











 text new page (beta)
text new page (beta) English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkBiodiversity is perhaps the most valuable asset on planet Earth. As such, conservation and research efforts are usually devoted to sites with outstanding species richness (Myers et al. 2000). Among terrestrial ecosystems, tropical rainforests have become an icon of biodiversity, and for good reasons: they not only harbor a huge fraction of the species on the planet, but also the largest number of species per square kilometer (Dirzo & Raven 2003). However, as it happens so frequently in ecology, everything is a matter of scale. If the area over which the number of plant species is recorded becomes small, tropical rainforests retain their preeminence down to 100 m2. At smaller scales, grasslands become the undisputed holders of plant-richness world records. The figures are surprising: grasslands may accommodate up to 89 plant species in a square meter, or even 5 species in a square centimeter (Wilson et al. 2012). One outstanding attribute of all record-holding grasslands is that they have been subjected to millennia of chronic disturbance by mowing or ungulate grazing (Wilson et al. 2012).
Species-rich communities are an endless source of awe and perplexity for biologists. Early studies on species interactions made it clear that species coexistence can only occur under restrictive conditions. Niche differentiation has been an all-time favorite hypothesis to explain coexistence, and much of the recent evidence seems to favor this proposition (Levine & HilleRisLambers 2009, Adler et al. 2010, Chu & Adler 2015). However, this raises the question of how there can be so many niches if plants depend on so few resources (Silvertown 2004).
The classic theory of niche-based coexistence suggests that there should be a “hard limit” to richness determined by the similarity of the species requirements and how finely niche space can be partitioned (e.g., Preston, 1962). Communities where richness matches that limit are called saturated. Alternative explanations for coexistence do not acknowledge such limit. This is the case of the hypothesis that local richness is largely determined by regional diversity, or of theories in which niche similarity does not necessarily preclude coexistence such as neutrality or Chesson’s coexistence theory (Chesson, 2000; Hubbell, 2001).
Thus, determining whether communities are saturated, or whether a hard limit to richness exists, is an important task to screen among different models of species coexistence. Keeping the world records of plant richness is a relevant contribution in this direction. Wilson et al. (2012) compiled such information for different spatial scales. They found that the function relating the records to area fits Preston’s (1962) canonical theory, which was developed on the assumption that communities are saturated. As they point out, if there are in fact saturated communities in nature, these are the ones in their database.
The known records will surely be beaten because it is highly unlikely that the richest places on Earth have been already surveyed (especially if we consider small scales). However, based on the close relationship between richness records and area, Wilson et al. propose that any new records should be near the current ones. Here we test this hypothesis, and analyze the role of chronic disturbance in promoting extremely high plant diversity in a species-rich grassland.
Materials and Methods
We worked in a semiarid grassland in Concepción Buenavista, Oaxaca, Mexico (Figure 1a). The climate is semiarid, with an annual precipitation of 578 mm and an annual mean temperature of 16.3 °C (Martorell & Martínez-López 2014). One of the outstanding features of this community is that it develops on very thin (usually < 15 cm) soils over a relatively impermeable tuff deposit. This community is probably the southernmost xeric grassland of North America, and is dominated by the same species or genera that characterize many of its northern counterparts (cf. Cruz-Cisneros & Rzedowski, 1980). According to Mueller-Dumbois & Ellenberg (1974) classification, the vegetation is a shortgrass prairie. We still do not know the total plant richness of this community. Our current list exceeds 200 species, but it is by no means exhaustive. We have found new species in our permanent plots almost every year since 2001 (For a sample of the community’s richness please visit http://conabio.inaturalist.org/projects/plantas-de-concepcion-buenavista-oaxaca-mexico).

Figure 1 Vegetation at Concepción Buenavista. a) Landscape during the flowering season. b) The 0.1 × 0.1 m quadrat with world record plant richness.
We recorded species richness in 21 0.5 ha sites occurring over a chronic anthropogenic disturbance gradient. Disturbance was measured through the Martorell & Peters (2009) indices, which estimate the intensity of three disturbance agents: livestock raising, human activities and land degradation. In eight 1 × 1 m randomly-placed plots at each site, we surveyed 20 randomly selected 0.1 × 0.1 m quadrats. We recorded the number of vascular plant species rooted in each of these quadrats.
