










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista Chapingo serie ciencias forestales y del ambiente
On-line version ISSN 2007-4018Print version ISSN 2007-3828
Rev. Chapingo ser. cienc. for. ambient vol.14 n.1 Chapingo Jan./Jun. 2008
Relaciones tróficas de los peces del embalse San Miguel Arco, de Soyaniquilpan, Estado de México
Relations trofics of the fish of the San Miguel Arco dam, in Soyaniquilpan, State of Mexico.
M. Á. Yubi Armendáriz; N. A. Navarrete Salgado; G. Elías Fernández, G. Vázquez Gómez; E. S. Urrieta Zapiain
Laboratorio de producción de peces e invertebrados. UNAM–FES–IZTACALA. Avenida de los Barrios Núm. 1. Tlanepantla, Estado de México.
Recibido: 27 de agosto, 2007
Aceptado: 11 de septiembre, 2007
RESUMEN
Las relaciones de los organismos en la naturaleza se basan en dos ciclos, uno de materia y el otro de energía, dichos ciclos dan origen a las cadenas tróficas las cuales dan origen a los tres niveles tróficos; autótrofos, herbívoros y carnívoros. Para el caso de los peces de agua dulce, los organismos jóvenes viven y se alimentan en la orilla y superficie del agua, para conocer lo que comen se analiza el tracto digestivo. Por lo cual el presente trabajo pretende determinar la trama trófica de los peces del embalse San Miguel Arco. Se capturaron los peces del embalse con un chinchorro de 30 metros de largo y 1/3 de pulgada, los organismos se fijaron con formalina al 10 %, los peces se identificaron al igual que el alimento que consumieron, con claves especializadas, a la abundancia de los grupos alimenticios se les aplicó el índice de Simpson. Se encontró que Chirostoma humboldtianum, es zooplanctófago especialista, Cyprinus carpió, y Carassius auratus son planctófagos generalistas y Poeciliopsis infans es planctófago especialista. La trama trófica se basa en zooplancton (Bosmina y Mastigodiaptomus) además de zoobentos (Corixidae).
Palabras clave: trama trófica, alimentación, índice de Simpson.
ABSTRACT
The relations of the organisms in the nature are based on two cycles, one of matter and the other of energy, these cycles give origin to the food chains which give origin to the three food levels; autotrophs, herbivorous and carnivores. For the case of freshwater fishes, the young organisms live and they are fed in the border and surface of the water, to know what they eat analyzes alimentary canal. Thus the present work tries to determine the trofhic plot of the fish of the San Miguel dam Arc. The fish of the dam with a small boat of 30 meters in length captured themselves and 1/3 of inch, the organisms paid attention with formalin to 10 %, the fish were identified like the food that consumed, with specialized keys, to the abundance of the nutritional groups I am applied the index to them of Simpson. One was that Chirostoma humboldtianum, is zooplanctofago specialistic, Cyprinus carpio, and Carassius auratus is planctofagos generalist and Poeciliopsis infans is planctófago specialistic. The trofic plot is based on zooplancton (Bosmina and Mastigodiaptomus) in addition to zoobentos (Corixidae).
Keywords: it plots trofhic, feeding, index of Simpson.
INTRODUCCIÓN
En los sistemas naturales, se presentan dos ciclos, uno de materia y otro de energía. El primero es cerrado e implica el consumo de materia orgánica viva y el segundo consiste en la degradación de la materia orgánica muerta, que pasa a formar parte de un organismo vivo (Ricklefs y Miller, 2000). Estas relaciones mantienen el material y el flujo energético dentro del ecosistema y proporcionan lo necesario para la subsistencia del mismo. Estas relaciones originan las cadenas tróficas, las cuales están compuestas por nodos, cada uno de los cuales representa una especie o grupos de especies que son tróficamente similares, esto es, que tienen similares presas y similares depredadores, por lo que las distintas especies están dispuestas en un orden determinado en el ecosistema (Ricklefs y Miller, 2000). Así se establecen tres niveles tróficos: autótrofos, herbívoros y carnívoros (Odum y Sarmiento, 1998). Dicho modelo se refiere a un hábitat simple y no considera los cambios a través del tiempo. Las tramas tróficas reales son sistemas multidimensionales dinámicos (Moore y Ruiter, 1991). Las tramas donde hay pocos bloques o especies tróficas pueden tener un mayor número de niveles que aquellas en las que se consideran muchas especies tróficas (Martínez, 1993).
La respuesta de los organismos ante diferencias en la cantidad de alimento depende de las características de cada especie, así como la estructuración de la trama trófica en la que se encuentren incluidos (Stevens y Steiner, 2006). Para peces de sistemas epicontinentales, los jóvenes habitualmente viven junto a la orilla o en la superficie del agua, donde se alimentan de organismos planctónicos microscópicos (Muus y Dahlstrõm, 1970). Cuando el número de taxas en una comunidad disminuye, el número proporcional de conexiones dentro de la trama trófica también pueden disminuir, lo cual simplifica la estructura de la cadena trófica (Parker y Huryn, 2006).
El alimento consumido se convierte en energía, el exceso, es utilizado para crecer, la magnitud del crecimiento depende del valor nutritivo del alimento y de la cantidad ingerida (Muus y Dahlström, 1970). Gracias a los estudios del contenido del tracto digestivo, se ha podido comprender muchas de las relaciones de las especies y sus hábitos alimenticios, muchos de los cuales son seleccionados por su tamaño y no por su clase (Hobson, 1991).
En México existen muchos cuerpos de agua, en muchos casos las especies que se encuentran son endémicas, por lo que es de gran interés realizar estudios detallados de estos organismos, las condiciones específicas del medio que influye en los ciclos de vida, reproducción y hábitos alimenticios y las relaciones tróficas de cada especie.
De los estudios referentes a la alimentación de peces podemos destacar a: quien estudió los hábitos alimenticios de los peces del Embalse Santa Helena en el Estado de México; realizaron un estudio similar en el Embalse Trinidad Fabela; Cohen, etal., (1990) examinaron 113 tramas tróficas de comunidades marinas, epicontinentales y terrestres. Biró, en 1995 estudió el manejo de estanques y las cadenas tróficas. Drossel, et al., en 2004 realizaron un trabajo sobre el impacto en la estructura a largo plazo de las tramas tróficas ante respuestas funcionales no lineales. Pereira y Espíndola en 2004 estudiaron la alimentación de los peces del Embalse Três Irmãos en Brasil. Por todo lo anterior el presente estudio tiene como objetivo determinar la trama trófica de los peces del embalse San Miguel Arco y compararlo con trabajos similares en la misma zona de estudio.
MATERIALES Y MÉTODOS
El estudio se realizó en el Embalse "San Miguel Arco" ubicado entre los 20° 37' latitud norte y los 99° 32' longitud oeste, en el municipio de Soyaniquilpan de Juárez, Estado de México (Figura 1), a una altitud de 2,460 m. El municipio de Soyaniquilpan se encuentra dentro del Eje Neovolcánico Transversal, en la región hidrológica "Alto Pánuco" (INEGI, 2002). El clima del lugar es el más húmedo de los templados subhúmedos con lluvias en verano, la temperatura oscila entre los 6.5 a 22 °C, con una precipitación anual de 749.5 mm (García, 1988).

