










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista Chapingo serie ciencias forestales y del ambiente
On-line version ISSN 2007-4018Print version ISSN 2007-3828
Rev. Chapingo ser. cienc. for. ambient vol.18 n.3 Chapingo Aug./Dec. 2012
https://doi.org/10.5154/r.rchscfa.2010.11.101
Caracterización estructural del manglar en el estero Palo Verde, laguna de Cuyutlán, Colima
Structural characterization of the mangrove forest in the estero Palo Verde, Cuyutlán lagoon, Colima
Citlalli P. Téllez-García1; Juan I. Valdez-Hernández2*
1 División de Ciencias Forestales Universidad Autónoma Chapingo - Restauración Forestal. km 38.5 Carretera México-Texcoco, Chapingo, Estado de México. C. P. 56230.
2 Programa Forestal. Colegio de Postgraduados. km 36.5 Carretera México-Texcoco, Montecillo, Estado de México. C. P. 56230, Correo-e: ignaciov@colpos.mx (*Autor para correspondencia).
Recibido: 25 de octubre de 2010
Aceptado: 2 de mayo de 2012
RESUMEN
En el Estero Palo Verde, ubicado en el vaso IV de la Laguna de Cuyutlán, estado de Colima, se establecieron 48 unidades de muestreo (UM) en tres rodales de mangle: 10 UM en rodal monoespecífico de Laguncularia racemosa, 25 UM en rodal monoespecífico de Rhizophora mangle y 13 UM en rodal mixto de L. racemosa-R. mangle. Se registró altura del dosel, diámetro del tronco, densidad de adultos y de repoblación, también se calculó área basal y volumen de madera. Los resultados indicaron que la densidad, área basal y volumen, para la categoría diamétrica (CD) de 5 cm fueron significativamente diferentes (P < 0.05) entre los rodales monoespecíficos de R. mangle (2,500 árboles·ha-1, 4.2 m2·ha-1 y 22 m3·ha-1, respectivamente) y L. racemosa (700 árboles·ha1, 1.9 m2·ha-1 y 10 m3·ha-1, respectivamente). En las CD de 20 a 30 cm, el área basal fue significativamente mayor (P < 0.05) en el rodal monoespecífico de L. racemosa (1.6 - 3.8 m2·ha-1) que en el mixto de L. racemosa-R. mangle (0.5 - 2.5 m2·ha-1). La repoblación natural fue más abundante en el rodal de R. mangle (138 a 270 individuos·ha-1) que en el de L. racemosa (70 a 80 individuos·ha-1). Los valores estructurales mayores se alcanzaron para altura del dosel (10 m) en el rodal de L. racemosa; para diámetro (40 cm) y área basal (14.1 m2·ha-1) en el rodal de L. racemosa-R. mangle; y para repoblación natural (624 individuos·ha-1) en el rodal de R. mangle.
Palabras Clave: Rodal monoespecífico y mixto, Laguncularia racemosa, Rhizophora mangle, características dasonómicas, repoblación natural, categorías
ABSTRACT
In the Estero Palo Verde, located in section IV of the Cuyutlán Lagoon, state of Colima, 48 sample units (SU) were established on three types of mangrove stands: 10 SU on pure stand of Laguncularia racemosa, 25 SU on pure stand of Rhizophora mangle and 13 SU on mixed forest stand of L. racemosa-R. mangle. Canopy height, trunk diameter, density of adults and natural regeneration were recorded, also basal area and tree volume were calculated. Results indicated that density, basal area and tree volume for diameter category (DC) of 5 cm were significantly (P < 0.05) different between the pure stands of R. mangle (2500 trees·ha1, 4.2 m2·ha-1, 22 m3·ha-1) and L. racemosa (700 trees·ha1, 1.9 m2·ha-1, 10 m3·ha-1). In the case of DC of 20 to 30 cm, the basal area was significantly (P < 0.05) higher in the pure stand of L. racemosa (1.6 - 3.8 m2·ha-1) than in the mixed forest stand of L. racemosa-R. mangle (0.5 - 2.5 m2·ha-1). Natural regeneration was more abundant in the stand of R. mangle (138 -270 individuals·ha-1) than in the stand of L. racemosa (70 - 80 individuals·ha1). The highest structural values were reached for canopy height (10 m) in the stand of L. racemosa, for trunk diameter (40 cm) and basal area (14.1 m2·ha-1) in the stand of L. racemosa-R. mangle, and for natural regeneration (624 individuals·ha-1) in the stand of R. mangle.
