











 text new page (beta)
text new page (beta) English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkApproximately 85 % of Iran’s territory is composed of arid, semiarid or hyper arid environment (Heshmati 2007). In such a country, desert vegetation is an abundant type among the plant life spectra, representing many xerophytic plant species and communities. As species composition and diversity are fundamental characteristics of ecosystems (Chapin et al. 1992, Hooper & Vitousek 1997, Burk 2001), variations in diversity are presumably correlated with the stability of various biotic and abiotic components of ecosystems (Leigh Jr. 1965). Among different factors, availability of water is the primary one controlling the distribution of species (Noy-Meir 1973, Li et al. 2004). Water availability in such desert regions with rare occasional rainfall, is dependent to a high degree on water holding capacity, which in turn is highly related to some soil physical characteristics of texture coarseness. The importance of environmental characters on vegetation patterns has been emphasized by Jafari et al. (2004), El- & Amer (2003) and He et al. (2007).
Having an annual mean precipitation of 109.2 mm, with values less than 100 mm in some years, and a maximum temperature exceeding 48 ºC in summer months, the northern Badrud region, located in the central Iranian Plateau-Dasht-e-Kavir, represents a very hot dry environment. This ecosystem is typical of a desert with two major land forms, including a sand and gravel plain, which is partly covered with sand dunes. The region is floristically distinctive for possessing special psammophytic and halophytic vegetation types, many of which are endangered here in their habitats. This region is exposed to serious threats of vegetation degradation due to excessive drought, wind erosion, and overgrazing leading to successional progress of desertification.
Recognition of plant communities as well as investigating diversity and the effects of some abiotic factors such as gravel cover vs. ground bareness on species and diversity variables, by means of accurate multivariate techniques of analysis, are essential to stop further degradation of the natural vegetation of this area. The main objectives of this research are to: 1) investigate species composition, plant communities, and plant diversity of this region, 2) compare and contrast two vegetation classification techniques, Ward’s method and TWINSPAN, and test the results with ordination procedures; and 3) use multivariate analyses to examine the effect of some environmental factors on distribution, abundance, and patterns of communities.
The findings of this research related to species diversity potentials for such regions can improve conservation planning management and vegetation restoration. Since this is the first comprehensive study on vegetation analysis and flora for the region, the results can also be a monitoring point for future studies of species diversity and succession.
Materials and methods
Study area. The Badrud north region is located 65 km north of the Isfahan province in central Iran (Figure 1). It lies between 33º 44’ N, 51º 51’ E and 33º 48’ N, 51º 56’ E latitude and longitude. The study area covers 1,194 hectares, with the elevation ranging from 946 m in the plains to 1,021 m in the sand dunes. It is geologically composed of terraces, aeolian sand sheets, alluvial fans, and low to high aeolian sand hills belonging to Miocene, Pleistocene, Holocene, and recent ages. It has a desert climate representing very hot dry conditions. The annual mean precipitation is 109.2 mm and the annual mean temperature is 20.56 ºC, with extreme minimum and maximum temperatures of -18 ºC (in January) and 48.5 ºC (in July), respectively.

