










Services on Demand
Journal
Article
 text in
text in  English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista Chapingo serie ciencias forestales y del ambiente
On-line version ISSN 2007-4018Print version ISSN 2007-3828
Rev. Chapingo ser. cienc. for. ambient vol.24 n.1 Chapingo Jan./Apr. 2018
https://doi.org/10.5154/r.rchscfa.2017.09.055
Artículo cientifico
Reservorios y dinámica de nutrientes en dos rodales bajo aprovechamiento de Pinus montezumae Lamb. en Tlaxcala, México
1Colegio de Postgraduados, Campus Montecillo. km 36.5 carretera México-Texcoco. C. P. 56230. Texcoco, Estado de México, México.
2Universidad Autónoma de Chihuahua, Campus Delicias, Facultad de Ciencias Agrícolas y Forestales. km 2.5 carretera Delicias-Rosales. C. P. 33000. Cd. Delicias, Chihuahua, México.
3Universidad Autónoma Chapingo. km 38.5 carretera México-Texcoco. C. P. 56230. Chapingo, Texcoco, Estado de México, México.
Introducción:
El estudio del ciclo de nutrientes es importante para proponer métodos de aprovechamiento sostenible en los bosques.
Objetivos:
Evaluar los reservorios de N, P, K, Ca y Mg en árboles, mantillo y suelo mineral de dos rodales de Pinus montezumae Lamb. bajo aprovechamiento; estimar la tasa de descomposición de mantillo; y cuantificar la cantidad anual de nutrientes demandada.
Materiales y métodos:
El estudio se realizó en Nanacamilpa, Tlaxcala. La biomasa y masa de nutrientes en el fuste, ramas y hojas se estimaron con un muestreo destructivo de 10 árboles.
Resultados y discusión:
Los datos dasométricos, reservorios y dinámica de nutrientes fueron similares en ambos rodales (t, P > 0.05). En el suelo mineral, los contenidos de N, P, K, Ca y Mg fueron 5 419, 68, 1 245, 9 163 y 2 029 kg·ha-1, respectivamente; y en la biomasa aérea fueron 242, 12, 28, 167 y 118 kg·ha-1, respectivamente. La tasa de descomposición de hojarasca fue -0.335. El arbolado requiere, para su crecimiento, 73 a 81 kg·ha-1·año-1 de N, 5.2 a 5.3 kg·ha-1·año-1 de P y 9.4 a 10 kg·ha-1·año-1 de K.
Conclusión:
Los nutrientes del suelo podrían mantener el crecimiento del bosque por 256 años, pero el P podría ser un limitante si las prácticas sustentables de cosecha se modifican.
Palabras clave: Ciclo de nutrientes; cosecha forestal; manejo forestal; bosques de pino
Objectives:
To evaluate the reservoirs of N, P, K, Ca and Mg in trees, litter and mineral soil of two managed stands of Pinus montezumae Lamb.; estimate the decomposition rate of litter; and quantify the annual amount of nutrients required for tree growth.
Materials and methods:
The study was conducted in Nanacamilpa, Tlaxcala. The biomass and mass of nutrients in trunk, branches and leaves were estimated with a destructive sampling of 10 trees.
Results and discussion:
Dasometric data, reservoirs and nutrient dynamics were similar in both stands (t, P > 0.05). In mineral soil, the contents of N, P, K, Ca and Mg were 5 419, 68, 1 245, 9 163 and 2 029 kg·ha-1, respectively; and in aerial biomass were 242, 12, 28, 167 and 118 kg·ha-1, respectively. The decomposition rate of leaf-litter was -0.335. Trees require 73 to 81 kg·ha-1·year-1 of N, 5.2 to 5.3 kg·ha-1·year-1 of P and 9.4 to 10 kg·ha-1·year-1 of K to grow.
Conclusion:
Soil nutrients could maintain forest growth for 256 years, but P could be a limiting factor if sustainable harvest practices change.
Keywords: Nutrient cycle; forest harvesting; forest management; pine forests
Introducción
El reservorio de nutrientes y su movimiento en los bosques es un conocimiento básico que permite saber si un sistema de aprovechamiento es sostenible (Johnson, Sogn, & Kvindesland, 2000). De acuerdo con la literatura, la salida de nutrientes a causa de la cosecha forestal no amenaza la sustentabilidad, debido que algunos de estos son liberados por el material parental del suelo (Binkley & Fisher, 2013; Sollins et al., 1980), regresan al suelo a través de la caída de hojarasca y tienen tasas altas de reabsorción en la masa foliar; además, las especies forestales demandan pocos de ellos (Imbert, Blanco, & Castillo, 2004; Machado, Sampaio, Ferraz, Camara, & Pereira, 2016). Sin embargo, no siempre debe asumirse que la cosecha forestal no impacta el nivel productivo de los terrenos, más bien debe ser evaluado, ya que la falta de información en el tema deja más incertidumbre que certeza, especialmente en el caso de México donde la información es escasa.
En el dosel, los nutrientes se almacenan en el fuste, ramas y follaje. La duración de follaje antes de la caída y la reabsorción de los nutrientes determinan el tiempo promedio que estos residen en la biomasa aérea antes de regresar al suelo. En este sentido, Rodríguez-Sanfuentes y Álvarez-Muñoz (2010) indican que la rapidez de los ciclos de nutrientes influye en la productividad del bosque. Aunque no siempre es posible estudiar todos los reservorios y transferencias, la medición de los flujos principales del suelo a la vegetación y viceversa es importante, ya que ayuda a comprender el funcionamiento de un ecosistema forestal (Imbert et al., 2004).
En los bosques maduros, el reservorio más grande de nutrientes está en el suelo (Schlesinger & Bernhardt, 2013); no obstante, la disponibilidad de algunos de estos depende de la velocidad de mineralización de la hojarasca más que de la magnitud del reservorio (Grigal, 2000; Landsberg & Gower, 1997). Cabe señalar que el aprovechamiento forestal inadecuado puede abatir los reservorios de nutrientes (Imbert et al., 2004; Landsberg & Gower, 1997; Maynard et al., 2014), debido a la remoción de biomasa y la vulnerabilidad del suelo a la erosión y compactación (Gomez, Powers, Singer, & Horwath, 2002; Grigal, 2000; Imbert et al., 2004); en consecuencia, la productividad también puede ser afectada (Goya, Pérez, Frangi, & Fernández, 2003; Grigal, 2000).
