










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista Chapingo serie ciencias forestales y del ambiente
On-line version ISSN 2007-4018Print version ISSN 2007-3828
Rev. Chapingo ser. cienc. for. ambient vol.17 n.2 Chapingo Aug. 2011
https://doi.org/10.5154/r.rchscfa.2010.11.117
Ecuaciones alométricas para estimar biomasa y carbono en Quercus magnoliaefolia
Allometric equations for estimating the above–ground biomass and carbon in Quercus magnoliaefolia Née
Jesús D. Gómez–Díaz1¶; Jorge D. Etchevers–Barra2; Alejandro I. Monterrosos–Rivas1; Julio Campo–Alvez3; Juan A. Tinoco–Rueda1
1 Departamento de Suelos, Universidad Autónoma Chapingo, km 38.5 Carretera México–Texcoco, Chapingo, Estado de México, C. P. 56230. (¶Autor para correspondencia) Correo–e: dgomez@correo.chapingo.mx.
2 Instituto de Recursos Naturales, Colegio de Postgraduados, km 36.5 Carretera México–Texcoco, Montecillo, México.
3 Instituto de Ecología, Universidad Nacional Autónoma de México, México, D.F.
Recibido: 22 de noviembre, 2010
Aceptado: 07 de mayo, 2011
RESUMEN
En el ejido El Zapote de la Reserva de la Biosfera "Sierra de Huautla", estado de Morelos, México, se seleccionaron doce árboles de Quercus magnoliaefolia Née y se determinó su biomasa en la parte aérea. El modelo alométrico quedó expresado como: B = 0.0345 * DAP29334 en donde B es la biomasa (kg) y DAP es el diámetro a la altura del pecho (cm), con un coeficiente de determinación (R2= 0.98; P<0.001). Adicionalmente, se estimó la biomasa mediante una regresión lineal múltiple que consideró el área basal (AB), la densidad específica de la madera (DEM) y la altura de los árboles (H). Una alta proporción de la varianza de la biomasa fue explicada solamente por el área basal. La DEM y H no incrementaron significativamente la precisión de los modelos. El porcentaje promedio de carbono en los árboles de Q. magnoliaefolia fue de 47.14 con valores de 46.29 % para el fuste, 46.83 % para las ramas y 48.31 % para el follaje. La proporción promedio de los componentes de la especie estudiada fue de 62.5 % para el fuste, 27.8 % para las ramas y 9.6 % para el follaje.
Palabras clave: Ecuaciones alométricas, Quercus magnoliaefolia, biomasa aérea, densidad específica de la madera.
ABSTRACT
At the El Zapote ejido in the "Sierra de Huautla" Biosphere Reserve in the state of Morelos, Mexico, twelve Quercus magnoliaefolia Née trees were selected and their above–ground biomass determined. The proposed allometric model was expressed as: B = 0.0345 * DBH2.9334, where B is biomass (kg) and DBH is diameter at breast height (cm), with a coefficient of determination (R2= 0.98; PO.001). In addition, above–ground biomass was estimated by a multiple linear regression based on basal area (BA), specific gravity of wood (SGW) and tree height (H). A high proportion of the biomass variation was explained by basal area alone. SGW and H did not significantly increase the accuracy of the models. The average percentage of carbon in Q. magnoliaefolia trees was 47.14 with values of 46.29 % for the bole, 46.83 % for branches and 48.31 % for foliage. The average proportion of the components of the species studied was 62.5 % for the bole, 27.8 % for branches and 9.6 % for foliage.
Key words: Allometric equations, Quercus magnoliaefolia, above–ground biomass, wood specific density.
INTRODUCCIÓN
Los bosques de encino históricamente han dominado grandes áreas de las tierras altas de las zonas tropicales y subtropicales de México; estas regiones son unos de los centros más importantes de diversidad del género Quercus en el mundo (Nixon, 1993). Dentro de los bosques de encino en México, el Quercus magnoliaefolia Née corresponde a las especies de distribución amplia media a nivel nacional de acuerdo a la clasificación realizada por Valencia (2004), ya que se localiza en 15 estados de la República mexicana, y como especie de desarrollo amplio puede encontrarse desde 170 a 2,900 m; así, se localiza en la ladera occidental de la Sierra Madre Occidental hasta la Sierra Madre del Sur, desde el sur de Sinaloa, incluyendo partes de Nayarit, Jalisco, Colima, Michoacán, Guerrero y Oaxaca, además de la ladera sur del Eje Neovolcánico en donde se incluyen áreas de algunas entidades ya mencionadas, además de los estados de México, Morelos y Puebla, y llega a presentarse como especie dominante en algunas regiones (Valencia, 2004).