Keywords: Pure and mixed forest stands, Laguncularia racemosa, Rhizophora mangle, forest characteristics, natural regeneration, diameter
INTRODUCCIÓN
Los manglares constituyen un recurso forestal importante en las costas tropicales y subtropicales del planeta. Cubren una superficie aproximada de 15.2 millones de hectáreas y han sido ampliamente utilizados en la mayoría de los países en los que existen (Food and Agriculture Organization [FAO], 1994, 2007).
El conocimiento de las condiciones anteriores y actuales de los manglares, es esencial para la toma de decisiones en su conservación y manejo sustentable. Como ejemplo, la falta de información básica en estructura de la vegetación y repoblación natural son los principales obstáculos para la conservación de manglares en Haití (M'elanie & Lise, 2001), Sri Lanka (Dahdouh-Guebas, Gitundu, Jayatissa, Cannicci, &I Koedam, 2002) y Kenya (Mohamed, Neukermans, Gitundu, Dahdouh-Guebas, & Koedam, 2009), ya que estos datos permiten conocer mejor la dinámica forestal y facilitan la formulación de pautas de manejo. En Brasil, a pesar de la presencia de grandes zonas de manglares en la costa de Río de Janeiro, el estudio de sus atributos estructurales y funcionales es casi inexistente (Soares, 1999).
Para manglares de México se tiene información de estructura de la vegetación en la costa del Pacífico (Flores-Verdugo, González-Farías, Segura-Zamorano, & Ramírez-García, 1992), estructura forestal en Tabasco (Corella et al., 2001), estructura forestal y repoblación natural en Nayarit (Valdez, 2002), así como estructura de la vegetación en Yucatán (Zaldivar, Herrera, Coronado, & Alonzo, 2004). No obstante, que estos datos son fundamentales para comprender la demografía y estructura de tamaños en comunidades naturales y manejadas (López & Ezcurra, 2002), no se tienen para gran parte de los manglares del país, particularmente del estado de Colima.
Por lo anterior, los objetivos del presente trabajo fueron describir la estructura forestal y evaluar la repoblación natural en rodales de mangles del Estero Palo Verde, Laguna de Cuyutlán, Colima, con el propósito de contribuir a su manejo adecuado. Se plantea la hipótesis de que no existen diferencias significativas en densidad de individuos entre clases de altura y categorías diamétricas para un mismo rodal o entre rodales para una misma clase de altura y categoría diamétrica.
MATERIALES Y MÉTODOS
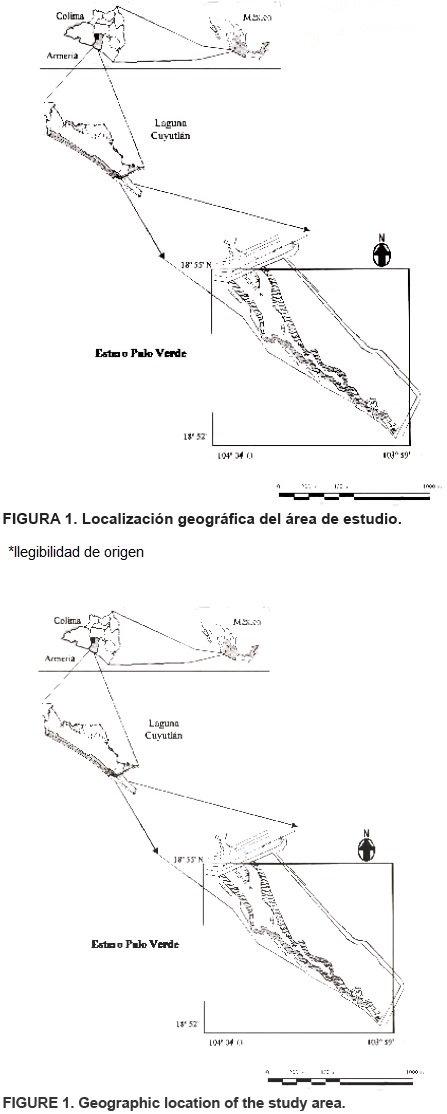
La investigación se realizó en el Estero Palo Verde, en el vaso IV de la Laguna de Cuyutlán, estado de Colima; localizado entre los 18° 52' a 18° 55' latitud norte y 103° 59' a 104° 04' longitud oeste (Figura 1). El clima del área de estudio es semiseco-cálido con lluvias en verano; BS1 (h') wi w''. La temperatura media anual es de 26 °C, con una máxima de 28 °C y mínima de 22 a 23 °C, la precipitación media anual es de 800 a 1,200 mm (García, 1987).
