











 text new page (beta)
text new page (beta) English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkPeople in tropical regions frequently depend on species-rich agroforestry systems such as homegardens for sustenance (Michon et al., 1986; Gajaseni and Gajaseni, 1999; Nair and Kumar, 2006). It is thought that homegarden management is sustainable, maintaining a high diversity of local and introduced taxa (Pulido et al., 2008), and enhancing long-term productivity (Michon et al., 1986; Gajaseni and Gajaseni, 1999). However, some traditional practices are aimed at a single species, as is the case of leaf harvesting and trimming, seed sowing, or fencing out saplings to protect them from livestock. Other practices are applied at the homegarden scale, such as flooding irrigation, or the manipulation of the canopy density related with the species that the managers chose to grow. Practices at both levels may unintentionally or indirectly be detrimental to some valuable species, but they may still be carried out if the stakeholder finds out that the overall benefits overcome the drawbacks. Thus, traditional management of agroforestry systems involves several compromises (Nair, 2001; Kehlenbeck and Maass, 2006; Torquebiau and Penot, 2006).
The management of Sabal Adans. palms in homegardens first appeared during the Colonial period (Caballero, 1994), and is currently one of the most common agricultural systems in Yucatan Peninsula, Mexico. Two species of Sabal palms (Arecaceae) are promoted in homegardens, Sabal yapa C. Wright ex Becc. and Sabal mexicana Mart. The same management practices aiming at caring for the palms were observed for both species. Seedlings are protected from domestic animals and occasionally seeds are planted. Both species of palms are equally harvested by cutting leaves once or twice a year, leaving one or two leaves on the palm, together with the developing apical meristem or “cogollo” (Gama-Rios, 2001). Only the intermediate-sized palms (juveniles) and the short adults are used for leaf harvesting. Palm leaves are constantly harvested even when these are not immediately needed, because, according to the Maya, this promotes the production of new leaves.
From an ecological perspective, sustainable use of a natural resource occurs when its utilization does not compromise its natural replenishment, the resource therefore being capable of renewing itself indefinitely (Mangel et al., 1993; Olmsted and Álvarez-Buylla, 1995; Joyal, 1996). However, sustainability involves processes ranging from individual organisms to whole ecosystems, as well as biological, economic and social factors acting at different spatial scales (Padoch and Sears, 2005; Spangenberg, 2011). In order to assess how management of the homegarden interacts with specific practices aimed at palm species, and to find out if such management promotes sustainability of palm populations, we conducted a study in two homegardens in Maxcanú, Yucatán, Mexico where both species of Sabal are management to obtain leaves. The specific questions addressed in this study were: How does management at the level of the whole homegarden affects the population dynamics of Sabal spp.? How do management practices at the levels of the whole homegarden and of individual species interact for achieving sustainable use?
Methods
Study site and species description. Sabal is a genus of solitary palms reaching over 10 m in height. They usually have about 10-20 fan-shaped leaves, although smaller individuals may rarely have up to 10 leaves. Flowering occurs in the spring in large spikes producing thousands of fruits in the summer. Most of the germination takes place during the rainy season. Seed banks are rarely formed in palms because seeds rapidly rot or are eaten by granivores (Vázquez-Yanes and Orozco-Segovia, 1993; Svenning, 2001).
The economy of the village of Maxcanú, located in the SW of the state of Yucatan, Mexico (Figure 1), is based on self-subsistence and commercial agriculture, as well as in cultivation of fruit trees. The mean annual temperature is 26.4 °C and the mean annual precipitation amounts to 1,074 mm. Rainfall is largely concentrated in the summer, with a harsh drought occurring from October to May (SMN n/d). We worked in two homegardens designated as H1 and H2. Homegarden H1 had a surface area of 0.61 ha, was frequently irrigated by flooding to promote the growth of fruit trees, and, due to a high density of trees, it was shaded throughout the year. H2 had a surface area of 0.25 ha, irrigation was absent thus entirely depending on rainfall. Tree density was lower than in H1 and many trees lost most of their leaves during the dry season, so that solar irradiation was higher than in H1.
Data collection. Along randomly located transects in H1 and H2 we selected 143 and 152 individuals of Sabal yapa, and 144 and 188 individuals of S. mexicana, respectively. Due to the low number of tall plants, all the adults in the homegardens were marked. Plants without evident stems were classified according to their leaf development as: seedlings (SD), recently germinated palm seedlings younger than 1 year having lanceolate leaves; saplings, which were divided in three categories according to leaf-morphology, as saplings with bifid leaves (SA1), saplings with costapalmate leaves with segments whose extremes are incompletely divided (SA2), and saplings with completely divided costapalmate leaves (SA3). Palms with evident stem were classified according to their height in two categories, each one subdivided by tallness: juveniles of 1-50 cm (J1), 51-100 cm (J2), 101-200 cm (J3), and 201-300 cm (J4) in height; and reproductive adults 301-400 cm (A1), 401-650 cm (A2), and more than 650 cm (A3) in height.