Los objetivos de este estudio fueron: evaluar los reservorios de nutrientes (N, P, K, Ca y Mg) en árboles, mantillo y suelo mineral (a una profundidad de 80 cm) en dos rodales de Pinus montezumae Lamb., aprovechados con el Método Mexicano de Ordenación de Bosques Irregulares; estimar la tasa de descomposición de mantillo; y cuantificar la cantidad anual de nutrientes que un rodal maduro de P. montezumae demanda.
Materiales y métodos
Área de estudio
El estudio se realizó en el predio San José Nanacamilpa (19° 29’ 0’’ N y 98° 32’ 0’’ O), municipio de Nanacamilpa, Tlaxcala. De acuerdo con la estación meteorológica de Nanacamilpa (29017), la temperatura media anual es 8 °C y la precipitación media anual es 744 mm (Eric III, 2013). Los dos rodales están conformados por masas puras de P. montezumae y el sotobosque se compone por Symphoricarpos microphyllus H. B. K., Baccharis conferta Kunth. y especies de los géneros Senecio, Salvia, Ceanothus y Muhlenbergia. La pendiente, exposición y altitud de los rodales son: 4 %, Noreste y 2 600 m, respectivamente, en el rodal I; y 8 %, Suroeste y 2 900 m, respectivamente, en el rodal II. El bosque ha estado sujeto a manejo mediante el Método Mexicano de Ordenación para Bosques Irregulares (Hernández-Díaz, Corral-Rivas, Quiñones-Chávez, Bacon-Sobbe, & Vargas-Larreta, 2008). La extracción de volumen medio anual es de 5.9 m3·ha-1, que equivale aproximadamente a 3 % de la existencia real total.
Sitios de muestreo
En cada rodal se delimitaron tres sitios circulares de 0.1 ha; uno al centro y dos complementarios ubicados a 50 m. En los tres sitios de muestreo se cavó un perfil de suelo de 2 × 1 × 1 m de largo, ancho y profundidad, respectivamente, y se tomaron muestras por horizonte, para su análisis y estimación de la masa de nutrientes. El diámetro normal (cm) y altura total (m) de todos los árboles, de los tres sitios de cada rodal, se midieron con cinta diamétrica y pistola Haga, respectivamente.
Biomasa aérea
En los rodales estudiados, 10 árboles con diámetro normal entre 40 y 55 cm se derribaron para estimar la biomasa aérea. El fuste se seccionó en trozas de 2.5 m en la base del árbol y 1.25 m en la parte superior, posteriormente se cubicaron con la fórmula de Smalian (Romahn & Ramírez, 2010). La biomasa seca por unidad de volumen verde se determinó a partir de tres rodajas de 5 cm de grosor, tomadas a lo largo del fuste. De cada rodaja se tomó una submuestra y se secó en una estufa con aire de circulación forzada (Sheldon Fx14-S, USA) a 85 °C hasta obtener peso constante.
Las ramas y el follaje de cada árbol se separaron y se pesaron en verde, directamente en campo, con una báscula portátil de gancho con capacidad de 15 kg y precisión de 0.1 kg (marca Pasola). Por separado se tomaron tres submuestras de aproximadamente 1 kg de follaje y 2 kg de ramas, pesadas con una balanza de capacidad de 2 kg y precisión de 0.1 g (Ohaus Scout Pro, USA). Las submuestras se secaron en estufa a 70 °C para estimar el contenido de humedad y posteriormente transformar el peso de campo a peso seco. El peso seco de campo de los componentes vegetales se obtuvo con la fórmula siguiente:
donde
PSc |
peso seco de campo |
Phc |
peso húmedo de campo |
Fh |
fracción de humedad (peso del agua/peso seco). |
Con la biomasa total de cada árbol se estimó la proporción de biomasa total aérea asignada al fuste, las ramas y el follaje. En los tres sitios de cada rodal, el volumen se estimó con los diámetros normales y las alturas totales del árbol, empleando la ecuación para volumen total del árbol. La masa se determinó a través del volumen y densidad de la madera estimada.
Piso forestal
Tres muestras de hojarasca del piso forestal (horizonte orgánico superficial) se obtuvieron en cada rodal, empleando un marco circular de 30 cm de diámetro (0.071 m2). Un punto de muestreo se ubicó al centro del rodal y, aproximadamente a 25 m, se ubicaron otros dos a ambos lados en posición perpendicular a la pendiente. Cada muestra se pesó en húmedo en campo, posteriormente se seleccionó una submuestra y se secó por 72 h a 70 °C en estufa con aire de circulación forzada (Riossa HCF-4848D, México). Una vez conocidos los pesos húmedos y secos, se estimó la cantidad de biomasa existente en el piso forestal.
Suelo mineral
Las muestras se obtuvieron en cada horizonte de los perfiles de suelo hasta llegar al horizonte C, el cual se encontró a 115 y 95 cm en el rodal I y II, respectivamente. En los sitios complementarios solo se muestrearon los primeros tres horizontes, donde la mayor cantidad de nutrientes se concentra. Determinaciones preliminares de suelo, realizadas en el área, indicaron que la mayor variabilidad se encuentra en el suelo superficial. También se tomaron muestras para obtener la densidad aparente de suelo con un muestreador con martillo de deslizamiento (AMS Inc., Idaho, USA) con cilindros 98.17 cm3. Las muestras de suelo se secaron a 105 °C durante 12 h en estufa con aire de circulación forzada (Riossa HCF-4848D); la densidad aparente se determinó con la información de volumen y masa del suelo seco. Cada uno de los perfiles se analizaron por horizonte, de acuerdo con la Norma Oficial Mexicana NOM-021-RECNAT-2000 (SEMARNAT, 2002) para determinar: textura, pH, capacidad de intercambio catiónico, materia orgánica, N total, P extractable (método Bray 1) y cationes intercambiables (K, Ca y Mg) en acetato de amonio (pH 7). Considerando la profundidad de los horizontes y su densidad aparente, la cantidad de nutrientes se estimó a una misma profundidad de 80 cm para cada rodal. La mayoría de los reservorios de nutrientes en los bosques se encuentran a este nivel de profundidad (Binkley & Fisher, 2013).
