










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista Chapingo serie ciencias forestales y del ambiente
On-line version ISSN 2007-4018Print version ISSN 2007-3828
Rev. Chapingo ser. cienc. for. ambient vol.16 n.2 Chapingo Jul./Dec. 2010
https://doi.org/10.5154/r.rchscfa.2010.02.003
Estructura y composición de la vegetación del Cañón del Zopilote, Guerrero, México
Structure and composition of vegetation in Zopilote Canyon, Guerrero, Mexico
P. Ávila Sánchez1, 2; A. Sánchez–González1; C. Catalán Everástico2
1 Laboratorio de Sistemática Vegetal, Centro de Investigaciones Biológicas, Universidad Autónoma del Estado de Hidalgo. Ciudad Universitaria, km 4.5 Carretera Pachuca–Tulancingo. Mineral de la Reforma, Hidalgo. C. P. 42184. MÉXICO. Tel. (01)–771 71 7–20–00. Correo–e: pedro940@hotmail.com, arturosg@uaeh.edu.mx.
2 Unidad Académica de Ciencias Agropecuarias y Ambientales, Universidad Autónoma de Guerrero. Periférico Poniente s/n. Colonia Villa de Guadalupe. Iguala, Guerrero. C. P. 40000. MÉXICO. Tel. (01)–733 11 0–15–36.
Recibido: 24 de febrero, 2010
Aceptado: 7 de abril, 2010
RESUMEN
Se analizó el cambio en la composición y estructura de la vegetación a lo largo de un gradiente altitudinal en el Cañón del Zopilote, localizado en la cuenca del río Balsas, en Guerrero. En un intervalo de 450 a 2,800 m de altitud se colocaron 28 parcelas de muestreo, en diferentes tipos de vegetación, definidos por su fisonomía. En cada tipo de vegetación se establecieron seis parcelas de 1,000 m2 para el muestreo de las especies arbóreas, y dentro de ellas una sub–parcela de 100 m2 para arbustivas y dos de 6 m2 para herbáceas. El atributo estructural estimado fue el valor de importancia relativa de las especies; para la clasificación de los tipos de vegetación se utilizó el análisis de agrupamiento. En el gradiente estudiado se identificaron 314 especies de plantas vasculares (cuatro incluidas en la NOM–059–ECOL–2001), en 204 géneros y 79 familias. Las familias mejor representadas fueron Asteraceae, Fabaceae, Burseracea, Lamiacaea y Euphorbiaceae. Se definieron cinco tipos de vegetación: bosque tropical caducifolio, palmar, bosque de Quercus, bosque de Pinusy bosque mesófilo de montaña. Es probable que el cambio abrupto en las condiciones topográficas y climáticas en el Cañón del Zopilote, definan la estrecha proximidad de tipos de vegetación contrastantes.
Palabras clave: Clasificación de la vegetación, bosques montanos, cuenca del río Balsas, gradiente altitudinal, estructura de comunidades.
ABSTRACT
An analysis was carried out of the change in the composition and structure of vegetation along an elevation gradient in Zopilote Canyon, located in the Balsas River Basin in Guerrero. In an altitude range from 450 to 2,800 m, 28 sampling plots were established in five different vegetation types, defined by their physiognomy. Within each vegetation type, six 1,000 m2 plots were selected for sampling tree species, and within these a 100 m2 subplot was established for shrubs and two 6 m2 subplots for herbaceous plants. The structural attribute estimated was the relative importance value of each species; cluster analysis was used for classifying vegetation types. In the elevation range studied, 314 species of vascular plants were identified (including four in the NOM–059–ECOL–2001), in 204 genera and 79 families. The most species–rich families were: Asteraceae, Fabaceae, Burseracea, Lamiacaea and Euphorbiaceae. Five vegetation types were identified: tropical deciduous forest, palm swamp, Quercus forest, Pinus forest and mountain cloud forest. The close proximity of these different vegetation types is most likely due to the abrupt changes in topographical and climatic conditions in Zopilote Canyon.
Keywords: Vegetation classification, mountain forest, Balsas River Basin, elevation gradient, plant community structure.
INTRODUCCIÓN
Se considera que la estructura de la comunidad vegetal es producto de las condiciones físicas locales y de las interacciones entre especies (Ohmann y Spies, 1998), por lo que los cambios ambientales que ocurren a lo largo de gradientes de altitud son considerados determinantes en la composición y estructura de la vegetación (Sánchez–González y López–Mata, 2005; Richter, 2008).
La cuenca del río Balsas es una de las más extensas e importantes de México, cuenta con una gran riqueza florística y tipos de vegetación debido a la variación climática a lo largo del amplio gradiente de altitud existente; presenta matorral xerófilo, bosque tropical caducifolio, bosque de encino, bosque de pino y bosque mesófilo de montaña (Fernández et al., 1998). La cuenca del río Balsas ha sido reconocida como un área fisiográfica con alta concentración de taxones endémicos y como sitio de origen de muchas especies (Rzedowski, 1978, Rzedowski et al., 2005; Peralta, 1995; Fernández et al., 1998; Rodríguez–Jiménez et al., 2005).