Vegetation sampling procedures. Data collection was carried out during five field trips between September 2008 and April 2010. A preliminary survey of vegetation types and the natural characteristics of the landforms guided the selection of three transect lines in different parts of the region, one in the plain and the other two in sand dunes. A total of sixty 1.5 × 2 m2 quadrats, 20 in each transect, were established using randomized systematic sampling. The size of sampling quadrats was determined using the minimal area method.
The presence of each species and its canopy cover in each quadrat were recorded using the visual estimation method and the ordinal scale of Domin (Kent & Coker 1992). Other variables for the percentage of gravel and ground bareness were also measured according to the quadrat area using the visual estimation method. Quadrat location, longitude, latitude, and altitude were also recorded by the use of a GPS receiver.
For each of the quadrats the Shannon-Wiener index of species diversity H´= ,i=1-s-pi log pi. (Magurran 1988) was calculated on the basis of the relative cover pi of the ith species. Shannon-Wiener is a widely used index for comparing diversity between various habitats. It counts all species according to their frequencies, and has the advantage of favoring neither rare nor common species disproportionately (Clarke & Warwick 2001). Effective number of species, (exp Shannon) was calculated in order to compare the Shannon diversity index of communities (Hill 1973).
Soil sampling and analysis. A random subsample of thirty soil samples were collected. Soils were taken from a 0-20 cm depth and sieved through a 2 mm screen. Samples were analyzed for soil moisture, electrical conductivity (EC), salinity, and acidity reaction (pH). A 100 g subset of each sample was oven-dried at 110 ºC for 24 h, and then weighed to determine its dry weight. Soil moisture was determined as a percentage of oven dried matter. A soil water extract (soil: water ratio = 1:5) was prepared. Electrical conductivity (EC), salinity, and pH of soil extracts were determined using a portable electrical conductivity meter and a glass electrode pH-meter (Consort 535). All of the procedures were performed according to McLean’s method (McLean 1982).
Data analysis method. The computer program PC-ORD 4.17 (McCune & Mefford 1999) was used for all classifications and ordinations. Two-way indicator species analysis (TWINSPAN) (Hill et al. 1975) and the cluster analysis using Ward’s minimum variance method (Ward Jr. 1963, Gauch Jr. 1982) were performed in order to classify the vegetation into groups based upon a relative cover percentage data matrix of 24 plant species in 52 quadrats (eight quadrats were eliminated from the data set due to the complete lack of vegetation cover). In TWINSPAN, pseudospecies cut levels were altered to 0, 2, 5, 10, 40, and 80, and the minimum group size for divisions to 6. Other program settings were retained as the defaults. A dendrogram was made based on the constructed two-way order table. Cluster analysis was performed using Ward’s method with Euclidean distance measure and the TWINSPAN groups superimposed. The results were compared with each other.
Robustness of classification was examined through ordination techniques including detrended correspondence analysis (DCA), non-metric multidimensional scaling (NMDS), and a grouping test procedure called multi response permutation procedure (MRPP). In DCA (Hill & Gauch Jr. 1980), rare species were down weighted and the defaults for rescaling axis were retained. The gradient length in standard deviation units and the magnitude of change in species composition were investigated. The NMDS method (Anderson 1971, Kent & Coker 1992), which operates on ranking dissimilarities between samples (Minchin 1987, Shupe 2005), was run choosing autopilot mode with slow and thorough speed and Sorenson similarity coefficient to search for the solution of best fit. The ordination was rerun with the recommended dimensionality. Among-group differences as well as the homogeneity of TWINSPAN classes were also examined by another non-metric test (MRPP) using the Sorenson similarity coefficient (PC-ORD, McCune & Mefford 1999, Van Sickle 1997).
Principal component analysis (PCA) (Orloci 1966, Kent & Coker 1992) and the canonical correspondence analysis (CCA) (ter Braak 1986, 1987) were used to determine the effects of diversity, species, and environmental variables (very coarse gravel %, percent very coarse gravel, bare ground % percent bare ground, altitude, species diversity, species richness, evenness, dominance, and species cover % percent species cover) on the separation of vegetation types as well as to find the relationships among the above mentioned variables. For PCA, a correlation matrix was used and non-trivial components were selected by comparing each principal component eigenvalue to a broken-stick eigenvalue (Jackson 1993). Useful or non-trivial components are those components with eigenvalues that exceed the eigenvalue of their broken-stick counterpart (Legendre & Legendre 1998). All the default settings were selected for CCA, and a Monte Carlo test (1,000 permutations) was done to test the canonical axes eigenvalues and the significance of the species-environment correlations. Pearson’s inter-set correlations from the CCAs were used to assess the significance of the tested variables. The correlations between variables and their effects on distribution of samples were also examined by using the CCA biplot procedure, which is carried out by superimposing radiating lines on the ordination space with their length proportional to the rate of variation (McCune & Mefford, 1999).
Results
Floristic summary. In total, 63 species representing 49 genera and 15 families were identified and recorded in the study area (see the Appendix 1 for details). Only 24 species occurred in vegetation samples. The recorded species mostly belonged to the Asteraceae (22.2 %), Chenopodiaceae (19.0 %), Poaceae (15.9 %), Brassicaceae (14.3 %), and Boraginaceae (9.5 %) families. Nine families were represented by a single species: Apiaceae, Caryophylaceae, Cyperaceae, Dipsacaceae, Geraniaceae, Papaveraceae, Polygonaceae, Tamaricaceae, and Zygophylaceae. Chronological analysis identified 57.14 % uniregional Irano-Turanian floristic elements. Therophytes (60.67 %) were the dominant life form, followed by hemicryptophytes (15.87 %), chamaephytes (9.52 %), and phanerophytes (7.94 %).
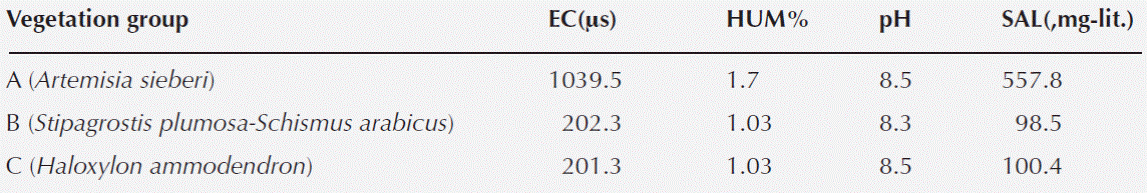
Soil analysis results. Quadrat values for soil moisture, EC, salinity, and pH were very similar among TWINSPAN groups. Interpreting the results as a mean value for each TWINSPAN group showed that the soil samples acidity reaction was strongly alkaline and nearly identical among samples. The results also indicated very low levels of soil moisture in all samples, confirming excessive soil aridity throughout the region. Soil electrical conductivity and soil salinity were nearly similar in soil samples of groups B and C, but showed remarkable variation in soils of group A (Table 1).
Table 1 Average values of the physico-chemical properties in top soil layer (0-15 cm) in 3 TWINSPAN communities
Multivariate analysis. Classification.-The TWINSPAN classification results of the 52 vegetation samples are represented in figure 2. According to the dendrogram and the eigenvalues of each division, the vegetation is divided into 3 major groups at level 2 of the TWINSPAN divisions. The groups were named after their dominant species.