El estudio se realizó el día 5 de mayo del 2006, estableciéndose cinco estaciones de muestreo a lo largo del Embalse "San Miguel Arco". En cada estación se registraron parámetros físicos y químicos como temperatura del agua por medio de un termómetro digital "Elite"; la transparencia del agua se obtuvo con un disco de Secchi. El pH del agua con un potenciómetro digital Cole Palmer. El oxigeno disuelto, la alcalinidad y la dureza del agua se determinaron con pruebas titulométricas (APHA, AWWA Y WPCF, 1992). La conductividad se midió con un conductímetro Sprite 6000 y la turbidez del agua con un turbidímetro La Motte 2002.
Los peces se colectaron con un chinchorro playero de 30 m de largo, 3 m de caída y 1/3 de pulgadas de abertura de malla; los organismos obtenidos se fijaron en formalina al 10 % (APHA, AWWA Y WPCF, 1992).
Los peces se identificaron con las claves de Álvarez (1970) y Barbour (1973), se contaron y posteriormente se pesaron en una balanza semianalítica ACCULAB modelo 333 y se midieron con un vernier. Se extrajo el tracto digestivo para determinar su contenido estomacal por medio de las claves de Pennak (1989) para zooplancton y Ortega (1984) para fitoplancton; con los valores numéricos obtenidos, se calculó el índice de Simpson (Krebs, 1994).
RESULTADOS
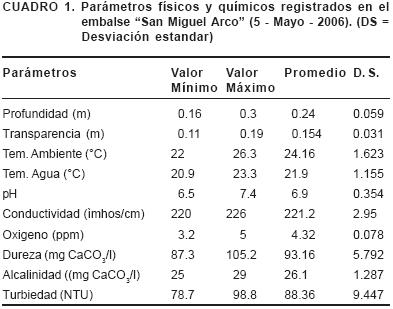
Los valores de los parámetros ambientales registrados se muestran en el Cuadro 1.
En el embalse se encontraron cuatro especies de peces, las cuales fueron Cyprinus carpio, Chirostoma humboldtianum, Poeciliopsis infans y Carassius auratus.
C. humboldtianum consumió 25 grupos de presas entre copépodos, cladóceros, rotíferos, microalgas y pastos, en cuanto al porcentaje numérico de los grupos ingeridos, las más representativas fueron Bosmina, los huevos de Copépodo y Mastigodiaptomus con 77.14, 7.27 y 4.76 % respectivamente (Figura 2a), de los cuales las más representativas en cuanto al porcentaje volumétrico fueron Bosmina con 88.56 %, seguido de Mastigodiaptomus y Daphnia con 7.48 y 1.16 % respectivamente (Figura 2b). Además, se obtuvo un valor de 0.37 del Índice de Simpson.