El conocimiento de la composición y estructura de la vegetación en las regiones montañosas de México todavía es incipiente (Vázquez y Givnish, 1998). En el Cañón del Zopilote, localizado dentro de la cuenca del río Balsas, existen algunos estudios florísticos y descriptivos de la vegetación (Peralta, 1995; Diego–Pérez et al., 2001; Fonseca et al., 2001; Pineda–García et al., 2007), pero los trabajos cuantitativos sobre la misma, son escasos (Catalán et al., 2003). El objetivo principal del presente estudio fue aportar información sobre los cambios que ocurren en la estructura de la vegetación y en la composición florística a medida que se incrementa la altitud en el Cañón del Zopilote, Guerrero, México.
MATERIALES Y MÉTODOS
Área de estudio. El Cañón del Zopilote tiene una extensión de 738 km2 y coordenadas extremas: 17°39'42", 18°03'59" latitud norte y 99°31'01", 99°46'35" de longitud oeste, se localiza en la cuenca del río Balsas, en el estado de Guerrero (Figura 1). Comprende los municipios de Eduardo Neri, Leonardo Bravo, Tepecoacuilco, Chilpancingo, Tixtla y Cocula (Peralta, 1995). El trabajo de investigación se realizó a lo largo de un gradiente altitudinal que comprende desde 450 m, en el Cañón del Zopilote, hasta los 2,800 m de altitud, en la comunidad de Carrizal de Bravo. La topografía del lugar está formada por cerros y laderas con pendientes pronunciadas y barrancas profundas. Con respecto a la hidrografía, se localiza el río el Zopilote el cual recibe las corrientes de los ríos Coloapán y Huacapán, que descienden de la región sur oriental y fluyen sólo en la época de lluvias (INEGI, 1984).

El clima en la parte baja, en el poblado de Xochipala, es cálido semi–húmedo, con lluvias en verano. En la parte alta, en Carrizal de Bravo el clima corresponde al más húmedo de los templados sub–húmedos, con régimen de lluvias en verano; la precipitación pluvial anual fluctúa de 100 a 1,500 mm y la temperatura media anual varía entre 17 y 21 °C (García, 1973). Los suelos del área en general están clasificados en el grupo de los litosoles, derivados de rocas calizas y lutitas calcáreas (INEGI, 1991). En Carrizal de Bravo los suelos son de origen volcánico, de más de 50 cm de profundidad, textura franco–arcillosa, color pardo rojizo oscuro, con drenaje intermedio y pH de 4.8 a 5.0. Los principales tipos de vegetación en el Cañón del Zopilote incluyen: matorral xerófilo (MX), bosque tropical caducifolio (BTC), bosque de encino (BE), bosque de pino (BP) y bosque mesófilo de montaña (BMM) (Peralta, 1995; Catalán et al., 2003).
Trabajo de campo y de laboratorio. La elección de los sitios de muestreo se hizo de forma preferencial (Matteucci y Colma, 1982) y se basó en el reconocimiento previo de las diferentes variantes fisonómicas de la vegetación presentes en el área de estudio. Los muestreos se realizaron en el sentido del gradiente altitudinal; es decir, desde 450 hasta 2,800 m; a lo largo de este gradiente de 2,350 m se establecieron 28 parcelas de muestreo (Figura 1).
En cada tipo de vegetación se trazaron al azar seis parcelas de muestreo de 1,000 m2 (40 x 25 m) cada una, en las cuales se estimó la densidad y área basal de las especies arbóreas. Dentro de cada parcela, se eligió de forma aleatoria una sub–parcela de 100 m2 (10 x 10 m) y dos sub–parcelas de 6 m2 (2 x 3 m) para contabilizar la densidad y cobertura de las especies del estrato arbustivo y herbáceo, respectivamente.
La estructura de la vegetación se definió con base en el valor de importancia relativa (VIR) de las especies (Mueller–Dumbois y Ellenberg, 1974), con las fórmulas:

para arbustos y herbáceas. La Cobertura (C) viene dada por la fórmula:
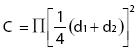
donde π = 3.1416, d1 y d2 son los diámetros mayor y menor perpendiculares entre sí de la proyección vertical de la copa del arbusto o cobertura de las herbáceas. El VIR para árboles se calculó con la fórmula:

El área basal (AB) fue calculada como AB = π r2, donde π = 3.1416 y r es el radio del árbol =  (diámetro a la altura del pecho) de los individuos con DAP ≥ 1 cm.
(diámetro a la altura del pecho) de los individuos con DAP ≥ 1 cm.