Producción y descomposición de la hojarasca
La caída de hojarasca se colectó en periodos aproximados de un mes durante un año. Para ello, en cada rodal se colocaron tres trampas cuadradas de 1 m2 a nivel del suelo. Las muestras de hojarasca se secaron a 70 °C por tres días en estufa con aire de circulación forzada (Riossa HCF-4848D, México.). La masa neta de nutrientes que regresa al suelo se estimó a partir del peso seco de hojarasca y la concentración de sus nutrientes (N, P, K, Ca y Mg).
Por otra parte, en cada rodal se colocaron 25 bolsas de polipropileno negro de 17 × 25 cm con abertura de 2 mm2 (Bärlocher, 2005). Estas bolsas de descomposición contenían 20 g de hojarasca previamente secada. La muestra no incluyó ramas o fragmentos de madera. Las bolsas se colectaron en diferentes intervalos de tiempo (63, 126, 217, 252 y 336 días) de tal forma que se pudiera calcular la tasa de descomposición de acuerdo con el siguiente modelo exponencial (Schlesinger & Bernhardt, 2013):
donde,
Y |
masa de hojarasca remanente (g) |
C |
masa de hojarasca inicial (g) |
k |
tasa de descomposición (g·día-1) |
t |
tiempo (días) |
El muestreo de bolsas se planeó inicialmente cada 60 días, pero finalmente se hizo acorde con la disponibilidad de recursos para el estudio. Los cambios en la masa neta de hojarasca durante el año permitieron hacer una estimación de la velocidad de descomposición.
Reabsorción de nutrientes
La reabsorción de N, P y K del follaje se calculó con la diferencia de la concentración de nutrientes entre las hojas de 2014 completamente desarrolladas (agosto) y las hojas senescentes (noviembre). La muestra total fue de 20 árboles (10 por rodal). La masa de N, P y K reabsorbido se obtuvo a partir del porcentaje de reabsorción estimado. La reabsorción se obtuvo solo para los macronutrientes más móviles en la hoja, ya que los menos móviles, especialmente el Ca, no se pueden estimar con el método empleado en este estudio.
Salida de nutrientes debido al aprovechamiento forestal
Con la finalidad de tener un escenario conservador, que incluyera remoción adicional de arbolado por causas distintas al plan de aprovechamiento, se tomó como referencia 4 % de la existencia real como el volumen de madera extraída anualmente y su equivalente en masa de fustes y ramas, considerando la densidad de la madera. Este dato se sustentó con información del plan de manejo del área que estima un incremento corriente anual de 2 a 3 %, el cual es consistente con lo reportado por Zepeda y Acosta (2000) en bosques de P. montezumae. La salida de cada nutriente se estimó con las concentraciones de nutrientes en fustes y ramas. No se tomó en cuenta la masa del follaje puesto que permanece en el sitio y no forma parte de las salidas.
Requerimiento anual de nutrientes y reservas potenciales
De acuerdo con los datos del historial de manejo del bosque y la demanda de nutrientes de otros componentes como las raíces, se consideró razonable mantener 4 % como incremento corriente anual, tal como se hizo en la salida de nutrientes. Por lo anterior, esta referencia sirvió para estimar la ganancia neta anual de nutrientes de los componentes maderables asumiendo las proporciones del fuste, ramas y hojas del muestreo destructivo. La masa de N, P y K, requerida anualmente (kg·ha-1) para la formación de los componentes, se calculó como lo indican Schlesinger y Bernhardt (2013). De acuerdo con estos autores, la absorción es la suma de los nutrientes en biomasa más lo que regresa al piso forestal, y el requerimiento es la suma de la absorción más la reabsorción. Debido a que las cantidades de reabsorción de Ca y Mg no se pudieron estimar, los requerimientos de los mismos tampoco se obtuvieron.
Asumiendo que las salidas principales de nutrientes se originan por la cosecha forestal y que el capital de nutrientes del suelo es el estimado en cada perfil, el número de años que el suelo podría proveer nutrientes para el crecimiento del arbolado se calculó bajo las condiciones actuales de aprovechamiento (Martiarena, Wallis, & Knebel, 2009). Es decir, la cantidad disponible estimada de cada nutriente se dividió entre el requerimiento anual del rodal.
Análisis estadístico
Se hizo una comparación de medias de los datos dasométricos, tasa de descomposición de hojarasca, masa de nutrientes del suelo y piso forestal entre los dos rodales, a través de la prueba t de Student (P = 0.05). La información a nivel de rodal sobre requerimiento, absorción y reabsorción de nutrientes no se comparó por este método, ya que solo se genera un valor medio por rodal.
Resultados y discusión
La comparación de datos dasométricos, tasa de descomposición de hojarasca, masa de nutrientes del suelo y piso forestal entre los dos rodales no indicó diferencias estadísticas (t, P > 0.05); por esta razón, la discusión del estudio se enfocó principalmente en el análisis general, más que en la comparación de rodales. Aunque la estrategia estadística de tres repeticiones por rodal en este estudio es limitada, se consideró importante presentar y discutir los datos desde una perspectiva general, dada la importancia de promover estudios para la evaluación de la sostenibilidad en áreas forestales bajo aprovechamiento en México.
Datos dasométricos y nutrientes en biomasa
El rodal I tuvo una densidad de 180 árboles·ha-1 con valores promedio de diámetro normal (DN) de 39.5 cm, altura total de 25.3 m, edad de 86.3 años y volumen de 269 m3·ha-1. El rodal II tuvo una densidad de 125 árboles·ha-1, DN de 44 cm, altura total de 24.0 m, edad de 90.3 años y volumen de 224 m3·ha-1.