C. carpio consumió un total de 29 grupos distribuidos entre copépodos, cladóceros, rotíferos, microalgas, pastos, coríxidos y quironómidos. Dentro de estos grupos, los porcentajes más representativos en cuanto al número de individuos ingeridos fueron Mastigodiaptomus, Bosmina y Cyclops con 36.53, 12.26 y 11.26 % respectivamente (Figura 3a). En cuanto al porcentaje volumétrico Mastigodiaptomus, Bosmina y los pastos de los grupos Setaria y Bromus, fueron las más importantes con 53.98, 14.75 y 11.04 % respectivamente (Figura 3b). Para C. Carpio el valor arrojado por el Índice de Simpson fue de 0.82.

Para C. auratus se tuvieron cuatro grupos de organismos ingeridos, los cuales fueron Mastigodiaptomus, Bosmina, Daphnia, y Cyclops, de los cuales los tres primeros fueron los más representativos con respecto al porcentaje e individuos ingeridos, con 65.31,14.28,11.22 y 9.18 % respectivamente (Figura 4a). En cuanto al porcentaje volumétrico, los grupos fueron Mastigodiaptomus con 79.84 %, Bosmina con12.59 % y Daphnia con un 5.73 % (Figura 4b). El valor del Índice de diversidad de Simpson fue de 0.54.

P. infans ingirió un total de ocho grupos dentro de los cuales se encuentran microalgas, cladóceros, copépodos y coríxidos. Los grupos con mayor porcentaje numérico de organismos ingeridos fueron Microcystis, Haematococcus y Synedra, con 42.36, 28.02 y 17.07 % respectivamente (Figura 5a), en cuanto al porcentaje volumétrico los grupos mejor representadas fueron, Corixidae con 67.9 % y Bosmina con 31.10 % (Figura 5b). El Índice de Simpson tuvo un valor de 0.7.

En la trama trófica se observó que la dieta de C. carpio fue a base de Bosmina y Mastigodiaptomus. C. auratus tenían como contenido estomacal Bosmina, Mastigodiaptomus y Daphnia, P. infans consumió principalmente Coríxidos. Mientras tanto para C. humboldtianum se encontró que Bosmina y Mastigodiaptomus fueron los principales grupos consumidos (Figura 6).