La vegetación predominante se compone de manglar (Rhizophora mangle, Laguncularia racemosa) con vegetación halófita (Batis marítima) (Instituto Nacional de Estadística y Geografía [INEGI], 1994), y el suelo, de origen aluvial, es clasificado como fluvisol éutrico (INEGI, 1995).
Mediciones en campo. Se establecieron, de manera aleatoria, 48 unidades de muestreo (UM) de 30 x 10 m en tres rodales de mangle: 10 UM en rodal monoespecífico de L. racemosa, 25 UM en rodal monoespecífico de R. mangle y 13 UM en rodal mixto de L. racemosa-R. mangle. Considerando el tipo de información a registrar (Valdez, 2002), cada UM fue subdividida en tres cuadros de 10 x 10 m (100 m2), sobre los que se establecieron en forma aleatoria dos cuadros de 4 x 4 m (16 m2) y cinco de 1 x 1 m (1 m2). En los cuadros de 100 m2 se registró el diámetro normal (DN) de todos los individuos mayores a 2.5 cm de DN; en los de 16 m2 se midió la densidad de los tallos menores a 2.5 cm de DN pero mayores que 1.3 m de altura; y en los de 1 m2 la densidad de los tallos que se encontraban en un intervalo de 0.30 m a 1.3 m de altura, así como el total de individuos menores a 30 cm de altura.
La altura de los árboles se obtuvo utilizando una pistola Haga. Los diámetros se midieron con una cinta diamétrica a una altura de 1.3 m del suelo y 20 cm arriba de la parte final de las raíces zancudas en R. mangle. Las UM se delimitaron con una cuerda. En cada UM se registró el total de individuos muertos, así como la altura y diámetro de éstos.
Cálculos. La información se organizó por especie, clase de altura (5 m de amplitud) y categoría diamétrica (5 cm de amplitud). Se tuvieron dos clases de altura: 5 y 10 m. La clase 5 corresponde a aquéllos árboles que midieron de 2.5 a 7.4 m de altura y la clase 10 a los árboles que midieron de 7.5 a 12.4 m. En el caso de la categoría diamétrica, la categoría 5 se refiere a aquéllos árboles que midieron de 2.5 a 7.4 cm de DN, la categoría 10 a los que midieron de 7.5 a 12.4 cm de DN, así sucesivamente, hasta la clase 40. El área basal de los tallos mayores a 2.5 cm de DN fue calculada con la siguiente fórmula:
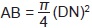
Donde:
AB = Área basal (m2)
DN = Diámetro Normal (1.3 m)
El volumen del arbolado se calculó mediante la fórmula consultada en Valdez (2004). Cabe mencionar que dicha fórmula fue generada para árboles de L. racemo-sa, pero en este caso también se aplicó para árboles de R. mangle.
V = 0.00543946 + 0.00003622 (DN2 H)
Donde:
V = Volumen (m3)
DN = Diámetro Normal (m)
H = Altura total (m)
Se utilizaron dos índices de valoración estructural de los rodales: a) Índice de Valor de Importancia (IVI) y b) Índice de Valor Forestal (IVF).