Para la clasificación numérica de la vegetación se utilizó el análisis de agrupamiento (CA, por sus siglas en ingles: cluster analysis), con datos del VIR de las especies. Se eligió el índice de S0rensen como la medida de semejanza entre los grupos y como método de unión de grupos el de promedio entre grupos (McCune y Grace, 2002). Para el CA se utilizó el programa PC–ORD (McCune y Mefford, 1999), los resultados se presentan en un dendrograma en el que la medida de distancia se transformó a la función objetiva de Wishart, expresada como porcentaje de información remanente. El porcentaje de información disminuye paulatinamente, hasta ser de cero cuando todos los grupos están fusionados. Para determinar el número de grupos en el dendrograma se eligió un nivel de corte que considerara un compromiso entre la pérdida de información y la simplificación de un número de unidades de vegetación interpretables desde un punto de vista natural (McCune y Grace, 2002).
La mayoría de los ejemplares recolectados fueron determinados con base en las claves para distintas familias, de los fascículos del proyecto de la Facultad de Ciencias de la Universidad Nacional Autónoma de México "Flora de Guerrero", y en algunos géneros se recurrió a taxónomos especialistas de varias instituciones. Los ejemplares recolectados se depositarán en el herbario de la División de Ciencias Forestales de la Universidad Autónoma Chapingo (DICIFO) y en el Herbario Nacional (MEXU).
La nomenclatura utilizada para las familias se basó en Smith et al. (2004), los nombres de las especies conforme a los de la página Web del proyecto Trópicos (2010) y los de las especies de pteridofitas con base en el trabajo de Mickel y Smith (2004). La forma de vida de las especies se estableció en campo, con base en el criterio de Moreno (1984), el tipo de vegetación se definió con base en la clasificación de Rzedowski (1978).
RESULTADOS Y DISCUSIÓN
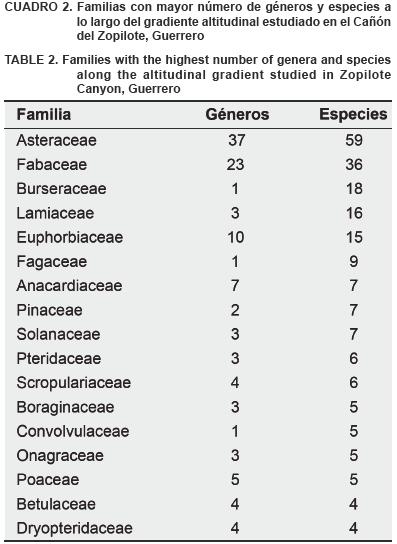
Estudio florístico. Se identificaron 314 especies de plantas vasculares, que corresponden a 79 familias y 204 géneros. (Cuadro 1); las familias con mayor número de especies fueron: Asteraceae (59 especies), Fabaceae (36), Burseraceae (18), Lamiaceae (16), Euphorbiaceae (15) y Fagaceae con 9 especies. Las familias restantes presentaron siete o menos especies (Cuadro 2). Cuatro de las especies se encuentran en alguna categoría de riesgo dentro de la NOM–059–ECOL–2001 (SEMARNAT, 2002): Chiranthodendron pentadactylon Larreat.(amenazada), Abies guatemalensis Rehder. y Zinowiewia concinna Lundell (en peligro de extinción); y Ostrya virginiana (Mill.) K. Koch (sujeta a protección especial).

La cuenca del río Balsas posee una elevada riqueza florística (alrededor de 4,442 especies de plantas vasculares), como resultado de la gran diversidad de condiciones ambientales generadas principalmente por su posición geográfica dentro de dos regiones fisiográficas distintas (Fernández et al., 1998). Ejemplos a nivel local de esta enorme riqueza de especies son los estudios florísticos realizados en zonas aledañas al Cañón del Zopilote, en el BTC y en el bosque de Quercus del municipio de Eduardo Neri (Jiménez et al., 2003) y en la región de Papalutla (Martínez et al., 1997), en donde registran la presencia de 1,342 y de 699 especies de plantas vasculares, respectivamente.
El inventario florístico del presente estudio fue menos rico en especies que los mencionados en el párrafo anterior (Cuadro 3), pues el muestreo se enfocó principalmente a caracterizar las especies que tienen mayor importancia estructural (por su abundancia y cobertura) dentro de los cinco distintos tipos de vegetación identificados. Es por ello que más de 63 % de las 314 especies que conforman la lista, son perennes (141 son especies arbóreas y 59 son arbustivas). Es importante destacar que cuatro de las especies identificadas en el presente estudio, se encuentran en alguna categoría de riesgo en la NOM–059–ECOL–2001 (SEMARNAT, 2002).