In order to estimate the number of fruits produced, once a year we counted the number of spikes produced, and measured the maximum and perpendicular diameters of their basal stem (Martínez-Ballesté et al., 2005). All the new seedlings found in transects were counted and incorporated to the sample. All plants were followed yearly during the years 1998 to 2000, and their developmental stage and survival were recorded.
Theoretical Models of the Prospective and Retrospective Analyses. Sustainable use of palm populations can be assessed through demographic studies, which provide data on the availability of the resource in the long term by means of the finite population growth rate (λ) (Grenand and Grenand, 1996; Caswell, 2001). Management is sustainable when the population is either in equilibrium (i.e., λ = 1), or growing (λ > 1) (Olmsted and Álvarez-Buylla, 1995; Joyal, 1996; Ticktin, 2002). Confidence values for each of the λ values obtained were calculated with the “Series Approximation” method (Caswell, 2001).
The size structures observed in populations of both species were evaluated using χ2 analysis and Haberman-adjusted residuals test. Also, prospective elasticity and retrospective -or Life Table Response Experiment (LTRE)- analyses were obtained. The elasticity analysis was used to evaluate the relative effect on λ of the small and hypothetical changes on vital rates (survival, growth, and fecundity of individuals in a population) as a result of environmental or management changes. This model assumes that the observed conditions will remain constant through time. Vital rates with the highest elasticity values showed the stages of the palm´s life cycle having the highest effect on λ in the eventuality of changes occurring (Caswell, 2001). The retrospective analysis (LTRE) was estimated in order to identify the stages of the palm life cycle (vital rates) that most contributed to the variation in population growth rates (λ). The variations observed are due to the observed differences between species (Sabal yapa and S. mexicana), sites (microenvironmental differences and management practices between homegardens), year (management differences between years), and the combined effects of these variables. A positive LTRE value corresponds to palm stages that contributed to increase the value of λ, while a negative value indicates that the changes seen in this stage of the life cycle favored a decrease in population growth rate (λ). Null values of LTRE indicate vital rates that showed no variation, therefore having no effect on λ (Caswell, 1989; Horvitz et al., 1997). If management negatively affects the vital rates with high elasticity values, and if the annual observed variation (LTRE) in those vital rates shows high and negative values, then the population growth rates would be negative (λ < 1) (Zuidema et al., 2007). In such case, we would be facing an unfavorable situation for sustainable resource management. By combining these analyses, we can understand the effect on palm population dynamics of management practices in the homegardens, as well as the effect on population growth rates (λ) of changes in management decisions.
Results
Size structure. Seedlings constituted the most abundant size category for both species and in both homegardens, but there were significant differences in other categories (χ2 = 131.09, P < 0.0001; Figure 2). Differences in size structures were mainly caused by variation in the proportions of saplings and juveniles. In Sabal mexicana the proportion of juvenile individuals (at the adequate harvesting size) was greater compared to the proportion of individuals in size categories that are not commonly harvested (saplings and adults). In H1, S. yapa saplings predominated, while in H2 the proportion of larger juveniles of S. mexicana (J2 and J3) was significantly higher (Figure 2).
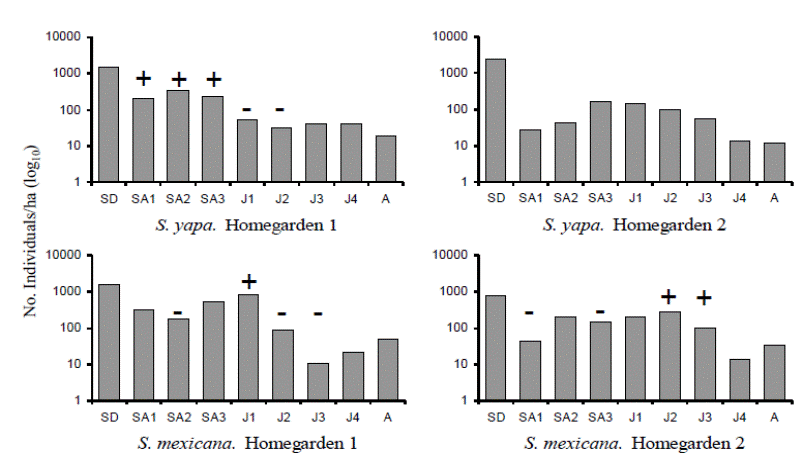
Figure 2 Density and size structure of palm populations. The +/- symbols show size categories with significantly larger or smaller proportion of individuals regarding the other palm populations studied (Haberman-adjusted residual test, (χ2 = 131.09, P < 0.0001).