Los rodales I y II tuvieron biomasa arbórea aérea total de 133 y 120 Mg·ha-1, respectivamente, valores esperados para bosques de pino del centro de México (Mendoza-Ponce & Galicia, 2010). Los resultados de la muestra destructiva de árboles indicaron que la biomasa total se distribuyó porcentualmente de la siguiente forma: 89.5 (± 0.8), 7.5 (± 0.7) y 3 (± 0.4) para fuste, ramas y hojas, respectivamente. Estos valores, a excepción del follaje, son diferentes a los reportados por Keyes, Zárate, Martínez, y Garcidueñas (1988) para la especie estudiada; en el caso del fuste, la biomasa fue 8 % mayor, y en las ramas fue 60 % menor. La masa de N se distribuyó en el orden fuste, follaje y ramas (Cuadro 1), que es el patrón observado en otras especies de Pinus (Goya et al., 2003; Merino, Rey, Brañas, & Rodríguez-Soalleiro, 2003), mientras que la masa de Ca y Mg se concentró en el fuste y con cantidades similares en ramas y hojas.
Cuadro 1 Masa de nutrientes por componente del árbol en dos rodales de Pinus montezumae Lamb. en Nanacamilpa, Tlaxcala.
| Rodal | Componentes | N | P | K | Ca | Mg |
|---|---|---|---|---|---|---|
| (kg·ha-1) | ||||||
| Rodal I | Fuste | 156.7 | 4.5 | 7.3 | 148.3 | 101.5 |
| Ramas | 29.0 | 1.8 | 7.5 | 9.4 | 8.3 | |
| Follaje | 56.2 | 6.1 | 13.1 | 8.94 | 8.0 | |
| Total | 241.9 | 12.4 | 27.9 | 166.7 | 117.7 | |
| Rodal II | Fuste | 141.6 | 4.1 | 6.6 | 134.0 | 91.7 |
| Ramas | 26.2 | 1.6 | 6.8 | 8.5 | 7.5 | |
| Follaje | 50.8 | 5.5 | 11.8 | 8.1 | 7.2 | |
| Total | 218.6 | 11.2 | 25.3 | 150.6 | 106.4 | |
| CV (%) | Fuste | 6.0 | 45.1 | 32.4 | 7.7 | 13.5 |
| Ramas | 17.4 | 54.4 | 29.5 | 13.8 | 42.8 | |
| Follaje | 5.9 | 16.3 | 33.4 | 26.9 | 24.9 | |
No se encontraron diferencias estadísticamente significativas entre los rodales (t, P > 0.05). CV = coeficiente de variación de las determinaciones en laboratorio.
Nutrientes del piso forestal
La biomasa del piso forestal en los rodales I y II fue 22.2 y 21.5 Mg·ha-1, respectivamente. Estos valores son menores que los reportados por Merino et al. (2003) para Pinus radiata D. Don., y por Garcidueñas (1987) para P. montezumae. La diferencia en la biomasa de mantillo podría estar relacionada con la edad de los rodales y la densidad de árboles, ya que los estudios citados se hicieron en rodales más jóvenes y con mayor densidad. Aunque no se encontraron datos de la misma especie para fines de comparación, los resultados muestran valores esperados para comunidades de pino; por ejemplo, en bosques maduros de Pinus hartwegii Lindl. del centro de México se han estimado 18.7 Mg·ha-1 en el piso forestal (Mendoza-Ponce & Galicia, 2010).
La masa de nutrientes del piso forestal en el rodal I fue de 150.2, 9.6, 7.9, 95.2 y 40.6 kg·ha-1 de N, P, K, Ca y Mg, respectivamente; y en el rodal II, en el mismo orden, fue de 173.6, 9.4, 8.3, 91.6 y 34.4 kg·ha-1. Al respecto, Garcidueñas (1987) reportó valores mayores en la masa de nutrientes del piso forestal, debido a que la concentración de nutrientes en mantillo fue mayor. Otra posible razón puede ser la velocidad de descomposición de la hojarasca. Aunque la precipitación media anual en el área del estudio citado es mayor (> 1 000 mm), la temperatura media anual fue 6 °C menor que la del presente estudio (8 °C); de acuerdo con Landsberg y Gower (1997), las temperaturas bajas limitan la descomposición. En una plantación de Pinus patula Schltdl. & Cham. de 42 años, el principal reservorio de nutrientes fue el piso forestal; esto resalta la importancia de la hojarasca como reservorio de nutrientes en el suelo (Dames, Scholes, & Straker, 2002).
Suelo mineral
Se identificaron cuatro horizontes en el perfil del suelo de cada rodal (A, AB, Bw y Bt). Las profundidades de los suelos hasta el horizonte C y la masa de nutrientes en el suelo mineral se muestran en el Cuadro 2. La cantidad en masa de nutrientes está influenciada por la profundidad de los horizontes y por la densidad aparente que aumentó con la profundidad del suelo de 0.68 a 1.07 Mg·m-3. Las estimaciones totales de nutrientes, a una profundidad de 80 cm, fueron similares para ambos rodales con valores promedio de 5 419, 68, 1 245, 9 163 y 2 029 kg·ha-1 de N, P, K, Ca y Mg, respectivamente (Cuadro 2).
Cuadro 2 Masa de nutrientes en el suelo mineral de dos rodales de Pinus montezumae Lamb. en Nanacamilpa, Tlaxcala.