El IVI es un índice de importancia estructural, desarrollado principalmente para rodales mezclados. Se calculó de acuerdo a la fórmula utilizada por Villavicencio-Enríquez y Valdez-Hernández (2003):
IVI = Dr + Domr + Fr
Donde:
IVI = Índice de Valor de Importancia (%)
Dr = Densidad relativa (%
Domr = Dominancia relativa (%)
Fr= Frecuencia relativa (%)
El IVF se aplicó con el propósito de considerar la altura de todos los individuos medidos en el rodal, así como la densidad de cobertura de dosel, mediante la fórmula utilizada por Corella et al. (2001):
IVF = DAPr + Ar + Cr
Donde:
IVF = Índice de Valor Forestal (%)
DAPr = Diámetro relativo a la altura del pecho (%)
Ar = Altura relativa (%)
Cr = Cobertura relativa (%)
Análisis estadístico. Se generó una base de datos con los atributos estructurales: densidad, área basal, altura del dosel y volumen; así como con el número de individuos muertos y de renuevo. Se elaboraron gráficas por especie para las variables de altura, diámetro, área basal, volumen y repoblación natural.
Se realizó un análisis de varianza con los datos promedio por rodal para cada atributo estructural (altura, diámetro, área basal, volumen, renuevos), distinguiendo individuos vivos y muertos. Los datos se analizaron en un diseño completamente al azar. Se hizo una comparación de medias con la prueba de Tukey (P < 0.05) para identificar posibles diferencias. El análisis se ejecutó con el paquete estadístico SAS (Statistical Analysis System [SAS] 1999).
RESULTADOS Y DISCUSIÓN
La especie con el valor mayor de Índice de Valor de Importancia en el rodal mixto fue L. racemosa (Cuadro 1). Este resultado coincide con lo reportado por Zaldívar et al. (2004) en laguna Celestún, Yucatán, donde la especie L. racemosa presentó dominancia mayor hacia la zona terrestre (IVI = 61 %), pero disminuyó hacia el borde de la laguna (IVI = 22 %) donde R. mangle fue dominante (IVI = 52 %).
También L. racemosa obtuvo el Índice de Valor Forestal más alto (169 %) en el rodal mixto (Cuadro 2), similar a lo señalado en Centla, Tabasco, por Corella et al. (2001).
En el presente trabajo se encontraron diferencias significativas (P < 0.05) para la variable densidad de individuos entre clases de altura y categorías diamétricas para un mismo rodal, así como entre rodales para una misma clase de altura y categoría diamétrica, por tal motivo, se rechaza la hipótesis inicialmente planteada.
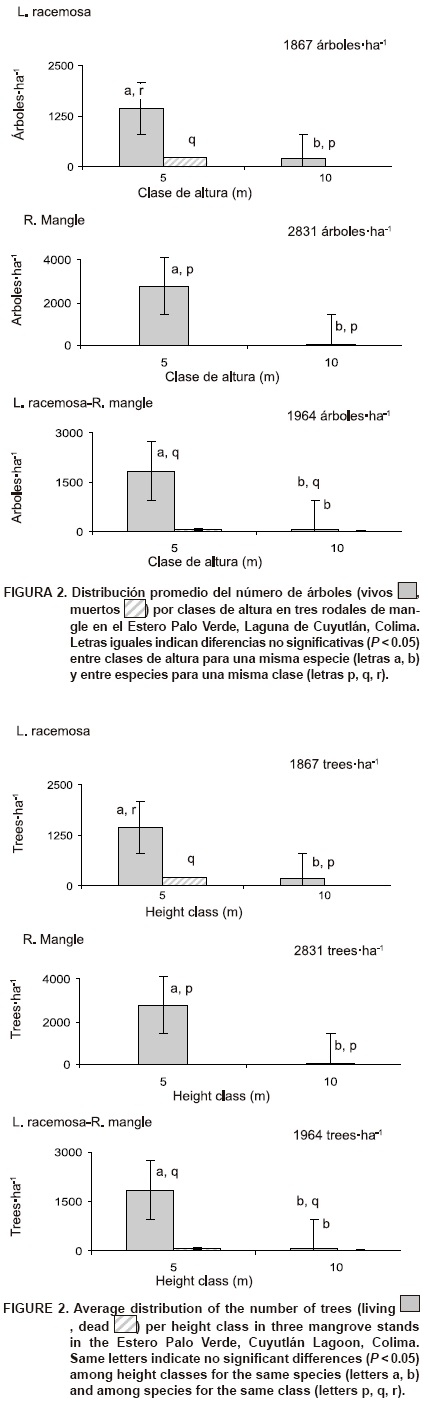
Altura y diámetro. Se presentaron diferencias significativas (P < 0.05) entre clases de altura para los tres rodales y entre los rodales para la clase 5 m; en contraste, los rodales monoespecíficos no fueron diferentes (P > 0.05) para la clase de altura 10 m (Figura 2). La altura del manglar en el área de estudio (11.8 m) fue mayor que la registrada en Haití (2 m) por M'elanie y Lise (2001), pero menor que la reportada en Tijuca, Brasil (16.1 m) por Soares (1999) y en Centla, Tabasco (23.5 m) por Corella et al. (2001). Este resultado puede estar relacionado, entre otros factores, con la precipitación anual (800-1,200 mm) en el área de estudio, ya que es menor que la señalada por Schaeffer-Novelli, Cintron-Molero, Rothleder y Camargo (1990) para que los manglares se desarrollen favorablemente (> 1,500 mm) y alcancen su máxima altura (> 2,500 mm). Además, Menéndez y Guzmán (2007) indican que la variación en altura del dosel depende de las condiciones de inundación y del sustrato.