Population growth rates. The average population growth rates obtained for the populations of each species during the three years (λ; interval of confidence of 95 %) were: 1.007 (0.924-1.090) and 1.092 (0.997-1.187) for Sabal yapa in H1 and H2, respectively; 1.048 (0.980-1.116) for S. mexicana in H1 and 0.983 (0.929-1.037) in H2. The estimated confidence intervals indicated that population growth rates did not differ from 1, or between species populations.
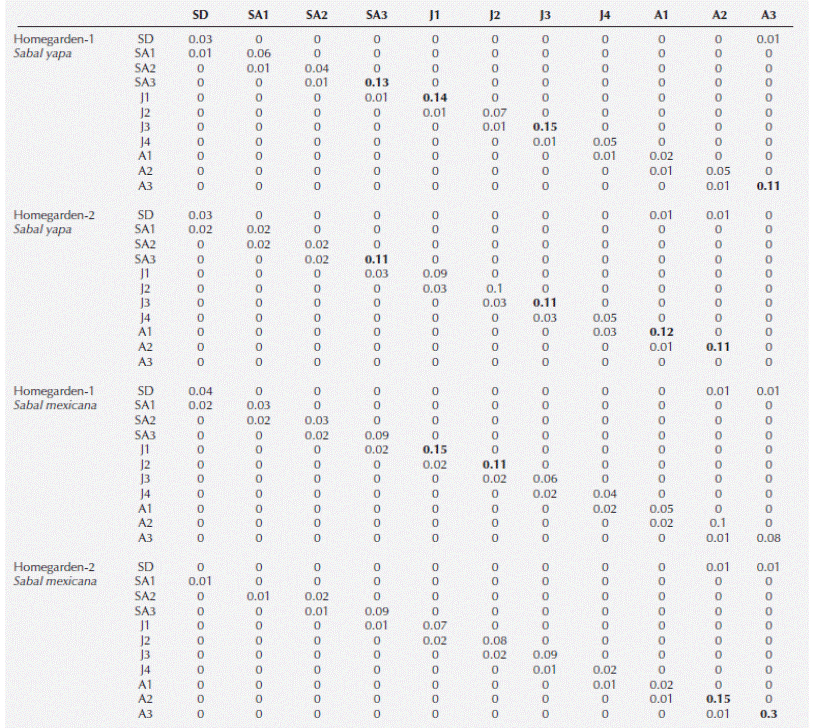
Prospective elasticity analysis. The elasticity analysis shows that survival of individuals remaining in the same size category was expressed by the vital rates with the highest elasticity values in both species, followed by the growth or transition to the next category and, finally, by the fecundity of the adult palms. The highest elasticity values were found in the survival of adult categories, saplings (SA3) and juveniles (Table 1).
Table 1 Elasticity matrices for Sabal yapa and Sabal mexicana in two traditional homegardens. High values are shown in boldface. Palm size categories: seedlings (SD); saplings lacking an above-ground stem with bifid leaves (SA1), with incompletely divided leaves (SA2), and with completely-divided leaves (SA3); juveniles with stems 1-50 cm (J1), 51-100 cm (J2), 101-200 cm (J3), and 201-300 cm (J4) in height; and reproductive adult individuals with stems 301-400 cm (A1), 401-650 cm (A2), and 651 cm or above (A3) in height.
