y reversión sexual, es factible reducir el periodo de masculinización ajustándolo al conocimiento actual y a la domesticación reproductiva de la tilapia del Nilo. El hecho de que el desove utilizado en el presente trabajo fuera adquirido a un laboratorio comercial en lugar de uno generado en nuestras instalaciones con reproductores experimentales, apoya la idea de que es factible reducir el tiempo necesario para alcan zar una masculinización del 100% en los laboratorios comerciales del país, los cuales utilizan variedades genéticas altamente domesticadas. Ha este respecto, Osure y Phelps (2006) reportan diferencias significativas en aspectos reproductivos y de crecimiento en cuatro variedades genéticas de tilapia del Nilo con diferente grado de domesticación.
Se sabe que temperaturas mayores de 34 oC tienen un efecto masculinizante en la tilapia del Nilo. En este sentido, Wang y Tsai (2000) reportan que la exposición de alevines de tilapia del Mozambique de 10 días de edad a temperaturas de 32-34 oC puede en algunos casos inducir masculinización gonadal e incrementar la proporción de machos. Lo anterior, es posible porque la determinación del sexo fenotípico en varias especies de tilapia, incluida tilapia del Nilo es un rasgo complejo, el cual no solo es determinado por factores genéticos primarios (locus XX/XY); sino que es producto de la interacción de estos factores genéticos primarios, con factores genéticos se cundarios (efectos parentales) y factores ambientales (Wang y Tsai 2000, Baroiller et al. 2009), en espe cial la temperatura del agua durante el periodo de alevín. Trabajos previos han observado una tendencia genética hacia la masculinización de las gónadas a temperaturas moderadamente elevadas (28 ± 1 oC), lo cual ha resultado en una proporción de machos elevada en varios grupos durante el proceso de reversión sexual (Wessels y Hörstgen-Schwark 2007, Alcántar-Vázquez et al. 2014b). En el presente tra bajo la temperatura del agua se mantuvo entre 26 y 27 °C durante todo el tratamiento con MT con la intención de reducir el posible efecto de la temperatura del agua en la proporción de machos obtenida, para que esta manera los porcentajes de machos observados fueran el resultado del efecto del esteroide suministrado durante el periodo de alevín y no de la interacción de la gónada en diferenciación con la temperatura del agua. En este sentido, es importante mencionar que aunque el grupo control registró un porcentaje de machos del 60%, en lugar del 50% esperado, este tipo de porcentajes son normalmente observados en la tilapia del Nilo y son atribuidos a factores genéticos parentales (producto de la cruza de machos y hembras normales) (Baroiller et al. 2009) y no al efecto de la temperatura del agua durante el periodo lábil.
Al término del experimento se obtuvo un crecimiento significativamente superior en los grupos tratados con MT en comparación con grupo control, lo cual apoya lo observado por diversos autores en varias especies de tilapia (Hafeez-ur-Rehman et al. 2014, Ajiboye et al. 2015, Rima et al. 2017, Singh et al. 2018, Chávez-García et al. 2020). Sin embargo, no todos los autores reportan diferencias en el crecimiento (Jiménez y Arredondo 2000, Torres-Hernández et al. 2010, Vinarukwong et al. 2018), especialmente al utilizar otro esteroide. El efecto anabólico resultante del uso de andrógenos en la masculinización de la tilapia del Nilo parece ser casi exclusivo de la MT, sin embargo, en algunos casos puede ser difícil de identificar. La mayoría de los estudios que han analizado mejoras en el crecimiento de los peces revertidos sexualmente en comparación con los no tratados, lo hacen comparando el crecimiento de las poblaciones compuestas exclusivamente por peces machos contra una población sexual mixta, en la cual las hembras presentes muestran una tasa de crecimiento menor. En el presente trabajo, en el grupo control se obtuvo un 40% de hembras, siendo este el grupo de los peces más pequeños al final del experimento. Lo anterior, puede ser explicado por la mayor proporción de hembras, que crecen más lento y son menos agresivas a la hora de alimentarse. Al respecto, Phelps y Popma (2000) mencionan que cualquier crecimiento mejorado de la tilapia tratada con andrógenos está más relacionado con el crecimiento superior de los machos que con la respuesta anabólica relacionada con el aumento de la síntesis de proteínas y el aumento de musculo. Mientras que Phumyu et al. (2012) reportan que en la tilapia del Nilo los peces revertidos con MT muestran un crecimiento similar al de los machos, sin embargo, las hembras alimentadas con el alimento adicionado con el esteroide muestran un mejor crecimiento y una mejor eficiencia alimenticia, lo cual se atribuye a características del tracto gastrointestinal.