Group A: Artemisia sieberi community. Twenty samples belonging to this community were found in the center plain, which is covered largely with very coarse gravel. The vegetation cover in this community is mainly composed of Artemisia sieberi, and the ground surface lacks almost any other vegetation cover. The soil of this community is more saline than for the other two. Companion species are Erodium cicutarium (L.) L’Hér., Halocharis sp., Schumannia karelinii, Scabiosa olivieri, Erysimum repandum L., Schismus arabicus Nees, and Astragalus tribuloides Delile. The average number of species per sample is 2.2, the Shannon-Wiener diversity index is 0.3, and the effective number of species, exp (0.3), is 1.35.
Group B: Stipagrostis plumosa - Schismus arabicus community. Twenty-seven samples belonging to this community were observed in the northern and eastern part of the region on the lower sand dunes. The soil in this community mainly lacked very coarse gravel particles and the soil reaction was alkaline. The community was sparsely covered with S. plumosa, S. arabicus, and very sparse and open woody Haloxylon ammodendron (C.A.Mey) Bunge ex Fenzl, not present in the samples. The companion species were Cornulaca aucheri Moq., Bromus tectorum L., Erodium cicutarium, Stipagrostis pennata (Trin.) De Winter, Boissiera squarrosa (Sol.) Nevski, Eremopyrum bonaepartis (Spreng.) Nevski, Londesia eriantha Fisch. & C.A. Mey, Heliotropium transoxanum Bunge, Salsola tomentosa (Moq.) Spach., and Cutandia memphitica (Spreng.) Benth. The average number of species per sample is 3.7, the Shannon-Wiener diversity index is 0.7, and the effective number of species, exp (0.7), is 2.01.
Group C: Haloxylon ammodendron community. The five samples representing this community were observed in the southern part of the region on higher sand dunes. It is an almost completely open woodland of H. ammodendron. Its only companion species is Schismus arabicus, with very little cover area. Very coarse gravel particles are rare., and soil reaction is similarly alkaline. The average number of species per sample is 1.2, the Shannon-Wiener diversity index is 0.2, and the effective number of species, exp (0.2), is 1.22.
Three relative cover clusters were recovered at a similarity level of 60 % (Figure 3). The clusters, referred to here as a, b, and c, were nearly identical with TWINSPAN groups A, B, and C, respectively. The only difference at this level of similarity was that three TWINSPAN group A samples (41, 46, and 51) were classified in cluster b (Figure 3).