Varios estudios realizados en distintos tipos de vegetación dentro de la cuenca del río Balsas, mencionan que las familias de plantas vasculares con mayor riqueza de especies son principalmente: Asteraceae, Fabaceae, Poaceae, Euphorbiaceae, Solanaceae, Burseraceae, Lamiaceae, Orchidaceae, Pteridaceae y Fagaceae (Peralta, 1995; Martínez et al., 1997; Fernández et al., 1998; Zepeda y Velázquez, 1999; Diego–Pérez et al., 2001; Fonseca et al., 2001; Catalán, 2003; Jiménez et al., 2003). Se ha documentado ampliamente que las familias mencionadas son las más representativas (por el número de especies) de la flora mexicana (Rzedowski, 1978, 1996), por lo que no sorprende su elevada riqueza de especies en la zona estudiada, que incluyó cinco tipos diferentes de vegetación.
Comunidades vegetales. Existen pocos estudios cuantitativos sobre la estructura de la vegetación dentro de la cuenca del río Balsas, sólo se puede citar el de Peralta (1995) en el BTC del Cañón del Zopilote y el de Catalán et al. (2003) en el BMM de Carrizal de Bravo. Otros estudios realizados en zonas aledañas incluyen descripciones cualitativas de los tipos de vegetación (Fernández et al., 1998; Diego–Pérez et al., 2001; Fonseca et al., 2001; Lozada et al., 2003).
En el presente estudio, el análisis de agrupamiento permitió distinguir cinco tipos de vegetación en el Cañón del Zopilote (Figura 2), que corresponden a los patrones fisonómicos observados a lo largo del gradiente altitudinal: 1) bosque tropical caducifolio, localizado en el intervalo de altitud de entre 450 y 1,200 m, 2) Palmar entre 1,100 y 1,700 m, 3) bosque de Quercus, distribuido en el intervalo de entre 1,700 y 2,000 m, 4) bosque de Pinus, entre 2,000 y 2,500 m y 5) bosque mesófilo de montaña, entre 2,500 y 3,000 m.

El primer grupo (Figura 2), corresponde al bosque tropical caducifolio (parcelas de muestreo 1 a 6), que se localiza entre 450 y 1,200 m de altitud y se desarrolla principalmente en laderas de cerros con pendientes pronunciadas, que varían de 15 a 29° y con orientación S, E, SO y NO. Los suelos son superficiales, de textura franco a franco arcilloso y el pH alcalino. La precipitación pluvial varía de 450 a 800 mm anuales y la temperatura media anual de 27 a 40° C. Las especies con mayor VIR en el estrato arbóreo fueron: Bursera aptera, Bursera morelensis, Lysiloma tergeminum y Neobuxbaumia mezcalaensis; en el estrato arbustivo sobresalen: Croton sonorae, Hauya elegans, Jatropha andrieuxii, Lippia graveolens y Salvia breviflora. En el estrato herbáceo dominan: Bouteloua disticha, Loeselia glandulosa, Melampodium divaricatum y Oplismenus burmannii (Cuadro 3).
El BTC es el tipo de vegetación más extendido en la cuenca del río Balsas (Fernández et al., 1998). En la zona estudiada este tipo de vegetación se desarrolla dentro del intervalo de altitud referido por varios autores para la cuenca del Balsas (Peralta, 1995, Fernández et al., 1998; Zepeda y Velázquez, 1999). El BTC se caracteriza por poseer una alta riqueza de especies, por lo que su composición y estructura es variable, dependiendo de las condiciones ambientales donde se desarrolle. Peralta (1995) menciona que la heterogeneidad en la composición florística, fisonomía y distribución del BTC en distintas localidades de la cuenca del Balsas está definida por la conjunción de factores físicos, climáticos, ecológicos y humanos. A pesar de la heterogeneidad estructural del BTC, varias de las especies arbóreas que presentaron VIR alto en el presente estudio, destacan también por su constancia y dominancia en otros trabajos realizados en la cuenca del Balsas (Peralta, 1995; Fernández et al., 1998): Acacia angustisima, Bursera morelensis, Bursera xochipalensis, Ceiba aesculifolia y Neobuxbaumia mezcalaensis.
Autores como Zepeda y Velázquez (1999) y Rzedowski et al. (2005), mencionan que el estado de Guerrero es el de mayor riqueza de especies de Bursera, en particular en los BTC de la Cuenca del río Balsas; en la zona estudiada este hecho se manifiesta en la dominancia de tres especies y en la presencia de 18 especies de Bursera en el BTC y en el Palmar. El BTC del Cañón de Zopilote se caracteriza por la dominancia de las formas de vida arbóreas, seguidas de las herbáceas y arbustivas. Zepeda y Velázquez (1999) postulan que la predominancia de especies herbáceas, trepadoras y arbustivas es un indicador de la alteración del BTC. Acorde con este punto de vista, Fernández et al. (1998) mencionan que este tipo de vegetación en condiciones de disturbio, suele dar lugar a un matorral secundario.