Los datos de la biomasa de los árboles son esenciales para evaluar el secuestro de carbono y su dinámica (Dixon et al., 1994; Binkley et al, 2004), la adaptación de las plantas al ambiente (West–Eberhard, 2003) y también para estudiar el impacto de las prácticas silvícolas en la productividad forestal (Johnson y Todd, 1998; Santa Regina, 2000). Varios factores asociados a la fenología de los árboles y las condiciones del sitio donde se desarrollan pueden influir en la acumulación de biomasa de una determinada especie (Binkley, 2004; Peri et al., 2006; Castilho et al., 2006; Gargaglione et al., 2010), por lo que es necesario diseñar metodologías para cuantificar su biomasa asociada a las condiciones en donde se asientan para establecer las relaciones de las masas forestales con el medio. En este sentido, las ecuaciones alométricas para una especie en particular pueden proporcionar una estimación más acertada de la biomasa y su relación con las condiciones del medio (Litton and Kauffman, 2008).
La mayoría de las ecuaciones alométricas emplean el diámetro a la altura del pecho (DBH) como la única variable independiente y establecen una relación entre esta variable y los componentes de la biomasa del árbol (Gower et al., 1999, Návar, 2009). Algunas estudios proponen incluir la altura de los árboles (H) como la segunda variable predictora y desarrollan ecuaciones con variables combinadas de DBH–H para mejorar la precisión de la estimación de la biomasa (Ketterings et al., 2001; Brandeis et al., 2006; Sampaio et al., 2010), mientras que otras incluyen, adicionalmente a las variables mencionadas, la densidad específica de la madera (DEM) (Martínez–Yrizar et al., 1992; 1996; Gómez et al., 2010).
Los modelos generalizados tienen un gran potencial para realizar inventarios de carbono a gran escala derivados de los datos de los inventarios forestales (Pastor et al., 1984); sin embargo, el conocer los modelos específicos de las especies dominantes en los bosques facilita el estudio para proyectos de investigación ecológicos y de silvicultura, como inventarios detallados de biomasa y carbono almacenado en los árboles, así como para establecer la relación de la acumulación de la biomasa y su relación con las condiciones del medio (Rapp et al., 1999; Wang, 2006).
Con base en lo anterior, los objetivos específicos del presente estudio fueron:
a) ajustar un modelo alométrico para estimar la biomasa aérea total a partir de la variable DAP en Quercus magnoliaefolia Née en el bosque de encino de la Reserva de la Biosfera " Sierra de Huautla", estado de Morelos; b) generar un modelo de regresión lineal múltiple para predecir la biomasa a partir del área basal (AB), densidad específica de la madera (DEM) y la altura (H) de la especie referida para estimar la contribución de estas variables a la estimación de la biomasa aérea total; c) determinar el contenido de carbono de los componentes de la especie estudiada, para de esta manera contribuir a la implementación de métodos de cuantificación de los almacenes de carbono en la biomasa aérea asociados a esta especie forestal.
MATERIALES Y MÉTODOS
El estudio se llevó a cabo en un sitio representativo de la vegetación dominante en la Reserva de la Biosfera Sierra de Huautla, ubicada al sur del estado de Morelos, México, en los terrenos correspondiente al ejido El Zapote (18° 27' 55" a 18° 28' 20" latitud N y 99° 19' 00" a 99° 19' 35" longitud O), en un rango altitudinal de 1,785 a 1,970 m, cuya vegetación característica es de un bosque de encino. El clima prevaleciente en el sitio corresponde a semicálido del grupo de los cálidos subhúmedos (García, 2006), con régimen de lluvias en verano, i.e. marcadamente estacional, con una estación seca que va de enero a principios de mayo, donde se acumula sólo el 4 % de la lluvia anual (1,200 a 1,300 mm) (Gómez, 2008). La temperatura media anual es de 19 a 20 °C, la media más baja ocurre en enero (17 a 18 °C) y la más alta se presenta en abril (22 a 23 °C). Los suelos son principalmente Feozems lúvicos (Hl) con texturas medias a Anas y muy pedregosos en la superficie y a lo largo del perfil; el sustrato es de brechas volcánicas riolíticas interestratificadas con tobas volcánicas de naturaleza ácida. Las topoformas dominantes son sierras de laderas escarpadas con pequeños valles intermontanos altos. Con base en su abundancia relativa en el bosque de encino (Gómez, 2008), se escogió al encino amarillo (Quercus magnoliaefolia Née) para desarrollar los objetivos propuestos.
Generación de las ecuaciones alométricas para estimar la biomasa
Se seleccionaron doce árboles, incluyendo individuos con un rango amplio de dimensiones de acuerdo al tamaño disponible en los sitios donde se realizó el muestreo.