Todas las categorías diamétricas presentaron diferencias significativas (P < 0.05) a excepción de las categorías 20 y 25 cm en el rodal de L. racemosa, 25 y 30 cm en el rodal de R. mangle, así como 15 y 20 cm en el rodal mixto de L. racemosa-R. mangle (Figura 3). El diámetro de los árboles en el área de estudio (18.5 cm) fue menor que los señalados en San Mateo, Brasil (22.9 cm), por Silva, Bernini y Carmo (2005) y en Centla, Tabasco (19.6 cm), por Corella et al. (2001). Esto quizá pueda deberse a un bajo aporte de nutrientes al manglar, ya que según lo expresado por Holguín et al. (1999), el flujo constante de nutrientes continentales a través de los ríos permite un buen desarrollo estructural de los árboles de mangle. Asimismo, Zaldívar et al. (2004) mencionan que factores ambientales como la salinidad del suelo y las descargas de agua subterránea son de importancia en la estructura de la vegetación.
Área basal y volumen. Los valores mayores para área basal y volumen se obtuvieron para el rodal de L. racemosa (19.1 m2·ha_1 y 68.9 m3·ha_1), seguidos por los del rodal de R. mangle (13.7 m2·ha_1 y 44.4 m3- ha1) y finalmente por aquéllos del rodal de L. racemosa-R. mangle (14.1 m2·ha-1 y 23 m3- ha-1) (Figuras 4 y 5). Sin embargo, estos valores fueron menores al compararlos con el área basal de manglares en Yucatán: 23.6 m2·ha-1 (Zaldívar et al., 2004), Tabasco: 24.4 m2·ha-1 (Corella et al., 2001), Brasil: 31.1 m2·ha-1 (Silva et al., 2005) y Cuba: 56.3 m2·ha-1 (Menéndez & Guzmán, 2007); así como menores en volumen que manglares de Nayarit: 86.6 m3·ha-1 (Valdez, 2004), Kenya: 153 m3·ha-1 (Gitundu, Dahdouh-Guebas, Giuada, Ochieng, & Koedom, 2002) e Indonesia: 226 m3·ha-1 (FAO, 2007). El área basal es un indicador del desarrollo de la estructura en un rodal. Conforme éste avanza en edad, posee menos árboles pero logra área basal mayor (Corella et al., 2001). Por lo tanto, se considera que el manglar en el Estero Palo Verde no ha alcanzado su madurez estructural.