El segundo grupo identificado corresponde al Palmar (parcelas 6 al 12), que se localiza en el intervalo altitudinal de 1,320–1,640 m, y se desarrolla principalmente en laderas de cerros con pendientes pronunciadas que varían de 6.4 a 17°, con orientación SE, S, E y NE. Los suelos en este tipo de vegetación son superficiales de textura franco arenosa y el pH varía de ligeramente ácido a ligeramente alcalino. Las especies con mayor VIR en el estrato arbóreo fueron: Acacia riparia, Bursera copalifera, Lysiloma tergeminum y Quercus glaucoides; en el estrato arbustivo dominan Brahea dulcis, Calea ternifolia y Trichospermum grewiifolium. En el estrato herbáceo las especies dominantes fueron Bouteloua disticha, Cosmos sulphureus y Melampodium divaricatum (Cuadro 4).
El Palmar del Cañón del Zopilote forma una franja amplia e irregular, que coincide con la cercanía de varios asentamientos humanos (poblados de Chichihualco, La Laguna y Xochipala), por lo que la mayoría de las parcelas de muestreo presentaron algún indicio de perturbación (presencia de ganado, tala, cultivos, troncos quemados, entre otros). Acorde con los resultados anteriores, García–Mendoza et al. (1994) refieren que en la mixteca alta Oaxaca–Puebla, el Palmar secundario de Brahea dulcis se encuentra en áreas que están sometidas a quemas periódicas y su crecimiento se ve favorecido por la tala del bosque de encino.
Illsley et al. (2001), mencionan que en la región de Chilapa, Guerrero, los palmares de Brahea dulcis crecen en sitios donde originalmente existían bosques de encinos (Quercus glaucoides y otras especies de encinos caducifolios), en asociación con especies de la familia Fabaceae y Burseraceae. Los mismos autores refieren que la densidad de manchones de palma aumenta en las localidades donde el disturbio es evidente y creciente (pastoreo de ganado e incendios), hasta que las demás especies son casi eliminadas del sitio. Los hallazgos de García–Mendoza et al. (1994) y de Illsley et al. (2001), permiten suponer que el Palmar presente en el Cañón del Zopilote se desarrolla en una zona de transición natural entre el BTC y el bosque de Quercus, en sitios donde estos bosques han cedido su lugar a una vegetación secundaria. Un vestigio más de este proceso de recambio es el elevado VIR dentro del Palmar de especies típicas del BTC (Bursera copalifera), del bosque de Quercus (Quercus glaucoides) y de vegetación secundaria (Brahea dulcis e Ipomoea arborescens).
El siguiente grupo, constituido por las parcelas de muestreo 13 a 18 (Figura 2), comprende el bosque de encino, un tipo de vegetación bien definido que se desarrolla entre los 1,915 y 1,966 m de altitud; principalmente en laderas de cerros con pendientes pronunciadas que varían de 13 a 26.5°, con orientación E, NE, E y N. Los suelos son superficiales, de textura franco arenosa a franco–arcillosa y el pH es ligeramente alcalino. Las especies con mayor VIR en el estrato arbóreo fueron: Quercus laeta, Q. obtusata y Q. resinosa; en el estrato arbustivo: Brahea dulcis, Brickellia veronicifolia, Dalea bicolor y Lippia graveolens; en el estrato herbáceo sobresalen Rhynchosia pringlei, Salvia keerlii y Stevia ovata (Cuadro 4).
Fernández et al. (1998) hacen una descripción general de los bosques de Quercus de la cuenca del río Balsas, que concuerda con la del presente estudio, en especial en la composición de especies, aunque el intervalo altitudinal de distribución del bosque es muy amplio (entre 900 y 2,000 m), comparado con lo registrado en el Cañón del Zopilote. Los encinares subtropicales forman asociaciones transicionales entre los bosques templados y húmedos de las serranías y el BTC de la cuenca del Balsas (Fernández et al., 1998; Fonseca et al., 2001). En la zona de estudio se observó una zona de transición similar en donde los bosques de Quercus son sustituidos gradualmente por bosques templados sub–húmedos de Pinus, que se desarrollan en el intervalo altitudinal de entre 2,145 y 2,490 m.
El cuarto grupo ilustrado en la Figura 2, corresponde al bosque de pino (parcelas de muestreo 19–24), que se desarrolla en altitudes de entre 2,145 y 2,490 m; se localiza principalmente en laderas de cerros con pendientes pronunciadas que varían de 13.5 a 38.5° y con orientación E, NE, S y NO. Los suelos son moderadamente profundos, de textura franco a franco arcilloso y de pH ácido. Las especies con mayor VIR en el estrato arbóreo fueron Arbutus xalapensis, Pinus devoniana, Pinus lawsonii y Pinus oocarpa; en el estrato arbustivo dominan Agave cupreata, Chromolaena odorata, Pteridium arachnoideum y Solanum rudepannum. En el estrato herbáceo sobresalen Gnaphalium roseum, Lamourouxia multifida, Russelia floribunda y Vernonia patens (Cuadro 4).