A cada individuo seleccionado se le midió el DAP antes de ser derribado. Los árboles pequeños (2.5 cm < DAP < 10 cm) se cortaron al nivel del suelo. Para este caso, el material cortado y medido se pesó en fresco en campo, y posteriormente se transportó al laboratorio e invernadero. Se secó a peso constante en bancales de invernadero por 30 días para obtener su peso seco. Los árboles más grandes (DAP ≥ 10 cm), tanto el fuste principal como las ramas, fueron cortados en secciones, para obtener submuestras. Se estimó la biomasa de cada árbol siguiendo los criterios propuestos por Acosta et al. (2002). De los individuos con DAP ≥ 10 cm, con el fin de determinar la relación peso seco/peso fresco del tronco y de la copa, a diferentes alturas, y calcular la biomasa total de cada árbol, se extrajeron siete rodajas de aproximadamente 5 cm de espesor en siete posiciones del tallo principal. Las primeras tres rodajas (P1 a P3) se obtuvieron de la sección correspondiente al fuste limpio; P1 a 0.30 m de la superficie del suelo, P2 a 1.3 m de éste (correspondiente a la altura del pecho) y P3 en la parte media entre P2 y la base de la copa. Las otras cuatro rodajas procedían de cuatro secciones en las que se dividió la copa desde su base hasta el ápice. Estas rodajas correspondieron a la base de cada una de esas secciones.
En cada individuo se midió, por separado el peso fresco de las rodajas, de cada sección del tronco y de la copa, incluyendo el follaje, así como de las ramas muestreadas en cada sección. Todas las muestras se transportaron al laboratorio y se sometieron al proceso de secado, como se indicó anteriormente.
La relación peso seco/peso fresco de cada rodaja se utilizó para obtener el peso seco de cada sección del tronco del árbol a partir de su respectivo peso fresco. El mismo procedimiento se utilizó para obtener el peso seco de las ramas (incluyendo el follaje) de cada sección de la copa a partir de la relación peso seco/peso fresco de las rodajas de las ramas muestra. La suma del peso seco de las secciones del tronco y copa, constituyó la biomasa total de cada árbol.
Después de obtener la biomasa y el DAP de cada uno de los árboles, se seleccionó el siguiente modelo, por ser uno de los más frecuentemente usados en este tipo de estudios (Ter–Mikaelian y Korzukhin, 1997; Acosta et al., 2002; Návar, 2009).

donde Y es la biomasa aérea (kg), X es el DAP (cm) y b y k son los parámetros estadísticos estimados. El proceso se realizó con el software estadístico MINITAB 15 (2009).
Para determinar si adicionalmente al DAP existe una correlación estadística entre otras variables y la biomasa, se realizó una regresión lineal múltiple tomando como base los datos transformados de log10 de AB (cm2), DEM (en gcm–3), y h (m) (Martínez–Yrízar et al., 1992; 1996). Los valores de DEM fueron determinados como la relación del peso (g) de las muestras secas de los árboles y el volumen del agua desplazada (cm3) cuando las muestras protegidas por plástico adherente se sumergieron completamente en un recipiente lleno de agua.
La regresión lineal múltiple se realizó con el software estadístico MINITAB 15 (2009), y fue de la forma (Kleinbaum et al., 1988):
donde Y es el log10 de la biomasa (kg); X1 X2 X3 Xi, son el log10 de las variables independientes; β11 es el coeficiente de regresión de Y como función de la variable X1 y βo es la constante de regresión en la intercepción de Y.
Para determinar si la participación de cada variable en el modelo es significativa, se empleó la estrategia de analizar la contribución de cada variable (X1, X2 y X3) estableciendo la relación lineal simple con cada una de las variables. Se probó la hipótesis H0: b1i = 0 mediante el valor de la probabilidad de ese parámetro con un nivel de significancia a= 0.05 (Kleinbaum et al., 1988). Si el valor de probabilidad fue menor de 0.05 se rechazó H0 y se consideró que la variable Xi contribuyó a explicar el modelo.
Determinación del contenido de carbono
De los 12 árboles seleccionados para la determinación de biomasa se eligieron seis para determinar el porcentaje de carbono para cada uno de sus componentes (fuste, ramas y follaje), cubriendo proporcionalmente el rango de diámetros que se muestreó. De las muestras de cada componente arbóreo, una vez secadas a peso constante, se tomó una submuestra de aproximadamente 50 gramos; la concentración de C de cada muestra se determinó por combustión seca con un analizador automático de C total (Shimadzu TOC 5000–A) provisto de una unidad para procesar muestras sólidas (Etchevers et al., 2005). Para estimar el contenido de carbono en la biomasa de los componentes de cada árbol, se multiplicó el valor de la biomasa total del componente (kg) por el porcentaje de carbono del componente determinado en el laboratorio (Díaz et al., 2007). El contenido del carbono de la biomasa total del árbol fue la suma del carbono de cada uno de los componentes de cada árbol.