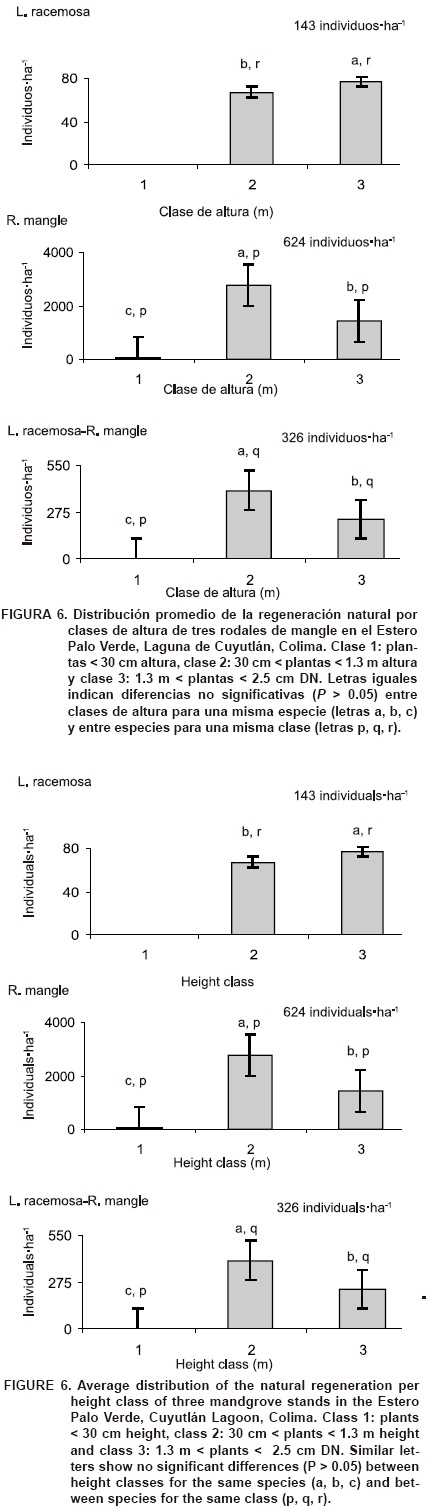
Repoblación natural. La densidad de renuevos fue mayor en el rodal monoespecífico de R. mangle (624 individuos·ha-1) que en el mixto de L. racemosa-R. mangle (326 individuos·ha-1) y que en el monoespecífico de L. racemosa (143 individuos·ha1). Datos similares fueron obtenidos en Nayarit (625 individuos·ha-1) por Valdez (2002) y en Kenya (461 individuos·ha-1) por Gitundu et al. (2002). Existen reportes que indican un crecimiento menor en plántulas de mangle bajo dosel cerrado (Koch, 1997) y la importancia que tiene la apertura de claros en los manglares para propiciar su renovación (Kathi-resan & Bingham, 2001). Incluso una alta mortalidad de plántulas no constituye un problema si existe abundante repoblación natural (Fonseca, Cortés, & Zamora, 2007).
Implicaciones de manejo. En el rodal monoespe-cífico de L. racemosa se presentó la densidad mayor de árboles muertos, principalmente en las categorías dia-métricas pequeñas: 5 y 10 cm (Figura 3). Los lugareños atribuyen la muerte de estos individuos a un aumento de salinidad por falta de entrada de agua del estero; sin embargo, no se tienen mediciones que permitan corroborar lo anterior. Además, la densidad de renuevos fue mucho menor en este rodal que en los otros dos y no se registraron individuos menores a 30 cm de altura (Figura 6). Por lo anterior y tomando en cuenta que reportes en otros sitios señalan que los mangles pueden morir por hundimiento del suelo (Jiménez, Lugo, & Cintron, 1985), altas tasas de sedimentación (Silva et al., 2005) o incremento en la salinidad (Menéndez & Guzmán, 2007), se recomienda considerar este rodal como zona de recuperación. Debido a esto, es necesario realizar mediciones tanto de las condiciones físicas (por ejemplo, salinidad del suelo y del agua) como de las biológicas (por ejemplo, daño por herbivoría o plagas) que permitan un diagnóstico adecuado para una aplicación correcta de futuras actividades de manejo.
El rodal monoespecífico de R. mangle es utilizado actualmente para recorridos turísticos en lancha, donde se menciona a los visitantes la importancia de la flora y fauna de los manglares. Sin embargo, faltan evaluaciones detalladas de los recursos escénicos y de las actividades recreativas (González & Valdez, 1993; González, Valdez, & Velasco, 1993) que permitan un manejo sustentable con fines tanto de esparcimiento como de educación ambiental.
Finalmente, y de acuerdo con el contenido alto de carbono orgánico (106.7 Mg C- ha-1) reportado en suelos de rodales de L. racemosa-R. mangle en Nayarit (Val-dés, Valdez, Gallardo, Ordaz, & Pérez, 2006), se sugiere destinar el rodal mixto del área de estudio como una zona de conservación de servicios ambientales, particularmente, como un reservorio de carbono orgánico.