A diferencia de otros bosques de Pinus de la cuenca del Balsas, en donde dominan Pinus montezumae, P. leiophylla o P. pseudostrobus (Fernández et al., 1998), en los bosques del Cañón del Zopilote las especies de pinos con mayor VIR fueron Pinus oocarpa, P. devoniana y P. lawsonii. En la zona estudiada el bosque de Pinus es reemplazado gradualmente por el BMM.
El siguiente grupo (Figura 2) corresponde al bosque mesófilo de montaña (parcelas 24 a 28), presente en altitudes de 2,620 a 2,784 m y se localiza principalmente en laderas de cerros con pendientes pronunciadas que varían de 9 a 29°, en sitios con orientación E y NE. Los suelos son profundos de textura franco arenosa, ricos en materia orgánica y de pH ácido. La precipitación pluvial anual varía de 1,100 a 1,500 mm y la temperatura media anual de 17 a 21 °C (García, 1973). Las especies con mayor VIR en el estrato arbóreo fueron Abies guatemalensis, Buddleja cordata, Critonia paneroi y Persea americana; en el estrato arbustivo fueron: Cestrum nitidum, Roldana reticulata, Rumfordia floribunda y Solanum nigricans; y en el estrato herbáceo dominan las especies: Dryopteris wallichiana, Prunella vulgaris, Salvia carnea y Salvia langlassei (Cuadro 4).
De acuerdo con Rzedowski (1978, 1996), el BMM es un tipo de vegetación complejo y heterogéneo, que incluye bosques bajos, de mediana estatura y muy altos, tanto perennifolios como caducifolios. En términos de su composición florística el BMM no presenta uniformidad y las especies dominantes varían ampliamente de un lugar a otro, dependiendo de las diferentes condiciones geográficas y ecológicas en las que se distribuyen. En la cuenca del Balsas la distribución del BMM es limitada y fragmentaría, debido a las condiciones templado–húmedas que requiere (Fernández et al., 1998).
La composición y estructura de distintas variantes de los BMM del Cañón del Zopilote y zonas aledañas han sido descritas por Fonseca et al. (2001), Diego–Pérez et al. (2001) y Catalán et al. (2003). Los tres autores coinciden en la descripción de una variante de BMM muy semejante a la encontrada en el presente estudio, en cuanto a la dominancia en el estrato arbóreo de Abies guatemalensis, Chiranthodendron pentadactylon, Cleyera integrifolia, Critonia paneroi, Oreopanax xalapensis, Persea americana y Pinus pseudostrobus. De acuerdo con Catalán et al. (2003), el BMM del Cañón del Zopilote es uno de los más diversos en especies leñosas en México y es único por la dominancia de Chiranthodendron pentadactylon en el estrato arbóreo, especie considerada en peligro de extinción (SEMARNAT, 2002).
En el Cañón del Zopilote existen tipos de vegetación con características contrastantes (Figura 3): el BTC y el Palmar son típicos de la zona tropical seca de México; el BE y BP son característicos de la zona templada sub–húmeda y el BMM que pertenece a la zona templada húmeda (sensu Toledo y Ordoñez, 1998). La topografía compleja de muchas montañas tropicales, que presentan terrenos extremadamente accidentados, con pendientes de exposición diferente, ha generado una extraordinaria variedad de hábitats húmedos y secos en estrecha proximidad (Fernández et al., 1998; Richter, 2008). En sitios alejados entre sí tan sólo unos cuantos kilómetros de distancia, la cantidad de lluvia puede diferir en varios órdenes de magnitud (Diego–Pérez et al., 2001), de aquí que una intrincada orografía y por ende complejidad micro–climática, pueda ser la explicación de la estrecha proximidad de tipos de vegetación característicos de ambientes contrastantes (secos, sub–húmedos y húmedos) dentro del Cañón del Zopilote.
CONCLUSIONES
El análisis de agrupamiento permitió reconocer cinco diferentes tipos de vegetación, dentro del Cañón del Zopilote, en un amplio intervalo altitudinal de 2,350 m. El cambio en la composición florística y en la estructura de cada tipo de vegetación fue más evidente en los extremos del gradiente altitudinal analizado. En pocos kilómetros de distancia la topografía y el clima en la zona estudiada cambian abruptamente y, por ende, las especies dominantes (las que definen la estructura de la comunidad) son completamente diferentes. El Cañón del Zopilote es un área importante desde el punto de vista de la conservación de la flora, en el presente estudio se identificaron cuatro especies que están incluidas en alguna categoría de riesgo dentro de la NOM–059–ECOL–2001.