Adicionalmente se separaron y pesaron en campo los componentes de los mismos seis árboles muestreados, cubriendo un rango de DAP de 10.1 a 31.5 cm, a fin de estimar el porcentaje promedio de biomasa (peso seco) del fuste, ramas y follaje para estimar el contenido ponderado de carbono a partir del porcentaje promedio de biomasa de cada componente y su contenido promedio de carbono.
RESULTADOS Y DISCUSIÓN
Características dasométricas de los árboles muestreados
El DAP de los árboles muestreados varió de 6.5 a 31.5 cm, y su DEM de 0.59 a 0.75 kg·dm–3 con los valores más bajos en los árboles de menor DAP, mismos que se fueron incrementando al aumentar este parámetro; la altura varió de 5.0 a 16.4 m, y su biomasa de 9.0 a 863.7 kg (Cuadro 1).
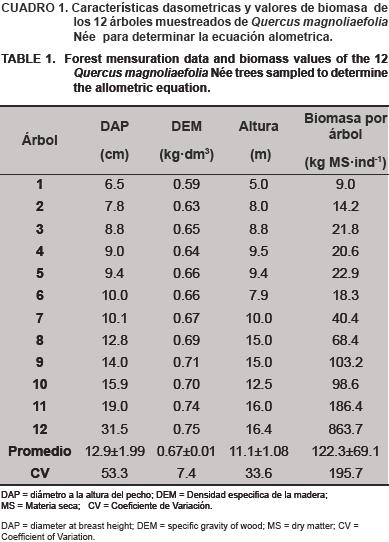
El promedio de la DEM fue de 0.67±0.01 kg·dm–3; el valor de este parámetro es menor al rango reportado por Bárcenas y Dávalos (2001) para los encinos de México, de 0.688 a 0.818 kg·dm–3, sin embargo, este valor de DEM promedio está dentro de los valores registrados por De la Paz y Dávalos (2008) para otros encinos blancos en el país, sección a la que pertenece la especie evaluada, en donde el rango encontrado para ocho especies fue de 0.64 a 0.77 kg·dm–3 y los reportados por Návar (2009) para encinos de Durango donde el valor medio fue de 0.64.
Modelos para la estimación de biomasa
La ecuación para determinar la biomasa del Q. magnoliaefolia Née. quedó expresada como: B =0.0345 *DAP2.9334 en donde B es la biomasa (kg) y DAP es el diámetro a la altura del pecho (cm). La tendencia de los valores de la biomasa aérea con respecto al DAP de los árboles se presenta en la Figura 1.

Modelos de regresión lineal múltiple para la estimación de la biomasa
Las ecuaciones de regresión lineal múltiple para estimar la biomasa se presentan en el Cuadro 2. La biomasa fue explicada solamente por el área basal (AB). La contribución de la densidad especifica de la madera (DEM) y la altura de los árboles (H) no fue significativa una vez que se consideró el AB, como lo demuestran los valores de probabilidad en donde p fue mayor de 0.05. Este resultado difiere de lo encontrado por Martínez–Yrizar et al. (1992, 1996), en donde la ecuación de regresión múltiple, que incluye las mismas variables independientes, predice con mayor confiabilidad la biomasa aérea de un bosque tropical seco en Chamela, Jalisco, que otras relaciones alométricas. Sin embargo, concuerda con lo reportado por Gómez et al. (2010) para los modelos individuales de cinco especies de un bosque tropical seco de la Reserva de la Biosfera Sierra de Huautla, Morelos, en donde sólo fue significativo el aporte del área basal en la estimación de la biomasa.
Estimación del contenido de carbono
De los seis árboles analizados para carbono, el porcentaje promedio para el componente del fuste fue de 46.29±0.27, para las ramas de 46.83±0.22 y en el follaje de 48.31±0.07 (Cuadro 3). El porcentaje promedio para carbono del árbol fue de 47.14±0.17. Estos valores son menores a los reportados para un bosque de encinos en la región Cuicateca de Oaxaca por Figueroa et al. (2005), con 47.3 % de C para el fuste y 49.54 % para las ramas y hojas.
De estos seis árboles, cuyos componentes se pesaron por separado, cubriendo un rango de DAP de 10.1 a 31.5 cm (Cuadro 3), con una biomasa aérea por individuo de 68.4 a 863.7 kg, el porcentaje promedio de biomasa (peso seco) de cada uno de los componentes fue como sigue: el fuste tuvo 62.9±4.3 % con un valor máximo de 74.7 % para uno de los especímenes con menor DAP y el valor mínimo de 45.6 % para el árbol con mayor DAP; para las ramas el porcentaje promedio fue de 27.8±3.2 % con un valor máximo de 40.3 % para la especie con DAP mayor y el valor mínimo de 19.0 % para una de las especies con DAP menor: el porcentaje promedio de la biomasa para el follaje fue de 9.6±1.2 %, con un valor máximo de 14.1 % para el árbol de mayor DAP y el mínimo de 6.28 % para la misma especie referida anteriormente con DAP bajo. En este sentido, Návar (2009) reportó en un estudio para el desarrollo de ecuaciones alométricas de encinos de Durango y Chihuahua una proporción de 68 % de biomasa para el fuste y 32 % para las ramas y el follaje, valores cercanos a los encontrados en este estudio que como se indicó, la proporción varía con el diámetro del árbol.