| Rodal | Horizontes | N | P | K | Ca | Mg |
|---|---|---|---|---|---|---|
| (kg·ha-1) | ||||||
| I | A (0-17 cm) | 1 693.4 | 7.0 | 179.4 | 1 818.3 | 279.9 |
| (12.0) | (54.6) | (5.26) | (15.5) | (13.4) | ||
| AB (17-40 cm) | 2 280.4 | 9.1 | 289.6 | 2 506.9 | 488.2 | |
| (35.2) | (91.1) | (8.8) | (10.3) | (40.6) | ||
| Bw (40-70 cm) | 1 026.5 | 41.7 | 521.9 | 3 393.1 | 856.7 | |
| (30.2) | (25.3) | (15.1) | (13.4) | (35.1) | ||
| Bt (70-115 cm) | 398.8 | 8.9 | 258.5 | 1 353.1 | 400.1 | |
| - | - | - | - | - | ||
| Total estimado a 80 cm | 5 399.1 | 66.7 | 1249.4 | 9 071.4 | 2 024.9 | |
| II | A (0-15 cm) | 1 734.4 | 10.4 | 171.6 | 2 002.1 | 289.1 |
| (35.7) | (19.3) | (17.7) | (2.3) | (9.12) | ||
| AB (15-32 cm) | 1 549.5 | 3.1 | 93.9 | 1 024.4 | 200.3 | |
| (15.8) | (69.3) | (19.6) | (21.5) | (9.21) | ||
| Bw (32-63 cm) | 1 691.1 | 60.4 | 110.0 | 2 493.8 | 710.7 | |
| (28.4) | (74.6) | (58.0) | (16.7) | (25.4) | ||
| Bt (63-95 cm) | 662.4 | 20.9 | 278.9 | 1 874.5 | 741.3 | |
| - | - | - | - | - | ||
| Total estimado a 80 cm | 5 440.1 | 70.1 | 1241.6 | 9 255.2 | 2 034.1 | |
| Diferencia entre rodales (%) | (0.76) | (5.10) | (0.62) | (2.03) | (0.45) | |
Las estimaciones para el horizonte Bt se ajustaron a la profundidad de 80 cm. Coeficiente de variación entre paréntesis (%); la medición del horizonte Bt no tuvo repetición.
Caída y descomposición de hojarasca
La Figura 1 presenta la producción de hojarasca en los dos rodales de P. montezumae. La producción en ambos rodales fue similar con 6.3 Mg ha-1 año-1, con un máximo en marzo (1.3 Mg·ha-1) que corresponde a la temporada más seca. Rodríguez-Sanfuentes y Álvarez-Muñoz (2010) mencionan que el factor más común que determina la abscisión de las hojas es la respuesta de los árboles al estrés hídrico por las temporadas de sequía. En septiembre hubo otro incremento en la producción de hojarasca que coincidió con el término de la temporada de lluvias. Este segundo pico de producción en otoño también lo muestra Pseudotsuga menziesii (Mirb.) Franco (Portillo-Estrada et al., 2013). Los resultados de este estudio se encuentran en el intervalo reportado para otras especies de Pinus que va de 3.9 a 7.8 Mg·ha-1·año-1 (León, González, & Gallardo, 2011; Luna & Hernández, 2009; Ramírez-Correa, Zapata-Duque, León-Peláez, & González-Hernández, 2007). Los bosques de Pinus sylvestris L. y P. menziesii en Europa producen 2.86 y 4.34 Mg·ha-1·año-1 de mantillo (Portillo-Estrada et al., 2013). La cantidad de nutrientes que regresa al suelo se estimó en 49.4, 2.7, 2.2, 21.4 y 8.4 kg·ha-1·año-1 de N, P, K, Ca y Mg, respectivamente. La cantidad de P que regresó al suelo fue escasa; al respecto, se ha reportado que este elemento es de baja movilidad tanto en sistemas naturales como intensivos; por ejemplo, en plantaciones de P. radiata regresan 3.5 kg·ha-1·año-1 al suelo vía hojarasca (Rivaie, 2014). Aunque la cantidad de K también es escasa, generalmente hay regreso de este elemento al suelo a través del lavado de copa y tronco (Binkley & Fisher, 2013), el cual no se realizó en este estudio.
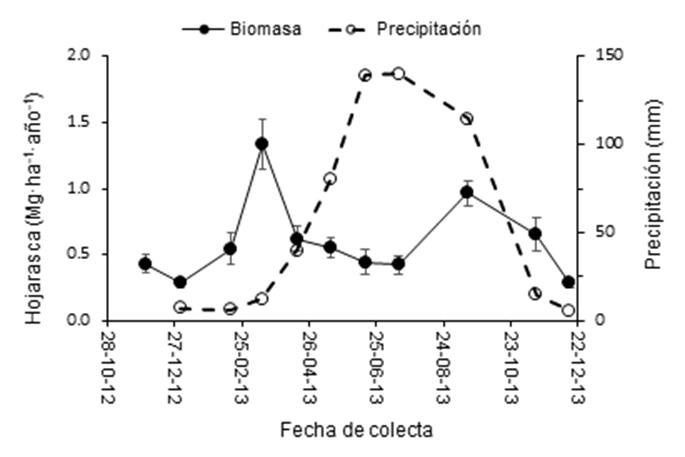
Figura 1 Producción de hojarasca media anual en el periodo 2012-2013 en dos rodales de Pinus montezumae en Nanacamilpa, Tlaxcala. Las líneas verticales representan el error estándar de la media.
Las tasas de descomposición de hojarasca fueron de -0.33 y -0.34 para el rodal I y II, respectivamente. Estos valores son los esperados para bosques templados (-0.14 a -0.69) (Landsberg & Gower, 1997). León et al. (2011) reportaron una tasa de descomposición de -0.5 para P. patula en sitios con 2 000 mm de precipitación, mientras que Rocha-Loredo y Ramirez-Marcial (2009) reportaron tasas de descomposición mayores de -0.5 en asociaciones de pino-encino al sur de México, donde la temperatura media anual varía de 14 a 25 °C.
Reabsorción de nutrientes
El N reabsorbido fue 39 % en el rodal I y 30 % en el II, para el P fue 36 % en el rodal I y 40 % en el II, y para K fue 51 % en el I y 53 % en el II. La masa de N, P y K retranslocada en el rodal I fue 21.9, 2.2 y 6.7 kg·ha-1·año-1, respectivamente, y en el rodal II fue 15.2, 2.2 y 6.2 kg·ha-1·año-1. Los valores de N y P son similares y los de K son 40 % menores que los encontrados en otras especies de Pinus (Dames et al., 2002; Martín, Santa Regina, & Gallardo, 1996; Peláez, Hernández, & Lancho, 2008; Pérez, Goya, Bianchini, Frangi, & Fernández, 2006). Comparado con Garcidueñas (1987), quien estudió P. montezumae, los valores de reabsorción de N y P son entre 1.7 y 2 veces menores. El resultado anterior podría deberse a que la retraslocación es mayor en rodales más jóvenes que en los maduros como el de este estudio. Dames et al. (2002) sugirieron que la limitación de P en el suelo se compensa con la tasa de reabsorción (40 % valor encontrado en este estudio), quizá como respuesta a la baja transferencia de este nutrimento del piso forestal al suelo mineral (Landsberg & Gower, 1997). La alta reabsorción de K puede estar asociada con su alta movilidad y no con la disponibilidad en el suelo (Martín et al., 1996).