A pesar del número de hembras observado en el grupo control, el número de días que se proporcionó la MT tuvo un efecto en el crecimiento, ya que los grupos que recibieron el esteroide por 20 y 25 días fueron los que presentaron un mejor crecimiento durante el proceso de masculinización y al finalizar el experimento. Lo que indica que el efecto del esteroide sobre el metabolismo del pez puede durar por varias semanas, incluso meses después de terminado el periodo de reversión sexual, ya que trabajos realizados con MT parecen confirmar lo anterior (Hafeez-ur-Rehman et al. 2014, Ajiboye et al. 2015, Singh etal. 2018, Chávez-García et al. 2020). Los resultados obtenidos para la BG y TCD apoyan la idea de que el efecto anabólico del esteroide pudo haber contribuido en los resultados observados, optimizando la absorción de nutrientes que se reflejó en un crecimiento somático mejorado. Aunque es posible que la diferencia en la tasa de crecimiento entre sexos, reportada para la tilapia del Nilo, pueda ser responsable de las diferencias observadas. En el presente trabajo, al finalizar el experimento la ausencia de un crecimiento gonadal que se observó al extraer las gónadas para realizar la técnica de squash, indica que la maduración sexual no se había presentado y por lo tanto los esteroides sexuales producidos natu ralmente (endógenos), que son los que permiten en gran medida las diferencias entre sexos, no se encontraban presentes en cantidad suficiente en la sangre como para ser un factor importante.
El factor de condición (FC) puede indicar el estado nutritivo de los organismos, definido por la cantidad de energía poseída (contenido de grasa disponible) en los peces para llevar a cabo varias funciones, incluido el crecimiento y la reproducción (Neff y Cargnelli 2004, Gupta et al. 2012). El factor de condición también se puede usar para evaluar el estado fisiológico bajo un estresante potencial (Cifuentes et al. 2012), como lo es un alevín de tilapia del Nilo alimentado con un esteroide sexual. Se observó en general un FC más bajo en los peces de grupos tratados con MT en comparación con los peces del grupo control. Al respecto, Cifuentes et al. (2012) mencionan que esta disminución podría interpretarse como el agotamiento de las reservas de energía almacenadas como glucógeno del hígado o grasa corporal. El rápido crecimiento de los alevines masculinizados en comparación con los del grupo control podría explicar porque la grasa corporal (energía de reserva) era menor en estos grupos y por lo tanto el FC. Para la tilapia del Nilo y sus híbridos bajo un desarrollo óptimo se han reportado valores cercanos a 2.0 (Fish Breeding Association 2003, El-Saidy y Gaber 2005, Gupta et al. 2012), similares a los observados en el presente experimento, lo que indica el buen desarrollo de los tratamientos evaluados.
La supervivencia obtenida al final de un experimento de masculinización depende de factores como la especie, la concentración del esteroide, el tiempo de inicio y la duración del tratamiento (Piferrer 2001). Los cuales pueden influir en la mortalidad durante la etapa de reversión sexual, después de que termine la reversión y hasta al final del experimento. Una de las razones por lo cual la MT es tan utilizada, además de sus altas tasas de reversión sexual, es que por lo general no ocasiona efectos negativos en la supervivencia (El-Greisy y El-Gamal 2012, Rima et al. 2017, Singh et al. 2018) si se emplea en concentraciones menores a 60 mg Kg-1 (El-Greisy y El-Gamal 2012, Jensi et al. 2016). En nuestro caso, la supervivencia final observada fue más alta en los grupos expuestos al esteroide en comparación con el grupo control. Pero la mortalidad observada en el grupo control probablemente este relacionada más al manejo que a un efecto positivo de la MT, ya que para el experimento se implementó un protocolo de cultivo que involucró 30 días de cultivo en acuarios de 85 L y otros 20 días de cultivo en estanques exteriores de 1000 L de capacidad. El traslado de los alevines y el cambio de alimento al final del periodo de alevín, no fueron bien aceptados por el grupo control, probablemente por el menor tamaño en comparación con los grupos alimentados con el esteroide, lo cual derivó en una mayor mortalidad durante los días que siguieron a los cambios mencionados. Lo que se puede apoyar en el hecho de que al final del periodo de alevín, previo al cambio de estanque y de dieta, la supervivencia del grupo control se mantuvo por encima del 90%, al igual que la de los grupos tratados con MT. En experimentos futuros será necesario optimizar el protocolo de manejo y cultivo, para reducir la probabilidad de una mortalidad como la observada en el grupo control.
Conclusiones
Los resultados indican que un tratamiento con 10 días es suficiente para inducir una reversión del 100% en variedades domesticadas de tilapia del Nilo como lo es la variedad Spring. La reducción de la duración del periodo de masculinización podría ayudar a reducir el uso de hormonas exógenas en el cultivo comercial y reducir su presencia en los efluentes de las granjas productoras de alevín. Sin embargo, el esteroide suministrado tiene un efecto anabólico positivo sobre los parámetros de crecimiento de los peces tratados, el cual perdura por semanas después de acabado el tratamiento, lo cual podría ser un incentivo para que los laboratorios productores de alevín masculinizado ofrecieran el esteroide por mayor tiempo.