El contenido de carbono por componente se obtuvo al multiplicar el porcentaje promedio de carbono específico (46.3, 46.8 y 48.3, para fuste, ramas y follaje, respectivamente) por la biomasa medida de dichos componentes (Cuadro 3). El contenido de carbono de cada árbol (CCA ponderado) obtenido a partir de la suma de cada uno de sus componentes y el mismo parámetro (CCA promedio total) pero obtenido a partir de aplicar el factor que representa el porcentaje promedio de carbono para todo el árbol a su biomasa, no presentan diferencias significativas (P = 0.986), por lo que se considera que estos valores son similares, de tal manera que para obtener el contenido de carbono de un árbol no se requiere detallarlo por componentes, resultando más sencillo aplicar la proporción promedio de contenido de carbono a la biomasa del árbol, tal como lo reportan Díaz et al. (2007) para Pinus patula Schl. et Cham.
Dentro de las características de la vegetación, la densidad específica de la madera varía en una misma especie, y su valor está relacionado con las diferencias de crecimiento de sus diámetros y la acumulación de biomasa (Baker et al., 2004; Chave et al., 2001; 2003), además está influida por las condiciones de fertilidad del suelo y el abastecimiento de humedad; en tal sentido, se presentan valores menores en individuos jóvenes de la misma especie y con buen desarrollo comparados con individuos de mayor edad y menor desarrollo asociado a las mayores restricciones en la disponibilidad de nutrientes y agua (Rapp et al., 1999; Wang et al., 2008). Esta variable cambia entre los componentes leñosos, así como con la edad de los árboles (Baker et al., 2004; Návar, 2009).
La ecuación de regresión generada para la especie estudiada predice satisfactoriamente la biomasa a partir del DAP, dado que presentó un coeficiente de determinación (R2) de 0.98 (P< 0.001) y los valores de los parámetros en el ajuste del modelo a forma lineal ln(Y)=b0 + b1 ln(X) fueron los siguientes: la ordenada al origen (b0)= –3.367; la pendiente (b1)= 2.9334, con niveles de significancia aceptables para ambos parámetros (P< 0.001).
En el modelo de regresión lineal múltiple, la poca contribución de la DEM y de la H adicionales al área basal (que es función del DAP) se explica con la poca o nula variación del error estándar y el mínimo incremento del coeficiente de determinación (R2) de la estimación del log10 de la biomasa al incorporar estas variables al modelo.
Si bien en la literatura se reporta un número variado de ecuaciones para la estimación de la biomasa del género Quercus, en la Figura 2 se muestran las líneas de tendencia de este estudio y algunos modelos reportados por diferentes autores: Návar (2009) para encinos de Chihuahua y Durango, Acosta et al., (2002) para un bosque de encino en la zona cuicateca de Oaxaca, y Wang (2006) para un estudio en bosque de Quercus en el noreste de China. En los primeros 20 cm de DAP los cuatro modelos no reportan grandes diferencias, pero para DBH mayores el modelo desarrollado en este estudio y los de Wang (2006) en el noreste de China estiman valores considerablemente mayores de biomasa que los otros dos modelos descritos, lo que puede asociarse a que las condiciones del medio en donde se desarrollan los bosques de este estudio y los del noreste de China tienen condiciones favorables de fertilidad del suelo y de abastecimiento de humedad, por lo cual la acumulación de biomasa es mayor cuando se incrementa el DBH; en cambio los resultados obtenidos por Návar (2009) y Acosta et al. (2002) corresponden a condiciones más restrictivas de suelos y abastecimiento de humedad, tanto en la zona cuicateca de Oaxaca como en las zonas de Durango y Chihuahua, lo que determina menor desarrollo para un determinado DBH de los especímenes de Quercus que allí se desarrollan (Rapp et al., 1999; Wang et al., 2008).
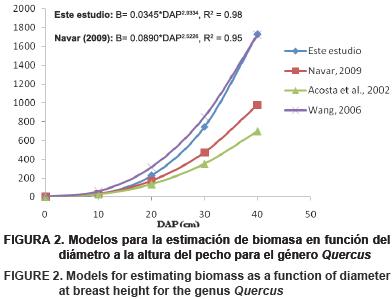
Como se ha indicado en otros trabajos, la variable que mejor explica la biomasa de un árbol es el DBH y es la más comúnmente usada para la alometría de los árboles (Ter–Mikaelian y Korzukhin, 1997; Zianis y Mencuccini, 2004; Návar, 2009). La altura de los árboles es más difícil de medir, y así como la DEM puede no explicar más la varianza de un sitio donde se originaron los datos, pero su incorporación tiene la ventaja de incrementar el potencial de aplicabilidad de las ecuaciones generadas en otros sitios con características similares (Ketterings et al., 2001).