CONCLUSIONES
Las características estructurales del manglar en el Estero Palo Verde, Laguna de Cuyutlán, Colima, fueron significativamente diferentes entre los rodales estudiados. La densidad, área basal y volumen fueron mayores, para la categoría diamétrica más pequeña (5 cm DAP), en el rodal monoespecífico de R. mangle que en el monoespecífico de L. racemosa. Sin embargo, para las categorías más grandes (20-30 cm DAP), el área basal fue mayor en el rodal monoespecífico de L. racemosa que en el mixto de L. racemosa-R. mangle.
La densidad de renuevos fue significativamente diferente (P < 0.05) entre clases de altura para un mismo rodal y entre rodales para una misma clase de altura. El rodal monoespecífico de R. mangle tuvo una mayor densidad de renuevos que el mixto de L. racemosa-R. mangle y que el monoespecífico de L. racemosa.
Con base en los resultados obtenidos en el presente trabajo se sugiere: a) considerar el rodal monoes-pecífico de L. racemosa como una zona de recuperación en su repoblación natural, b) mantener el rodal monoespecífico de R. mangle para uso turístico, pero aplicando técnicas de evaluación detalladas de recursos escénicos, y c) destinar el rodal mixto de L. racemosa-R. mangle como una zona de conservación de servicios ambientales (reservorio de carbono orgánico).
AGRADECIMIENTOS
Al Dr. Eduardo Valdés y M. C. Emilio Castillo, al personal del Centro Ecológico del Tortugario en Cuyutlán, Colima; así como a los árbitros designados por la revista para la revisión del presente trabajo.
REFERENCIAS
Corella J. F., Valdez H. J. I., Cetina A. V. M., González C. F., Trinidad S. A., & Aguirre Rivera, J. R. (2001). Estructura forestal de un bosque de mangles en el noreste del estado de Tabasco, México. Ciencia Forestal en México, 26(90), 73-102. [ Links ]
Dahdouh-Guebas, F., Gitundu, K. J., Jayatissa, I. P., Cannicci, S., & Koedam, N. (2002). An ordination study to view vegetation structure dynamics in disturbed and undisturbed mangrove forests in Kenya and Sri Lanka. Plant Ecology, 161, 123-135. doi: 10.1023/A:1020333706650 [ Links ]
Food and Agriculture Organization of the United Nations. (FAO). 1994. Mangrove forest management guidelines. Rome, Italy: USDA Forest Service. [ Links ]
Food and Agriculture Organization of the United Nations. (FAO). (2007). The world's mangroves 1980-2005. Rome, Italy: Autor. [ Links ]
Flores-Verdugo, F. J., González-Farías, F., Segura-Zamorano, D., & Ramírez-García, P. (1992). Mangrove ecosystems of the pacific coast of Mexico: Distribution, structure, litterfall, and detritus dynamics. In V. Seeliger (Ed.), Coastal plant communities of Latin America (pp. 269-288). New York, USA: Academic Press. [ Links ]
Fonseca, E. A. C., Cortés, J., & Zamora P. (2007). Monitoreo del manglar de Gandoca, Costa Rica (sitio CARICOMP). Revista de Biología Tropical, 55(1), 23-31. [ Links ]
García, E. (1987). Modificaciones al sistema de clasificación climática de Köppen. (4a ed.). México, D.F.: SIGSA. [ Links ]
Gitundu K. J., Dahdouh-Guebas, F., Gwada, P. O., Ochieng, C., & Koedam, N. (2002). Regeneration status of mangrove forests in Mida Creek, Kenya: A compromised or secured future? Ambio, 31(7/8), 562-568. [ Links ]
González G. M. J., & Valdez H. J. I. (1993). Análisis de metodologias y técnicas de inventario en áreas forestales recreativas. Agrociencia: Serie Recursos Naturales Renovables, 3(1), 61-80. [ Links ]
González G. M. J., & Velasco G. C. (1993). Valoración de la recreación en áreas forestales. Agrociencia: serie Recursos Naturales Renovables, 3(3), 95-113. [ Links ]