Salida de nutrientes por la cosecha forestal
La salida de nutrientes por la cosecha forestal se muestra en el Cuadro 3. La biomasa extraída del bosque por la cosecha forestal es 5.17 Mg·ha-1·año-1 en el rodal I y 4.29 Mg·ha-1·año-1 en el rodal II. Estos valores son menores que los reportados en plantaciones de Pinus taeda L. de 20 años, bajo práctica de conservación, cosecha solo de fustes y retención del resto de residuos (Goya et al., 2003; Martiarena et al., 2009). El porcentaje de nutrientes que se extrae por la cosecha, con respecto a la magnitud del reservorio en biomasa aérea es bajo, no supera el 4.2 %; respecto al reservorio del piso forestal, las salidas representan 3 % para P y 13 % para Mg. No obstante, el problema de la conservación de la productividad del bosque radica en promover prácticas que estimulen la mineralización del material orgánico, para garantizar la provisión de nutrientes, sobre todo en el reciclaje del P (Maynard et al., 2014).
Cuadro 3 Masa promedio de nutrientes extraídos anualmente por el aprovechamiento forestal en dos rodales de Pinus montezumae Lamb. en el ejido de Nanacamilpa, Tlaxcala.
| Rodal | Componentes | Biomasa | N | P | K | Ca | Mg |
|---|---|---|---|---|---|---|---|
| kg·ha-1·año-1 | |||||||
| I | Fuste | 3 909 | 5.1 | 0.15 | 0.24 | 4.9 | 3.3 |
| Ramas | 386 | 1.1 | 0.07 | 0.29 | 0.4 | 0.3 | |
| Total | 4 296 | 6.3 | 0.22 | 0.53 | 5.2 | 3.6 | |
| II | Fuste | 4 708 | 6.2 | 0.18 | 0.29 | 5.9 | 4.0 |
| Ramas | 465 | 1.4 | 0.08 | 0.35 | 0.4 | 0.4 | |
| Total | 5 174 | 7.5 | 0.26 | 0.64 | 6.3 | 4.4 | |
Requerimiento de nutrientes y reservas potenciales
El crecimiento promedio anual expresado en biomasa fue de 5.3 y 4.8 Mg·ha-1, en los rodales I y II, respectivamente. Estos valores son ligeramente menores que el reportado por Garcidueñas (1987) para la misma especie, quizá porque dicho estudio se realizó en una plantación donde las prácticas silvícolas fueron más controladas, lo cual influye en el crecimiento más rápido del árbol. Con base en las determinaciones del presente estudio, las cantidades de N, P, K que el arbolado requiere para su crecimiento son de 73 a 81, 5.2 a 5.3 y 9.4 a 10 kg·ha-1·año-1, respectivamente (Cuadro 4). Tomando en cuenta que el nitrógeno potencialmente mineralizable es aproximadamente 3 % del nitrógeno total (Schlesinger & Bernhardt, 2013), los resultados indican que los 5 399 kg de N estimados en este estudio representan 162 kg de N mineralizable; es decir, el suelo abastece hasta dos veces los requerimientos anuales de la masa arbórea. Por otra parte, los resultados resaltan la importancia de mantener el material orgánico superficial del suelo y sugerir algunas prácticas silvícolas más intensivas, como aclareos fuertes y tempranos, para exponer la hojarasca y acelerar su descomposición.
Cuadro 4 Absorción y requerimiento anual de nutrientes en dos rodales de Pinus montezumae Lamb. en el ejido de Nanacamilpa, Tlaxcala.
| Rodal | Nutriente | GNA | RPF | Absorción | Reabsorción | Requerimiento |
|---|---|---|---|---|---|---|
| (kg·ha-1·año-1) | ||||||
| I | N | 9.7 | 49.5 | 59.2 | 21.9 | 81.1 |
| P | 0.5 | 2.6 | 3.1 | 2.2 | 5.3 | |
| K | 1.1 | 2.2 | 3.3 | 6.7 | 10.0 | |
| II | N | 8.7 | 49.5 | 58.2 | 15.3 | 73.5 |
| P | 0.4 | 2.6 | 3.0 | 2.2 | 5.2 | |
| K | 1.01 | 2.2 | 3.2 | 6.2 | 9.4 | |
GNA: ganancia neta anual, RPF: retorno al piso forestal
Bajo las condiciones actuales, el tiempo que los nutrientes del suelo podrían sostener el crecimiento del arbolado varía entre 256 y 1 952 años. El K es el nutrimento que mayor tiempo podría mantener el sistema, entre 1 234 a 1 952 años; seguido de Ca, entre 1 422 a 1 439 años; N entre 706 a 909 años; Mg entre 458 a 539 años; mientras que para P es de 256 a 430 años. De acuerdo con los resultados, las condiciones actuales de aprovechamiento pueden considerarse como sostenibles, ya que los nutrientes evaluados podrían mantener el crecimiento de la vegetación por 256 años. Aun cuando este cálculo está basado en una simplificación, es útil como una referencia preliminar, dada la falta de información para los bosques aprovechados en México. No obstante, el término sostenible en realidad es poco riguroso, ya que cada especialista lo interpreta de manera distinta. En sentido estricto, un sistema sostenible mantiene la productividad de manera permanente. Este concepto no aplica a un sistema forestal, ya que las comunidades arbóreas están sujetas a cambio. Una escala de tiempo para definir la sustentabilidad a largo plazo en un sistema forestal se estima de 150 a 200 años (Patzek & Pimentel, 2005). Por lo anterior, los resultados indican que, si no hay un manejo adecuado, el P es el elemento más probable que podría impactar la sustentabilidad del bosque estudiado.