Por otro lado, la variación del contenido de C por componentes en este estudio fue consistente, siendo mayor para el follaje y las ramas, dado que son las estructuras que acumulan altas concentraciones de carbohidratos o compuestos de reserva en contraste con el tronco, en donde la mayoría del C es celulosa y lignina, cuyo contenido de este es menor que en los carbohidratos (Watson y Casper, 1984). Por lo antes señalado, el contenido de C entre los componentes varía también con la edad de los árboles, lo que puede explicar las diferencias entre este estudio y lo reportado por Figueroa et al. (2005).
CONCLUSIONES
El modelo alométrico propuesto para predecir la biomasa de Quercus magnoliaefolia Née, se ajustó con un coeficiente de determinación R2 = 0.98, utilizando la variable diámetro normal.
En la ecuación de regresión lineal múltiple de biomasa estimada a partir de área basal, DEM y altura de los árboles (H), una alta proporción de la varianza de la biomasa fue explicada solamente por el área basal. La DEM y H no incrementaron significativamente la precisión de los modelos individuales, y su contribución para explicar la varianza de la biomasa fue pequeña, al considerar inicialmente el área basal.
El porcentaje promedio de carbono en los árboles de Q. magnoliaefolia fue de 47.14 con valores de 46.29 % para el fuste, 46.83 % para las ramas y 48.31 % para el follaje. La proporción promedio de los componentes de la especie estudiada fue de 62.5 % para el fuste, 27.8 % para las ramas y 9.6 % para el follaje. Los valores del contenido de carbono almacenado por cada árbol, estimado a través de la suma del carbono de sus componentes y el de la resultante del porcentaje promedio de carbono en todo el árbol aplicada a la biomasa de éstos, no presentaron diferencias significativas (P = 0.986).
AGRADECIMIENTOS
Al Consejo Nacional de Ciencia y Tecnología (CONACYT) por el apoyo para la realización de este trabajo mediante la beca para realizar estudios de doctorado del primer autor, al Laboratorio de Fertilidad de Suelos del Colegio de Postgraduados, Montecillos, México, y a la Universidad Autónoma Chapingo, Chapingo, México. Agradecemos el apoyo de las autoridades del ejido El Zapote, Morelos, México, por permitirnos realizar este estudio en sus terrenos, en especial al señor Bulmaro García, la señora Rosenda Montiel y el Señor Miguel Marquina y familia. Apreciamos el apoyo en campo de los señores Pedro López Aguilar, Maximino Baena e Isabel López.
LITERATURA CITADA
ACOSTA M., M.; VARGAS H., J.; VELÁSQUEZ M., A.; ETCHEVERS B, J. D. 2002. Estimación de la biomasa aérea mediante el uso de relaciones alométricas en seis especies arbóreas en Oaxaca, México. Agrociencia 36(6): 725–736. [ Links ]
BAKER, T. R.; PHILLIPS, O. L.; MALHI, Y.; ALMEIDAS, S.; ARROYO, L.; DI FIORI, A.; ERWIN, T.; KELLEN, T. J.; LAURANCE, S. G.; LAURANCE, W. F.; LEWIS, S. I.; LLOYD, J.; MON–TEAGUDOS, A.; NEILL, D. A.; PATIÑO, S.; PITMAN, N. C. A.; NATALINA, J.; SILVA, M.; MARTÍNEZ, R. V. 2004. Variation in wood density determines spatial patterns in Ama–zonian forest biomass. Global Change Biology 10: 545–562. DOI: 10.1111/j.1365–2486.2004.00751.x [ Links ]
BÁRCENAS, G.; DÁVALOS, R. 2001. Shrinking values for 106 mexican Woods. Journal of Tropical Forest Products 7(2): 126–135. [ Links ]
BINKLEY, D. 2004. A hypothesis about the interaction of tree dominance and stand production through stand development. Forest Ecology and Management 190: 265–271. DOI:10.1016/j.foreco.2003.10.018 [ Links ]
BINKLEY, D.; STAPE, J. L.; KYAN, M. G. 2004. Thinking about efficiency of resource use in forest. Forest Ecology and Management 193: 5–16. DOI:10.1016/j.foreco.2004.01.019 [ Links ]