Figure 3 Cluster dendrogram using Ward’s linking method and Euclidian distance measure with TWINSPAN groups superimposed
The MRPP procedure for testing whether there is no difference among TWINSPAN groups vs. there is significant variation among them, revealed that the average distance of each group was less than the overall mean distance, except for that of group B (Observed delta = 0.28, Expected delta= 0.47, MRPP test statistic: T = - 27. 29, P = 0.0000).
Multivariate analysis. Ordination.- In order to compare the classification and ordination results, the TWINSPAN groups were superimposed on to the DCA diagram (Figure 4) and separated along the first axis (eigenvalue = 0.957). The length of the gradient along the first axis was 5. 5 SD units and thus indicated a complete turnover in species composition. The diagram also shows that the three group A samples (41, 51, and 46) along with two group B samples (47 and 60) are in a transitional state between the two groups. The second axis (eigenvalue = 0.249), with a gradient length of 1.75, was not important.

The NMDS ordination diagram with 3-dimensional solutions and a stress value of 9.025 confirmed the separation of TWINSPAN groups along the axes of ordination (Figure 5). NMDS axis 1 separated the samples of group C from the samples of groups A and B. The samples of groups A and B were also separated along the NMDS axis 2. The diagram also shows that the group B samples are widely distributed along axis 2 and 3 of the NMDS ordination.

Figure 5 NMDS dimensions 1 and 2 of 3-dimensional ordination for 52 vegetation samples in the Badrud north region
Multivariate analysis. Vegetation-Environmental relationships.-The results of the PCA ordination for determining the effects of the measured variables on the separation of vegetation communities are presented in table 2 and figure 6. Comparing each axis eigenvalue with its broken-stick counterpart confirmed the first axis to be the only useful non-trivial component (4.633 vs. 2.718, respectively); it accounted for 57.91 % of the total variance. The correlation between variables and the first principal component confirms that this axis is negatively correlated with very coarse gravel %, and is positively correlated with ground bareness, species richness, evenness, diversity, and relative concentration of dominance. The ordination samples belonging to the Artemisia sieberi community (mostly in the negative part of the axis) are correlated with a high percentage of coarse gravel, and the Stipagrostis plumosa - Schismus arabicus community, mainly in the positive part of the axis, is correlated with ground bareness.


For CCA (Figure 7), the first canonical axis eigenvalue was 0.552, and the Monte Carlo test for evaluating it was highly significant (P = 0.007). The first axis correlation with species-environmental variables was 0.83 and the Monte Carlo test was significant (0.04). The second axis eigenvalue was 0.371 and the Monte Carlo test was highly significant (P = 0.002). Species-environmental correlation with the second axis was 0.68 and the Monte Carlo significance level was 0.07. The relations between environmental factors and the CCA axis indicate that the first canonical axis is strongly and positively correlated with ground bareness, richness, evenness, diversity, and relative concentration of dominance, while negatively correlated with altitude. The second axis has strong negative correlation with bare ground, and species cover, and is positively correlated with very coarse gravel % (r = 0.726).

Figure 7 CCA biplot diagram showing the diversity and environmental variables as radiating lines and the TWINSPAN groups superimposed
The correlation between environmental variables and diversity variables (table 3) shows that very coarse gravel cover has a strong negative effect on diversity (r = 0.591 and r = 0.590) and the number of species (species richness). The results also confirm that very coarse gravel has a negative effect on plant cover rate. Bare ground has strong positive effects on diversity variables.