Conclusiones
La cantidad de nutrientes del suelo en los rodales de Pinus montezumae superó la cantidad de nutrientes en la biomasa aérea. Las tasas de descomposición de hojarasca de -0.33 y -0.34 son las esperadas para ecosistemas templados. Dichas tasas indican que el mantillo tarda en promedio tres años en descomponerse, asumiendo que la cantidad de este se mantiene estable. Las reservas de nutrientes en piso forestal y suelo mineral son suficientes para la intensidad de aprovechamiento actual. El nitrógeno potencialmente mineralizable se estimó en dos veces los requerimientos de la masa arbolada. La transferencia de mantillo al piso forestal y la alta reabsorción de los macroelementos podrían satisfacer los requerimientos nutrimentales de los rodales estudiados. Si las condiciones actuales se mantienen, el manejo de los rodales de P. montezumae podría continuar por más de dos siglos, pero si las prácticas sustentables de cosecha dejaran de utilizarse, el fósforo sería el primer elemento que pondría en riesgo tal sustentabilidad. A pesar de ser un estudio limitado en rodales y repeticiones, la información que el estudio generó es relevante y se espera que pueda detonar el planteamiento de otros trabajos para evaluar la sostenibilidad del aprovechamiento forestal en México.
Agradecimientos
Al Consejo Nacional de Ciencia y Tecnología por el apoyo financiero de estudios de maestría de la primera autora. Al ejido de Nanacamilpa, Tlaxcala, por permitir el acceso a su área forestal.
REFERENCIAS
Binkley, D., & Fisher, R. (2013). Ecology and management of forest soils (4th ed.) UK: Wiley-Blackwell. [ Links ]
Bärlocher, F. (2005). Leaf mass loss estimated by litter bag technique. In A. S. Graca , F. Bärlocher, & M. O. Gessner (Eds.), Methods to study litter decomposition: a practical guide (pp. 36-42). Dordrecht, Alemania: Springer. doi: 10.1007/1-4020-3466-0_6347p [ Links ]
Dames, J. F., Scholes, M. C., & Straker, C. J. (2002). Nutrient cycling in a Pinus patula plantation in the Mpumalanga Province, South Africa. Applied Soil Ecology, 20(3), 211-226. doi: 10.1016/S0929-1393(02)00028-8 [ Links ]
Eric III. (2013). Versión 3.2 - extractor rápido de información climatológica. México: Instituto Mexicano de Tecnología del Agua. Retrieved from https://www.imta.gob.mx/productos/software/eric-iii-version-3-2-extractor-rapido-de-informacion-climatolo-detail [ Links ]
Garcidueñas, M. A. R. (1987). Biomass production and nutrimental accumulation in a Pinus montezumae Lamb. Retrieved January 7, 2016 from http://agris.fao.org/org/agris-search [ Links ]
Gomez, A., Powers, R., Singer, M., & Horwath, W. (2002). Soil compaction effects on growth of young ponderosa pine following litter removal in California's Sierra Nevada. Soil Science Society of America Journal, 66(4), 1334-1343. doi: 10.2136/sssaj2002.1334 [ Links ]
Goya, J. F., Pérez, C., Frangi, J. L., & Fernández, R. (2003). Impacto de la cosecha y destino de los residuos sobre la estabilidad del capital de nutrientes en plantaciones de Pinus taeda L. Ecología Austral, 13, 139-150. Retrieved from http://www.scielo.org.ar/pdf/ecoaus/v13n2/v13n2a03.pdf [ Links ]
Grigal, D. F. (2000). Effects of extensive forest management on soil productivity. Forest Ecology and Managment, 138(1-3), 167-185. doi:10.1016/S0378-1127(00)00395-9 [ Links ]
Hernández-Díaz, J. C., Corral-Rivas, J. J., Quiñones-Chávez, A., Bacon-Sobbe, J. R., & Vargas-Larreta, B. (2008). Evaluación del manejo forestal regular e irregular en bosques de la Sierra Madre Occidental. Madera y Bosques,14, 25-41. Retrieved from http://www.scielo.org.mx/pdf/mb/v14n3/v14n3a2.pdf [ Links ]
Imbert, J. B., Blanco, J. A., & Castillo, F. J. (2004). Gestión forestal y ciclos de nutrientes en el marco del cambio global. In F. Valladares (Ed.), Ecología del bosque mediterráneo en un mundo cambiante (pp. 479-506). Madrid, España: Ministerio de Medio Ambiente - EGRAF, S. A. Retrieved from http://www.adaptecca.es/sites/default/files/documentos/cap17_-_gestion_forestal_y_ciclos_de_nutrientes_en_el_marco_del_cambio_global.pdf [ Links ]
Johnson, D. W., Sogn, T., & Kvindesland, S. (2000). The nutrient cycling model: lessons learned. Forest Ecology and Management, 138(1-3), 91-106. doi: 10.1016/S0378-1127(00)00414-X [ Links ]
Keyes, H. M. R., Zárate, D. L. G., Martínez, M. M., & Garcidueñas, M. A. R. (1988). Estimación de biomasa para árboles de Pinus montezumae Lamb. Agrociencia, 72, 27-40. [ Links ]
Landsberg, J. J., & Gower, S. T. (1997). Applications of physiological ecology to forest management. USA: Academic Press. [ Links ]
León, J. D., González, M. I., & Gallardo, J. F. (2011). Ciclos biogeoquímicos en bosques naturales y plantaciones de coníferas en ecosistemas de alta montaña de Colombia. Revista Biología Tropical, 59(4), 1883-1894. Retrieved from http://www.scielo.sa.cr/pdf/rbt/v59n4/a37v59n4.pdf [ Links ]
Luna, J. A. N., & Hernández, E. H. (2009). Acumulación de biomasa aérea en un bosque coetáneo de la región de El Salto, Durango. Ra Ximhai, 5, 225-230. Retrieved from http://uaim.edu.mx/webraximhai/Ej-14articulosPDF/07BIOMASA.pdf [ Links ]