AGRADECIMIENTOS
El primer autor agradece a la Universidad Autónoma de Guerrero, su apoyo al otorgar el permiso para realizar estudios de doctorado y al CONACYT por concederle una beca económica (número 16303) para dichos estudios. También se agradece al M. en C. Enrique Guízar Nolasco, encargado del herbario de la Universidad Autónoma Chapingo, por su apoyo en la identificación de algunas especies.
LITERATURA CITADA
CATALÁN H., C.; LÓPEZ–MATA, L.; TERRAZAS T. 2003. Estructura, composición florística y diversidad de especies leñosas en un bosque mesófilo de montaña de Guerrero, México. Anales del Instituto de Biología, Universidad Nacional Autónoma de México, Serie Botánica 74: 209–230. [ Links ]
DIEGO–PÉREZ N.; PERALTA–GÓMEZ S.; LUDLOW–WIECHERS B. 2001. El Jilguero. Bosque Mesófilo de Montaña. In: DIEGO–PÉREZ N. y R. M. FONSECA (eds.). Estudios florísticos en Guerrero. Número 11. Las Prensas de Ciencias. Facultad de Ciencias, Universidad Nacional Autónoma de México. México, D.F. 42 p. [ Links ]
FERNÁNDEZ N., R.; RODRÍGUEZ J., C.; ARREGUÍN S., M. L.; RODRÍGUEZ J., A. 1998. Listado florístico de la cuenca del río Balsas, México. Polibotánica 9: 1–151. [ Links ]
FONSECA R. M.; VELÁZQUEZ E.; DOMÍNGUEZ E. 2001. Carrizal de Bravos. Bosque Mesófilo de Montaña. In: DIEGO–PÉREZ N y R. M. FONSECA (eds.). Estudios florísticos en Guerrero. Número 12. Las Prensas de Ciencias. Facultad de Ciencias, Universidad Nacional Autónoma de México. México, D.F. 41 p. [ Links ]
GARCÍA E. 1973. Modificaciones al sistema de clasificación climática de Koppen. Offset Larios, México, D.F. [ Links ]
GARCÍA–MENDOZA A.; TENORIO L., P.; REYES S., J. 1994. El endemismo en la flora fanerogámica de la mixteca alta, Oaxaca–Puebla, México. Acta Botánica Mexicana 27:53–73. [ Links ]
ILLSLEY G., C.; AGUILAR J.; ACOSTA G., J.; GARCÍA B., J.; GÓMEZ A., T.; CABALLERO N., J. 2001. Contribuciones al conocimiento y manejo campesino de los palmares de Brahea dulcis (HBK) Mart. en la región de Chilapa, Guerrero. In: RENDÓN A. B., S. REBOLLAR D., J. CABALLERO N., M. MARTÍNEZ A. (eds.). Plantas, Cultura y Sociedad. Primera edición. Universidad Autónoma Metropolitana. México, D.F. pp. 259–286. [ Links ]
INEGI, 1984. Carta topográfica 1:50,000 Xochipala E14C18. Instituto Nacional de Estadística Geografia e Informática. [ Links ]
INEGI, 1991. Carta Geológica 1:250,000. Chilpancingo E14–8. Instituto Nacional de Geografía e Informática. [ Links ]
JÍMENEZ R., J.; MARTÍNEZ G., M.; VALENCIA A., S.; CRUZ D., R.; CONTRERAS J., J.L.; MORENO G., E.; CALÓNICO S., J. 2003. Estudio florístico del municipio de Eduardo Neri, Guerrero. Anales del Instituto de Biología, Serie Botánica 74(1): 79–142. [ Links ]
LOZADA L.; LEÓN M.E.; ROJAS J.; DE SANTIAGO R. 2003. Bosque mesófilo de montaña en el Molote. In: DIEGO–PÉREZ N. y R. M. FONSECA (eds.). Estudios florísticos en Guerrero. Número 13. Las Prensas de Ciencias. Facultad de Ciencias, Universidad Nacional Autónoma de México. México, D.F. 35 p. [ Links ]
MARTÍNEZ G., M.; VALENCIA A., S.; CALÓNICO S., J. 1997. Flora de Papalutla, Guerrero y de sus alrededores. Anales del Instituto de Biología, Serie Botánica 68(2): 107–133. [ Links ]
MATTEUCCI S., D.; COLMA A. 1982. Metodología para el estudio de la vegetación. Serie Biología. Monografía No. 22. Secretaría General de la Organización de los Estados Americanos. Programa Nacional de Desarrollo Científico y Tecnológico. Washington, D.C. 168 pp. [ Links ]
MCCUNE B.; GRACE J. B. 2002. Analysis of ecological communities. MjM Software Designe. Gleneden Beach, Oregon. 300 pp. [ Links ]
MCCUNE B.; MEFFORD M. J. 1999. PC–ORD. Multivariate Analysis of Ecological Data. Version 4 for Windows. MjM Software Design. Gleneden Beach, Oregon. 237 pp. [ Links ]