BRANDEIS, T. J.; DELANEY, M.; PARRESOL, B. R.; ROYER, L. 2006. Development of equations for predicting Puerto Rican subtropical dry forest biomass and volume. Forest Ecology and Management 233: 133–142. DOI:10.1016/j.foreco.2006.06.012 [ Links ]
CASTILHO, C. V. d.; MAGNUSSON, W. E.; ARAUJO, R. N. O.d.; LUIZAO, R. C. C.; LUIZAO, F. J.; LIMA, A. P.; HIGUCHI, N. 2006. Variation in aboveground tree live biomass in a central Amazonian forest: effects of soil and topography. Forest Ecology and Management 234: 85–96. DOI:10.1016/j.foreco.2006.06.024 [ Links ]
CHAVE, J.; RIÉRA, B.; DUBOIS, M. A. 2001. Estimation of biomass in a neotropical forest of French Guiana: spatial and temporal variability. Journal of Tropical Ecology 17: 79–96. DOI:do1:10.1017/S0266467401001055 [ Links ]
CHAVE, J., CONDIT, R., LAO, S.; CASPERSEN, J. P.; FOSTER, R. B.; HUBBELL, S. P. 2003. Spatial and temporal variation in biomass of a tropical forest: results from a large census plot in Panama. Journal of Ecology 91: 240–252. DOI: 10.1046/j.1365–2745.2003.00757.x [ Links ]
DE LA PAZ, C.; DÁVALOS, R. 2008. Algunas características anatómicas y tecnológicas de la madera de 24 especies de Quercus (encinos) de México. Madera y Bosques 14(3): 43–80. [ Links ]
DÍAZ F., R.; ACOSTA M., M.; CARRILLO A., F.; BUENDÍA R., E.; FLORES A., E.; ETCHEVERS B., J. D. 2007. Determinación de ecuaciones alométricas para estimar biomasa y carbono en Pinus patula Schl. Et Cham. Madera y Bosques 13(1): 25–34. [ Links ]
DIXON R. K.; BROWN, S.; HOUGHTON, R. A.; SOLOMON, A. M.; TREXLER, M. C.; WISNIEWSKI, J. 1994. Carbon pools and flux of global forest ecosystems. Science, 263: 185–190. DOI:10.1126/science.263.5144.185 [ Links ]
ETCHEVERS, J. D.; MONREAL, C. M.; HIDALGO, C.; ACOSTA, M.; PADILLA, J.; LÓPEZ, R. M. 2005. Manual para la determinación de carbono en la parte aérea y subterránea de sistemas de producción en laderas. Colegio de Postgraduados, México. 29 pp. [ Links ]
FIGUEROA N., C.; ETCHEVERS B., J. D.; VELÁZQUEZ M., A.; ACOSTA M., M. 2005. Concentración de carbono en diferentes tipos de vegetación de la Sierra Norte de Oaxaca. Terra 23(1): 57–64. [ Links ]
GARCÍA, E. 2006. Modificaciones al Sistema de Clasificación Climática de Koppen, 2° ed., UNAM, México, 228 pp. [ Links ]
GARGAGLIONE V.; PERI, P. L.; RUBIO, G. 2010. Allometric relations for biomass partitioning of Nothofagus antárctica trees of different crown classes over a site quality gradient. Forest Ecology and Management 259: 1118–1126. DOI:10.1016/j.foreco.2009.12.025 [ Links ]
GÓMEZ D., J. D. 2008. Determinación de los almacenes de carbono en los compartimentos aéreo y subterráneo de dos tipos de vegetación en la reserva de la biosfera "Sierra de Huautla", Morelos, México. Tesis de Doctor en Ciencias, Colegio de Postgraduados, Montecillos, México. 194 pp. [ Links ]
GÓMEZ D., J. D.; ETCHEVERS B., J. D.; MONTERROSO R., A. I.; PAUSTIAN, K.; HIDALGO, C.; CAMPO A. J.; TINOCO R. J. A.; GUIZAR N., E. 2010. Relaciones alométricas para estimar la biomasa aérea de especies tropicales de bosques estacionalmente secos del Centro de México. En Revisión. [ Links ]
GOWER, S. T.; KUCHARIK, C. J.; NORMAN, J. M. 1999. Direct and indirect estimation of leaf area index, F(APAR), and net primary production of terrestrial ecosystems. Remote Sensing Environment 70: 29–51. [ Links ]
JOHNSON, D. W.; TODD, D. E. 1998. Harvesting effects on long term changes in the nutrient pools of mixed oak fo–rest. Soil Science Soc. Am. J. 62: 1725–1735. DOI:10.2136/sssaj1998.03615995006200060034x [ Links ]
KETTERINGS, Q. M.; COE, R.; VAN NOORDWIJK, M.; AMBAGAU, Y.; PALM, C. A. 2001. Reducing uncertainty in the use of allometric biomass equations for predicting above–ground tree biomass in mixed secondary forests. Forest Ecology and management 146: 199–209. DOI:10.1016/S0378–1127(00)00460–6 [ Links ]