Machado, M. R., Sampaio, P. D. T. B., Ferraz, J., Camara, R., & Pereira, M. G. (2016). Nutrient retranslocation in forest species in the Brazilian Amazon. Acta Scientiarum. Agronomy, 38(1), 93-101. doi: 10.4025/actasciagron.v38i1.26805 [ Links ]
Martiarena, R., Von Wallis, A., & Knebel, O. (2009). Pérdida de nutrientes durante la cosecha y el establecimiento forestal, en un rodal de Pinus taeda en Misiones, Argentina. Revista Forestal Venezolana, 53(2), 165-173. Retrieved from http://inta.gob.ar/sites/default/files/script-tmp-perdidanutrientes_cosecha_ptaeda.pdf [ Links ]
Martín, A., Santa Regina, I., & Gallardo, J. F. (1996). Eficiencia, retraslocación y balance de nutrientes en bosques de Quercus pyrenaica bajo diferente pluviometría en la Sierra de Gata (Centro-Oeste Español). Ecología, 10, 79-93. Retrieved from http://digital.csic.es/bitstream/10261/34603/1/Ecologia199679.pdf [ Links ]
Maynard, D., Paré, D., Thiffault, E., Lafleur, B., Hogg, K., & Kishchuk, B. (2014). How do natural disturbances and human activities affect soils and tree nutrition and growth in the Canadian boreal forest? Environmental Reviews, 22(2), 161-178. doi: 10.1139/er-2013-0057 [ Links ]
Mendoza-Ponce, A., & Galicia, L. (2010). Aboveground and belowground biomass and carbon pools in highland temperate forest landscape in Central Mexico. Forestry, 83(5), 497-506. doi: 10.1093/forestry/cpq032 [ Links ]
Merino, A., Rey, C., Brañas, J., & Rodríguez-Soalleiro, R. (2003). Biomasa arbórea y acumulación de nutrientes en plantaciones de Pinus radiata D. Don en Galicia. Investigación Agraria. Sistemas y Recursos Forestales, 12, 85-98. Retrieved from https://www.researchgate.net/profile/Roque_Rodriguez-Soalleiro/publication/28063125_ [ Links ]
Patzek, T. W., & Pimentel, D. (2005). Thermodynamics of energy production from biomass. Critical Reviews in Plant Sciences, 24(5-6), 327-364. doi: 10.1080/07352680500316029 [ Links ]
Peláez, J. L., Hernández, M. G., & Lancho, J. G. (2008). Retranslocación y eficiencia en el uso de nutrientes en bosques del centro de Antioquia. Colombia Forestal, 12, 119-140. Retrieved from http://www.scielo.org.co/pdf/cofo/v12n1/v12n1a09.pdf [ Links ]
Pérez, C. A., Goya, J. F., Bianchini, F., Frangi, J. L., & Fernández, R. (2006). Productividad aérea y ciclo de nutrientes en plantaciones de Pinus taeda L. en el norte de la provincia de Misiones, Argentina. Interciencia, 31, 794-801. Retrieved from http://www.redalyc.org/html/339/33912207/ [ Links ]
Portillo-Estrada, M., Korhonen, J. F., Pihlatie, M., Pumpanen, J., Frumau, A. K., Morillas, L., …Niinemets, Ü. (2013). Inter-and intra-annual variations in canopy fine litterfall and carbon and nitrogen inputs to the forest floor in two European coniferous forests. Annals of Forest Science, 70(4), 367-379. doi: 10.1007/s13595-013-0273-0 [ Links ]
Ramírez-Correa, J. A., Zapata-Duque, C. M., León-Peláez, J. D., & González-Hernández, M. I. (2007). Caída de hojarasca y retorno de nutrientes en bosques montanos andinos de Piedras Blancas, Antioquia, Colombia. Interciencia , 32, 303-311. Retrieved from http://www.redalyc.org/html/339/33932503/ [ Links ]
Rivaie, A. A. (2014). The effects of understory vegetation on P availability in Pinus radiata forest stands: A review. Journal of Forestry Research, 25(3), 489-500. doi: 10.1007/s11676-014-0488-4 [ Links ]
Rodríguez-Sanfuentes, J., & Álvarez-Muñoz, J. (2010). Nutrición y fertilización de plantaciones forestales. Santiago, Chile: Gráfica LOM. [ Links ]
Rocha-Loredo, G. A., & Ramirez-Marcial, N. (2009). Litter production and leaf-litter decomposition in different pine-oak forest successional conditions in Chiapas, Mexico. Boletín de la Sociedad Botánica de México, 84, 1-12. Retrieved from http://www.scielo.org.mx/pdf/bsbm/n84/n84a1.pdf [ Links ]
Romahn, V. C. F., & Ramírez, M. H. (2010). Dendrometría. México: Universidad Autónoma Chapingo. [ Links ]
Schlesinger, W. H., & Bernhardt, E. S. (2013). Biogeochemistry: An analysis of global change (3rd ed.) San Diego, California,USA: Academic Press . [ Links ]
Secretaría de Medio Ambiente y Recursos Naturales (2002). Norma Oficial Mexicana NOM-021-RECNAT-2000 que establece las especificaciones de fertilidad,sanidad y clasificación de suelos, estudios, muestreo y análisis. México: DOF. Retrieved from http://biblioteca.semarnat.gob.mx/janium/Documentos/Ciga/libros2009/DO2280n.pdf [ Links ]
Sollins, P., Grier, C., Mccorison, F., Cromack, K., Fogel, R., & Fredriksen, R. (1980). The internal element cycles of an old‐growth Douglas‐fir ecosystem in western Oregon. Ecological Monographs, 50, 261-285. doi: 10.2307/2937252 [ Links ]
Zepeda, B. E. M., & Acosta, M. M. (2000). Incremento y rendimiento maderable de Pinus montezumae Lamb., en San Juan Tetla, Puebla. Madera y Bosques , 6(1), 15-27. doi: 10.21829/myb.2000.611339 [ Links ]
Recibido: 05 de Septiembre de 2017; Aprobado: 24 de Noviembre de 2017
 This is an open-access article distributed under the terms of the Creative Commons Attribution License
This is an open-access article distributed under the terms of the Creative Commons Attribution License