Discussion
The floristic analysis of the study area recorded many drought resistant, xerophytic, halophytic, xerohalophytic and xeropsammophytic species, such as Artemisia sieberi, Salsola nitraria, Anabasis setifera Moq., and Cornulaca aucheri, reflective of an arid, salty, and sandy ecosystem. The life form spectra of the study area indicated the dominance of a typical desert life form. The dominance of annuals due to unfavorable conditions of little rainfall and excessive drought is in accordance with many findings in other deserts in Iran and other parts of the world (Asri 2003, Zarei et al. 2008, El-Ghanim et al. 2010, El-Ghani & Amer 2003, El-Sheikh et al. 2010). Communities with a single dominant species such as A. sieberi or Haloxylon ammodendron, which were recognized by multivariate techniques, are also reported in other deserts of Iran such as the Poshtkouh rangelands of the Yazd province (Jafari et al. 2004), which is dominated by A. sieberi. Wadi El-Sheikh of south Sinai and the gorges of Gebel Uweinat (Egypt) are also reported to have monotypic communities dominated by Artemisia judaica and Cornulaca monacantha, respectively (El-Ghareeb & Shabana 1990, Boulos 1982).
The other monotypic plant community of the region, Haloxylon ammodendron, is a component of the halophytic shrubby semi-woody communities on salty and dry soils, which is a characteristic of large parts of Iranian halophytic communities in the Irano-Turanian region and often occurs with Salsola tomentosa (Akhani 2004). As Salsola tomentosa was observed and recorded in the study area, its absence in such a community may be due to its very low relative cover percentage or because of the sampling method.
TWINSPAN eigenvalues for each division were greater than 0.7, confirming the strength of classification in level 2. Comparing the two classification methods showed the results to be slightly different; three TWINSPAN group A samples (41, 46, and 51) were classified in the Ward’s cluster b. Evaluating the results with the DCA and NMDS ordination techniques showed the three top samples to be more related to group A than B, suggesting that TWINSPAN groups in first levels are more interpretable than Ward’s method in final clusters. Considering the nearly identical and high relative cover percentage of the dominant species in these samples as well as two group B samples (60 and 47), which are classified in one cluster, indicates that primary clusters in Ward’s method were more interpretable than TWINSPAN final groups.
The ordination results also displayed transitional states in the distribution of samples along ordination axes. The MRPP results confirmed not only the separation of TWINSPAN groups but also the heterogeneity of group B, which is in accordance with the broad distribution of its samples along DCA axis 1 and NMDS axes 2 and 3. This can suggest a distinct group with Artemisia sieberi and Schismus arabicus as the dominant species.
The CCA eigenvalues lower than DCA’s and some samples which are out of their respective TWINSPAN groups in PCA and CCA ordination space indicated that some important explanatory variables were not included in the analysis or some of the variations in the data set were not explained by environmental variables (Franklin & Merlin, 1992; McDonald et al. 1996). There is thus no significant difference between groups in relation to these factors. As the correlations emphasize, these variables have greater effects on the number of species and diversity variables than on the amount of vegetation cover, and the relative cover has a stronger relationship with some variables that have not been measured in this research. Similar findings that soil texture have a greater influence on the number and nature of plants vs. the amount of cover have been reported by Shupe (2005) in southwest Arizona.
The soil analysis also showed the Artemisia sieberi community is remarkably related to higher levels of electrical conductivity (EC). The findings are not in accordance with the findings of Jafari et al. (2004).
Comparing the EC and pH values measured in this study with the values reported by the Iran Research Institute of Forests and Rangelands in 1997 suggests a succession gradient in soil properties. The soil acidity has changed from 7.7 (neutral) to alkaline (8.9) and the EC levels in the top soil layer (0-15 cm) from 2.66 mS to 0.47 ?S. These can be signs of succession towards the process of desertification. These changes may be due to the physical processes of wind erosion occurring quite often in sand dunes and also due to overgrazing pressure, which in turn increases soil erosion (Shao et al. 1988, Dong et al. 1993). As the previous report was incomplete from the viewpoint of species composition, determining the effects of these gradients on the floristic structure of the region is not fully possible. Ephedra strobilacea, related with neutral pH, (Jafari et al. 2004) was supposed to be seen in the region according to the previous report. Its absence, together with the absence of many halophilous species that were expected to be seen in the region, can be due to the above-mentioned processes. The soil texture coarseness in this area and its negative effects on species diversity, together with high rates of soil erosion and heavy grazing with their increasing effects on soil coarseness leading to severe and fast moving process of desertification, confirms the necessity to draw more attention to the area, including the complete exclusion of livestock grazing in order to prevent further degradation of vegetation and to preserve potential sources of biodiversity confined to this region. The results also could help guide better management for the conservation of species diversity in other arid and semi-arid ecosystems.