The site with the largest richness had 94 species. This is not far from the 113 species recorded at Laelatu, Estonia, the site that holds the world record at the scale of 0.1 × 0.1 m (Kull & Zobel 1991). This suggested that our community was a candidate to test Wilson et al.’s proposition. Thus, in September 2014 we looked for the 0.1 × 0.1 m quadrat with the largest number of vascular plant species at one of the sites where we had recorded a large species richness in 2012. Only plants rooted inside the quadrat were recorded. Because our sampling was non-random and followed no systematic procedure, it cannot be strictly replicated. As a measure of sampling effort, we can only state that it comprised about 10 man-hours.
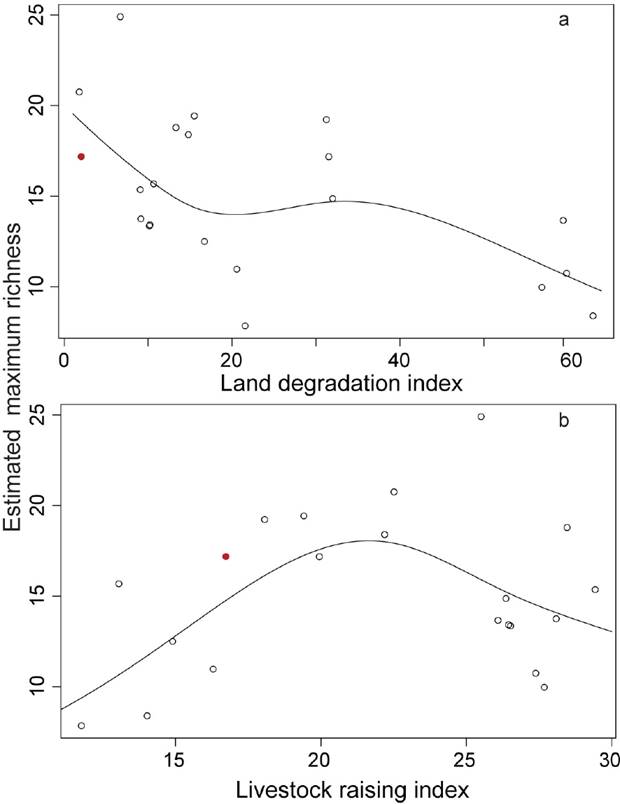
To analyze the effect of disturbance on promoting high species richness, we first needed an estimate of the maximum richness in each site. To calculate this figure, we first tried a number of probability distributions to describe the frequency of 0.1 × 0.1 m quadrats having different number of species. By means of the Akaike information criterion we determined that a gamma distribution performed best. We then estimated what the maximum observed richness at each site would be if we had sampled 1,000 0.1 × 0.1 m quadrats (i.e., 50 1 × 1 m plots) using the same protocol described above. To do so, we obtained the 99.9 percentile of the gamma distribution fitted for each site. This estimated maximum richness (EMR) was then regressed separately on the measurement of each disturbance agent using generalized additive models (Wood, 2011).
Results
We found a 0.1 × 0.1 m quadrat with 25 plant species (Figure 1b), meaning that the grassland of Concepción Buenavista currently shares the world record of plant richness with the grassland at Laelatu (Kull & Zobel, 1991). The identities of the species recorded in this quadrat are reported in Table 1. This figure was greater than the EMR for the site where we conducted the directed search (17.2 species), but was very similar to the EMR calculated for the richest site (24.9).
Table 1 Species composition of the richest quadrat in Concepción Buenavista, Oaxaca.