Holguín, G., Bashan, Y., Mendoza, S. R. A., Amador, E., Toledo, G., Vázquez, P., & Amador, A. (1999). La microbiología de los manglares: Bosques en la frontera entre el mar y la tierra. Ciencia y Desarrollo, 144, 27-35. [ Links ]
Instituto Nacional de Estadística y Geografía (INEGI) (1994). Armería, estado de Colima. Cuaderno Estadístico Municipal. México: Autor. [ Links ]
Instituto Nacional de Estadística y Geografía (INEGI) (1995). Estudio hidrológico del estado de Colima. México: Autor. [ Links ]
Jiménez, J. A., Lugo, A. E., & Cintron, M. G. (1985). Tree mortality in mangrove forests. Biotropica, 17(3), 177-185. doi: 10.2307/2388214. [ Links ]
Kathiresan, K., & Bingham, B. L. (2001). Biology of mangroves and mangrove ecosystems. Advances in Marine Biology, 40, 81-251. doi:10.1016/S0065-2881(01)40003-4. [ Links ]
Koch, M. S. (1997). Rhizophora mangle L. seedling development into the sapling stage across resource and stress gradients in subtropical Florida. Biotropica, 29(4), 427-439. doi: 10.1111/j.1744-7429.1997.tb00037.x [ Links ]
López Portillo, J., & Ezcurra, E. (2002). Los manglares de México: Una revisión. Madera y Bosques, Número especial, 27-51. [ Links ]
M'elanie, A., & Lise, C. (2001). The mangroves of the north coast of Haiti: A preliminary assessment. Wetlands Ecology and Management, 9, 271-278. doi:10.1023/A:1011173226724. [ Links ]
Menéndez C. L., & Guzmán, J. M. (2007). Ecosistemas de manglar en el archipiélago cubano. Cuba: Academia Cuba. [ Links ]
Mohamed, O. S., Neukermans, G., Gitundu, K. J., Dahdouh-Guebas, G. F., & Koedam, N. (2009). Mangrove forests in a peri-urban setting: The case of Mombasa (Kenya). Wetlands Ecology and Management, 17, 243-255. doi:10.1007/s11273-008-9104-8. [ Links ]
Statistical Analysis System (SAS). (1999). SAS Procedures Guide Version 6.0. Cary, North Carolina, USA. [ Links ]
Schaeffer-Novelli, Y., Cintron-Molero, G., Rothleder A. R., & Camargo, T. M. (1990). Variability of mangrove ecosystems along the Brazilian coast. Estuaries, 13(2), 204-218. doi:10.2307/1351590. [ Links ]
Silva, M. A. B., Bernini, E., & Carmo, T. M. S. (2005). Características estruturais de bosques de mangue do estuário do rio São Mateus, ES, Brasil. Acta Botánica Brasilica, 19 (3), 465-471. [ Links ]
Soares, M. L. G. (1999). Estrutura vegetal e grau de perturbação dos manguezais da Lagoa da Tijuca, Rio de Janeiro, RJ, Brasil. Revista Brasileira de Biologia, 59 (3), 503-515. doi:10.1590/S0034-71081999000300016. [ Links ]
Valdés V. E., Valdez H. J. I., Gallardo L. J. F., Ordaz C. V. M., & Pérez N. J. (2006). El carbono edáfico en manglares de Nayarit (México). In J. F. Gallardo Lancho (Ed.). Medioambiente en Iberoamérica: Visión desde la física y la química en los albores del siglo XXI (pp. 519-523). Badajoz, España: SIFYQA. [ Links ]
Valdez, H. J. I. (2002). Aprovechamiento forestal de manglares en el estado de Nayarit, costa Pacífica de México. Madera y Bosques, Número especial, 129-145. [ Links ]
Valdez H. J. I. (2004). Manejo forestal de un manglar al sur de Marismas Nacionales, Nayarit. Madera y Bosques, Número especial 2, 93-104. [ Links ]
Villavicencio-Enríquez, L., & Valdez-Hernández, J. I. (2003). Análisis de la estructura arbórea del sistema agroforestal rusticano de café en San Miguel, Veracruz, México. Agrociencia, 37(4), 413-423. [ Links ]
Zaldívar J. A., Herrera S. J., Coronado M. C., & Alonzo P. D. (2004). Estructura y productividad de los manglares en la reserva de biosfera Ría Celestún, Yucatán, México. Madera y Bosques, Número especial 2, 25-35. [ Links ]