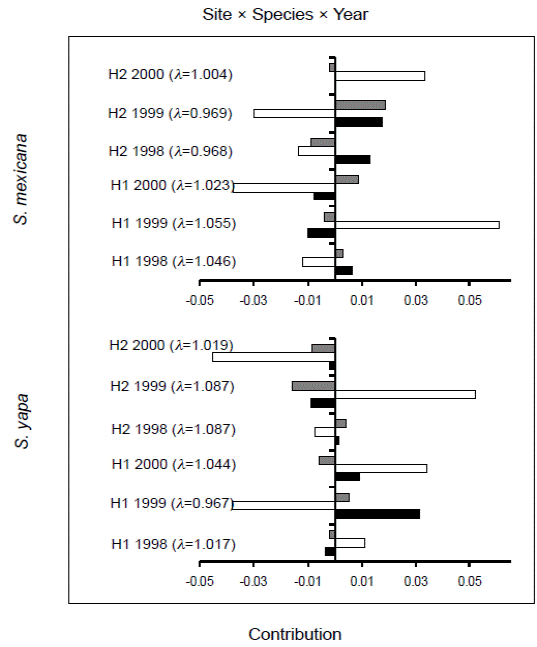
Retrospective analysis. The interaction “site × species” of retrospective analysis (Figure 3) showed that the populations of Sabal yapa in H2 and of S. mexicana in H1 had positive contributions from all of their population parameters -survival, growth and fecundity-, and their population growth rates (λ) were higher than those obtained for S. mexicana in H2 and for S. yapa in H1. In the two latter populations the contributions were negative for the three vital rates. Nevertheless, when the effect of the factor “year” is added in a triple interaction (year × site × species) (Figure 4), the values of λ in S. yapa populations in H1 and of S. mexicana in H2 decreased bellow one in 1999, while in that same year, S. mexicana in H1 and S. yapa in H2 showed an increase in their population growth rates (λ). In 1998, the populations of S. mexicana and S. yapa in H2, and of S. mexicana in H1 showed negative contributions due to the growth of individuals, while in the year 2000 the vital rate growth increased the values of λ of the populations of S. mexicana from H2 and of S. yapa in H1.
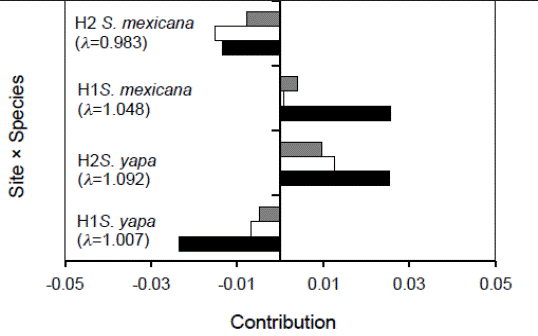
Figure 3 Contribution of vital rates -survival (solid), growth (clear), and fecundity (shaded)- to the observed variation in population growth rates (λ) due to the effect of the site × species interaction.
Discussion
The effect of homegarden management on Sabal populations. The main purpose of homegardens as managed agroforestry systems is to insure family livelihood. If a given resource decreases, or if it disappears from other sources, the family will try to intensify management practices in the homegardens (Arnold and Dewees, 1995; Kremen et al., 1998; Nair, 2001). These practices could affected the vital rates and the size structure of natural resources populations, for example, it has been found that populations of oak growing in the agroforestry systems known in Spain as “dehesas” contain individuals with thicker and taller stems than individuals from unmanaged forests (Pulido et al., 2001). Another study made in the Los Tuxtlas Biosphere Reserve showed differences in the size structure of palms and cycads growing in unmanaged sites relative to the size structure observed in managed tropical forests with presence of Chamaedorea plantations. The management of such plantations promotes the pruning of understory species to increase light availability and to reduce competition, which favors an increase in the proportion of smaller palms and cycads relative to the same in unmanaged forest (Trauernicht and Ticktin, 2005).
Typically, the size structure of unmanaged populations of wild perennial plants shows an inverted-j, in which seedlings and adults are predominant (Piñero et al., 1984; Ramp, 1989; Ratsirarson et al., 1996). Also, Pulido and Caballero (2006) found that populations of Sabal yapa growing in mature tropical forest or secondary vegetation where management is unintensive had size structures with significantly more adults and seedlings.
The Sabal populations that we studied showed a size structure with a significantly higher proportion of saplings and juvenile palms. Some management practices in the homegardens could be related to such an increase in these palm size categories. Seedling care and the occasional planting could increase the density and survival probabilities of individuals in the earlier categories. In addition, the shady environment of homegardens -particularly in H1 in which many tree species are promoted- decreases palm growth (Figure 3 see H1 S. yapa), thus increasing the proportion of saplings and lower height juveniles (J1) (see Figure. 2) remaining at an adequate harvesting size for a longer period of time. Other practices in the homegarden could increase survival probabilities (Figure 3 see H1 S. mexicana and H2 S. yapa), such as weeding (that eliminates competitors) and, indirectly, watering of fruit trees, which increases the humidity of homegardens.
Although, the three-year average λ value of each population was equal to 1, indicating that Sabal yapa and S. mexicana populations are capable of long-term persistence. The variation analysis (LTRE) of population parameters observed in the studied populations of palms showed that homegarden management change the population dynamics, affecting in several ways the resource´s sustainability.