| Species | Family |
|---|---|
| Aristida adscensionis L. | Poaceae |
| Bouteloua chondrosioides (Kunth) Benth. ex S. Watson | Poaceae |
| Bouteloua hirsuta Lag. | Poaceae |
| Bouteloua scorpioides Lag. | Poaceae |
| Buchnera pusilla Kunth | Orobanchaceae |
| Bulbostylis tenuifolia (Rudge) J.F. Macbr. | Cyperaceae |
| Crusea diversifolia (Kunth) W.R. Anderson | Rubiaceae |
| Cyperus seslerioides Kunth | Cyperaceae |
| Dalea sericea Lag. | Fabaceae |
| Digitaria bicornis (Lam.) Roem. & Schult. | Poaceae |
| Drymaria leptophylla (Cham. & Schltdl.) Fenzl ex Rohrb. | Caryophyllaceae |
| Florestina pedata (Cav.) Cass. | Asteraceae |
| Helianthemum glomeratum (Lag.) Lag. ex Dunal | Cistaceae |
| Heterosperma pinnatum Cav. | Asteraceae |
| Microchloa kunthii Desv. | Poaceae |
| Muhlenbergia phalaroides (Kunth) P.M. Peterson | Poaceae |
| Oxalis lunulata Zucc. | Oxalidaceae |
| Plantago nivea Kunth | Plantaginaceae |
| Salvia axillaris Moc. & Sessé ex Benth. | Lamiaceae |
| Setaria parviflora (Poir.) Kerguelen | Poaceae |
| Stenandrium dulce (Cav.) Nees | Acanthaceae |
| Stevia ephemera Grashoff | Asteraceae |
| Tagetes micrantha Cav. | Asteraceae |
| Thymophylla aurantiaca (Brandegee) Rydb. | Asteraceae |
| Tridax coronopifolia (Kunth) Hemsl. | Asteraceae |
EMR diminished with land degradation (P = 0.035, Figure 2a), but reached its maximum at intermediate-high intensities of livestock raising (P = 0.028, Figure 2b). Human activities had no significant relation with maximum richness.
Discussion
Because we could not beat but only match the current record, our results support Wilson et al.’s conjecture that current estimations are close to the actual maxima. The intensity of disturbance caused by livestock and land degradation at the site where the record was found is close to that where the greatest EMR is expected. The maximum EMR is also close to the maximum richness that we observed. This suggests that the value that we observed is close to the actual maximum at the study grassland. We must bear in mind that our sampling was biased towards high-richness areas, while the previously reported records resulted from random sampling. Thus, a direct comparison cannot be made with other studies. Nevertheless, the findings reported in Wilson et al. (2012) are based on very heterogeneous methods and yet render a quite close relationship between area and record diversity. This suggests that sampling protocol may have a minor effect on maximum richness.
If future research confirms that the current records reflect more or less correctly what happens in nature, it would appear that there is a hard limit to species richness. Moreover, the species-area relationship found by Wilson et al. for such limit is quite strong (R2 = 0.976), so its confirmation would mean that there is something extremely predictable going on that calls for an explanation.
The relationship between disturbance and EMR is in line with what we know about individual-species responses to disturbance. Many of the species of our study grassland are known to increase in density with chronic disturbance (Martínez-Blancas 2014). Land degradation has a severe negative effect on the population density of several cacti, but many species are benefitted by livestock (Martorell & Peters 2009). Land degradation at the study site results in exposed tuft where very few species can grow, reducing overall richness. In contrast, livestock raising may resemble grazing by the now-extinct large herbivores that roamed Concepción Buenavista (Guerrero-Arenas et al. 2010), and to which local plant species may have been adapted.
The plant community of Concepción Buenavista shares several attributes with other record-holding sites. All of them are meso- to oligotrophic grasslands with chronic disturbances due to grazing, mowing or fire (Wilson et al. 2012). The soils at our site are not necessarily nutrient poor, but due to their low depth they cannot retain water for long periods. The currently scarce biomass of our grassland is insufficient to sustain wildfires, but cattle, sheep and goat grazing is intense at the site. It must be noted that livestock was introduced to Oaxaca early in the sixteenth century (García 1996, Romero-Frizzi 1996). Thus, our results agree with the proposal that resource limitation and long-term disturbance promote high diversity.
We hope that, by reporting this record, we also help to change the notion shared by many biologists that Mexican grasslands are secondary communities having a negligible biological value. The grassland of Concepción Buenavista is not only home to a remarkable plant richness, but also to two microendemic species: Mammillaria hernandezii and Sedum oteroi. Both are incapable of growing in wooded areas. Their endemicity and habitat specificity indicate the great antiquity of the grassland. The negative view of grasslands has resulted in a lack of protection and serious degradation. 95 % of Mexican arid grasslands are overgrazed (SEMARNAT, 2003). Concepción Buenavista is adjacent to a Biosphere Reserve, but its polygon twists around the grassland, leaving it outside the protected area. At the time the reserve was established, nobody spoke for herbs. If biodiversity captivates biologists, grasslands should not be disregarded.