MICKEL J. T.; SMITH A.R. 2004. The Pteridophytes of Mexico. Memoirs of the New York Botanical Garden. New York Botanical Garden Press, Nueva York, Volumen 88. 1054 pp. [ Links ]
MORENO N. P. 1984. Glosario botánico ilustrado. Continental. México, D.F. 300 pp. [ Links ]
MUELLER–DOMBOIS D.; ELLENBERG H. 1974. Aims and methods of vegetation ecology. John Wiley. New York, New York. 547 pp. [ Links ]
OHMANN L. J.; SPIES A. T. 1998. Regional gradient analysis and spatial pattern of woody plant communities of Oregon forest. Ecological Monographs 68: 151–182. [ Links ]
PERALTA G., S. 1995. Cañón del Zopilote (Área Papalotepec). In: DIEGO–PÉREZ N. y R. M. FONSECA (eds.). Estudios Florísticos en Guerrero. Las Prensas de Ciencias. Facultad de Ciencias, UNAM. México, D.F. 37 pp. [ Links ]
PINEDA–GARCÍA F.; ARREDONDO–AMEZCUA L.; IBARRA–MANRÍQUEZ G. 2007. Riqueza y diversidad de especies leñosas del bosque tropical caducifolio El Tarimo, Cuenca del Balsas, Guerrero. Revista Mexicana de Biodiversidad. 78: 129–139. [ Links ]
RODRÍGUEZ–JÍMENEZ C.; FERNÁNDEZ NAVA R.; ARREGUÍN–SÁNCHEZ M. L.; RODRÍGUEZ–JÍMENEZ A. 2005. Plantas vasculares endémicas de la cuenca del río Balsas, México. Polibotánica 20: 73–99. [ Links ]
RZEDOWSKI J. 1978. La vegetación de México. Limusa. México. 432 pp. [ Links ]
RZEDOWSKI J. 1996. Análisis preliminar de la flora vascular de los bosques mesófilos de montaña de México. Acta Botánica Mexicana 35: 25–44. [ Links ]
RZEDOWSKI J; MEDINA L., R.; CALDERON DE RZEDOWSKI G. 2005. Inventario de conocimiento taxonómico, así como de la diversidad y del endemismo regionales de las especies mexicanas de Bursera (Burseraceae). Acta Botánica Mexicana 70: 85–111. [ Links ]
RICHTER M. 2008. Tropical Mountain forests – distribution and general features. In: GRADSTEIN S. R., HOMEIER J. y GANSERT D. (eds.). The tropical Mountain Forest – Patterns and Processes in a Biodiversity Hotspot. Gottingen Centre for Biodiversity and Ecology. The Biodiversity and Ecology Series 2: 7–24. [ Links ]
SÁNCHEZ–GONZÁLEZ A.; LÓPEZ–MATA L. 2005. Plant species richness and diversity along an altitudinal gradient in the Sierra Nevada, Mexico. Diversity and Distribution 11: 567–575. [ Links ]
SEMARNAT [Secretaría del Medio Ambiente y Recursos Naturales]. 2002. Norma Oficial Mexicana NOM–059–ECOL–2001, Protección ambiental–Especies nativas de México de flora y fauna silvestres–Categorías de riesgo y especificaciones para su inclusión, exclusión o cambio–Lista de especies en riesgo. Diario Oficial de la Federación. 2a Sección, 6 de marzo de 2002. [ Links ]
SMITH N.; MORI S. A.; HENDERSON A.; STEVENSON D. W.; HEALD S. V. 2004. Flowering plants of the Neotropics. The New York Botanical Garden. Princeton University Press. USA. 594 pp. [ Links ]
TOLEDO V. M.; ORDÓÑEZ M. J. 1998. El panorama de la biodiversidad de México: Una revisión de los hábitats terrestres. En: RAMAMOORTHY T.P., R. BYE, A. LOT y J. FA (eds.). Diversidad Biológica de México: orígenes y distribución. Instituto de Biología, Universidad Nacional Autónoma de México. México, D.F. pp. 739–757. [ Links ]
TROPICOS. 2010. Tropicos.org. Missouri Botanical Garden. <http://www.tropicos.org>. © 2010 Missouri Botanical Garden – 4344 Shaw Boulevard – Saint Louis, Missouri 63110. [ Links ]
VÁZQUEZ G., J. A.; GIVNISH T. J. 1998. Altitudinal gradient in tropical forest composition, structure, and diversity in the Sierra de Manantlán. Journal of Ecology 86: 999–1020. [ Links ]
ZEPEDA G., C.; VELAZQUEZ M., E. 1999. El bosque tropical caducifolio de la vertiente sur de la Sierra de Nanchititla, Estado de México: La composición y la afinidad geográfica de su flora. Acta Botánica Mexicana 46: 29–55. [ Links ]