The homegarden is an inhabited agroforestry system. Unlike other agroforestry systems homegardens are spaces inhabited by human families, therefore containing all sorts of useful resources while simultaneously providing people with a relaxing and restful environment, and are the space in which the family lives and develops (Lok, 1998; Bhatti and Church, 2004; Abdoellah et al., 2006). While along the three years of this study we did not observe the death of adult or juvenile palms due to traditional harvesting practices, we did observe that other activities in the homegarden that were not directed to the management of Sabal individuals had negative effects on juvenile and adult palms. Whereas these size categories palms have high elasticity values, their proportional contribution to the value of λ is important, such that the increase of mortality in these size categories resulted in a decreased value of λ (Zuidema and Zagt, 2000).
In the year 2000, a member of the family harvested all the leaves included the cogollo of some saplings and juvenile individuals in H2. Although this is an uncommon harvest practice, the event increased the annual negative variation (LTRE) of the three population parameters of Sabal yapa, thus decreasing the annual growth (λ) of the population from 8 % (λ = 1.087) in 1998 and 1999, to 1 % in 2000 (λ = 1.019) (Figure 4, see H2 S. yapa 2000). Likewise, in the year 1999 many adults of Sabal palms were cut down in both homegardens in order to clear land for building new houses for newly married sons or daughters. This felling event reduced the survival rates of adult palms, thus affecting the growth rates (λ) of Sabal populations. In both species the values of λ for the year 1999 were negative (< 1) (Figure 4: H1, λ = 0.967 and H2, λ = 0.969).
The domestic events occurring during the three years in which this study was conducted showed us how the population dynamics of species managed in homegardens can be altered for reasons beyond the interest of promoting a natural resource, but must be taken into consideration when studying the sustainability of species management in this type of agroforestry systems.
The microenvironmental conditions of the homegarden. Although annual climatic variation is an important variable in the analysis of population dynamics of natural resources, in the case of the studied homegardens the yearly differences in management and the microenvironmental conditions generated by the different management practices applied were also relevant factors affecting the sustainability of the resource. The meta-analysis of 29 studies of homegardens made in Latin American (Pulido et al., 2008) showed that species richness was more related to microenvironmental conditions created by management of the homegardens, than to physical variables such as altitude, latitude, and precipitation.
By analyzing the effects of management on the population dynamics of the two species of Sabal (Figure 3) we found that in shadier environments with more available water (from irrigation), such as in H1, the population growth rate (λ = 1.048) of S. mexicana was higher than that observed for the S. yapa population (λ = 1.007), the three population parameters of the population of S. mexicana being positive, while those of S. yapa were negative. The opposite was true in H2 --a homegarden with less trees-- in which S. yapa was benefited by the higher availability of light, thus its three demographic parameters having positive values (λ = 1.092), while the population growth rate of S. mexicana (λ = 0.983) was below 1 due to the negative contribution of survival, growth and fecundity of its individuals.
From an ecological point of view, both species are capable of colonizing human-altered sites and are adapted for survival in the absence of irrigation (Zona, 1990). Nevertheless, under the microenvironmental conditions of each homegarden the ecological performance of each species was different, appearing to be related to the environmental requirements of the species. Being Sabal yapa a species native to the Yucatán Peninsula it is adapted to seasonally dry tropical forests, karstic soils, and a sub-humid warm climate with a yearly precipitation of 600 to 2,000 mm. In contrast, S. mexicana naturally grows in tropical forests with a higher total annual precipitation that can be of up to 4,000 mm (Zona, 1990; Quero, 1992; Caballero, 1994). Although in both homegardens both palm species grow under the same environmental conditions (same average annual precipitation and temperature), irrigation in H1 increased water availability and favored the growth of individuals of S. mexicana, which naturally prefers more humid locations, while S. yapa grew best in H2 having a lower availability of water.
Conclusion. Homegardens are complex and dynamic systems with continuously changing management and microenvironmental conditions. The decisions taken by Maya peasants within these agroforestry systems can vary from one homegarden to another, which generates differences in the sustainability balance of the involved populations. Although the populations in both homegardens showed a stable numerical behavior reflecting the sustainability of the resource, the way in which this happened varied according to the specific characteristics of each site. The factors that affected the population dynamics in both species of Sabal were related to differences in management, domestic events, and performance of each species under the specific microenvironment of each one of the studied homegardens.
Our study shows that the management of natural resources in homegardens occurs as an interaction of events that are not always focused on the sustainability of specific resources, but rather on the integrated management of the whole agroecosystem within the context of the activities of the inhabiting families.