KLEINBAUM, D. G.; KUPPER, L. L.; MULLER, K. E. 1988. Applied regression analysis and other multivariable methods. PWS–Kent Publishinc Co. Boston 718 pp. [ Links ]
LITTON, C. M.; KAUFFMAN, J. B. 2008. Allometric models for predicting above–ground biomass in two widespread woody plants in Hawaii. Biotropica 40: 313–320. DOI: 10.1111/j.1744–7429.2007.00383.x [ Links ]
MARTÍNEZ–YRÍZAR, A.; SARUKHÁN, J.; PÉREZ–JIMÉNEZ, A.; RINCÓN, E.; MAASS, J. M.; SOLIS–MAGALLANES, A.; CERVANTES, L. 1992. Above–ground phytomass of a tropical deciduous forest on the coast of Jalisco, Mexico. J. of Tropical Ecology 8: 87–96. DOI:10.1017/S0266467400006131 [ Links ]
MARTÍNEZ–YRÍZAR, A.; MAASS, J. M.; PÉREZ–JIMÉNEZ, A.; SARUKHÁN, J. 1996. Net primary productivity of a tropical deciduous forest ecosystem in western Mexico. J. Trop. Ecol. 12: 169–175. DOI:10.1017/S026646740000938X [ Links ]
NÁVAR, J. 2009. Allometric equations for tree species and carbon stocks for forest of northwestern Mexico. Forest ecology and Management 257: 427–434. DOI:10.1016/j.foreco.2008.09.028 [ Links ]
NIXON, K. C. 1993. The genus Quercus in Mexico. In: T.P. Ramamoorthy, Bye, R., Lot, A., and Fay, J. (Eds). Biological Diversity of Mexico: Origins and Distribution, pp 447–458, Oxford University Press. New York. [ Links ]
PASTOR, J.; ABER, J. D.; MELILLO, J. M. 1984. Biomass prediction using generalized allometric regressions for some northeast tree species. Forest Ecology and Management 7: 265–274. DOI:10.1016/0378–1127(84)90003–3 [ Links ]
PERI, P. I.; GARGALIONE, V.; MARTÍNEZ–PASTUR, G. 2006. Dynamics of above and below–ground biomass and nutrient accumulation in an age sequence of Nothofagus antarctica forest of Southern Patagonia. Forest Ecology and Management 233: 85–99. DOI:10.1016/j.foreco.2006.06.009 [ Links ]
RAPP, M.; REGINA, I. S.; RICO, M.; GALLEGO, H. A. 1999. Biomass, nutrient content, litterfall and nutrient return to the soil in Mediterranean oak forest. Forest Ecology and Management. 119: 39–49. DOI:10.1016/S0378–1127(98)00508–8 [ Links ]
SAMPAIO, E.; GASSON, P.; BACARAT, A.; CUTLER, D.; PAREYN, F.; COSTA–LIMA, K. 2010. Tree biomass estimation in regenerating areas of tropical dry vegetation in northeast Brazil. Forest Ecology and Management. 259: 1135–1140. DOI:10.1016/j.foreco.2009.12.028 [ Links ]
SANTA REGINA, I. 2000. Biomass estimation and nutrient pools in four Quercus pyrenaica in Sierra de Gata Mountains, Salamanca, Spain. Forest Ecology and management 132: 127–141. DOI:10.1016/S0378–1127(99)00219–4 [ Links ]
TER–MIKAELIAN, M. T.; KORZUKHIN, M. D. 1997. Biomass equation for sixty–five North American tree spe–cies. Forest Ecology and Management. 97: 1–24. DOI:10.1016/S0378–1127(97)00019–4 [ Links ]
VALENCIA A., S. 2004. Diversidad del género Quercus (Fagaceae) en México. Boletín de la Sociedad Botánica de México. 75: 33–53. [ Links ]
WANG, C. 2006. Biomass allometric equations for 10 co–occurring tree species in Chinese temperate forest. Forest Ecology and Management. 222: 9–16. DOI:10.1016/j.foreco.2005.10.074 [ Links ]
WANG, X.; FANG, J.; ZHU, B. 2008. Forest biomass and root–shoot allocation in northeast China. Forest Ecology and Management 255: 4007–4020. DOI:10.1016/j.foreco.2008.03.055 [ Links ]
WATSON, M. A.; CASPER, B. B. 1984. Morphogenetic constraints on pattern of carbon distribution in plants. Annu. Rev. Ecol. Sys. 15: 233–258. DOI: 10.1146/annurev.es.15.110184.001313 [ Links ]
WEST–EBERHARD, M. J. 2003. Developmental Plasticity and Evolution. Oxford University Press. New York. 794 pp [ Links ]
ZIANIS, D.; MENCUCCINI, M. 2004. On simplifying allometric analyses of forest biomass. Forest Ecology and Management 187: 311–332. DOI:10.1016/j.foreco.2003.07.007 [ Links ]