DISCUSIÓN
El embalse San Miguel Arco es un sistema lótico donde son descargados residuos de una planta cercana productora de queso, el agua del embalse es clara de acuerdo a los criterios mencionados por Navarrete et al. (2004) y tiene una profundidad promedio de 0.24 m, presenta ligeras deficiencias de O2. Por su dureza el agua es ligeramente dura según la clasificación de la U. S. Geological Survey (Navarrete, 2004) mientras que la clasificación de Jiménez et al. (citado por Navarrete et al. (2004), indica que el agua es moderadamente dura; por su alcalinidad se clasifican como agua blanda, con lo que se esperaría una producción de peces y plantas de media a baja. La alcalinidad y la dureza son factores que pueden alterar el pH, que para este sistema correspondió en promedio a 6.9 ubicándola como agua ligeramente ácida, por último, la conductividad del agua lo ubica como agua dulce dura.
Dentro del embalse, C. humboldtianum se desarrolló como un organismo zooplantófago, que basó su dieta en Bosmina y Mastigodiaptomus, lo cual concuerda con los trabajos de Orbe–Mendoza et al. (2002), Blancas et al. (2003) y Navarrete y Cházaro (1992); y como especialista según los criterios de Navarrete et al. (2007), que con base en el Índice de diversidad de Simpson, nos dice, que se alimenta de zooplancton, principalmente de los géneros Daphnia sp., Bosmina sp., Ceriodaphnia sp. y de un copépodo diaptómido en estado juvenil que no fue posible identificar.
C. carpio basa su alimentación en el de plancton consumiendo en mayor volumen Mastigodiaptomus, además de Bosminas, algas y pastos, caracterizando a esta especie como omnívora, lo que concuerda con el trabajo de Blancas et al. (2003) y de acuerdo con el Índice de Simpson encontramos que es muy generalista, de acuerdo a los criterios de Navarrete et al. (2007).
C. auratus basó su alimentación en organismos planctónicos donde la especie mas consumida fue Mastigodiaptomus junto con Bosmina, como lo reporta Navarrete, et al. (2006), que nos dice, que con relación a los ítems alimentarios encontrados en C. auratus durante la época primaveral, se alimentaron de Bosmina, larvas de Chironomidae, Daphnia y Keratella, con esto podemos decir que C. auratus es una especie planctófaga, y de acuerdo con el Índice de diversidad de Simpson es considerada generalista con base en los criterios de Navarrete et al. (2006), quienes reportan que la dieta de C. auratus varia a lo largo del año. Pero los mismos autores, nos dicen que los factores físicos y químicos registrados en la temporada de primavera, son los mejores factores para el adecuado desarrollo del pez.
De acuerdo al porcentaje volumétrico se determinó que P. infans es un organismo planctófago–bentófago debido a que en el tracto digestivo se encontraron Coríxidos y Bosminas, y otros organismos encontrados en menor proporción, siendo los primeros los más importantes, caracterizando a P. infans como una especie planctófaga–bentófaga generalista, de acuerdo a los criterios de Navarrete et al. (2007).
Un aspecto importante es que en la estructura de la trama trófica de mayo de 2006 no aparecen dos depredadores que estaban presentes en la estructura anterior que son: Ctenopharyngodon idella y Girardinichthys multiradiatus (Navarrete et al., 2006). Esto no indica que las especies estén ausentes en el cuerpo de agua pero sí es posible que las poblaciones de éstas hayan reducido drásticamente a lo largo del tiempo entre muestreos. En un cuerpo de agua en el que existen dos presas y dos depredadores, un especialista y un generalista es probable que si ocurre una baja en la densidad de población de las presas, los organismos más afectados sean los especialistas ya que el generalista se vería menos afectado al poder optar por el consumo de otras presas (Anderson et al., 2006).
Todas estas variaciones en la trama trófica también se ven influenciadas por las características físicas y químicas del agua del sistema como el oxigeno el cual es un factor importante ya que si se reduce su concentración, también puede reducir la actividad fotosintética. En el embalse se encontró que el agua era deficiente en oxigeno por lo que se puede suponer que la producción primaria estaba siendo afectada por este factor, reduciendo por consecuencia la cantidad de energía que fluye en la cadena trófica; otro factor importante es el pH, el cual depende, en parte, de la concentración de oxigeno y su relación con la respiración y la fotosíntesis, estos procesos puede causar el aumento progresivo de materia orgánica en el medio el cual provoca la explosión poblacional de algunas especies zooplanctónicas y propiciar la eutrofización del sistema (Legendre y Rivkin, 2002).
El fitoplancton y el zooplancton son muy sensibles a las variaciones del pH, si éste cambia pueden poner en estrés a las poblaciones zooplanctónicas, al reducir el fitoplancton, esto a su vez también reduciría la disponibilidad de presas para los peces que consumen zooplancton obligándolos a diversificar su dieta si es que estos son generalistas o a reducir su población si es que estos son más especialistas, según la cadena trófica de abril de 2005 C. idella y G. multiradiatus se encontraban como especies especialistas ya que consumían pastos y Quironómidos respectivamente, al haber cambios en las poblaciones de zooplancton probablemente las especies menos especialistas ganaron recursos que éstas necesitaban y así se pudiese explicar la ausencia de Ctenopharyngodon idella y Girardinichthys multiradiatus en la cadena trófica de especies encontradas para mayo de 2006.
La trama trófica cambió hacia zooplancton y zoobentos, dejando a un lado a los pastos (Bromus y Setaria) que eran abundantes en 2005 (Navarrete et al., 2007) y dejaron de serlo en 2006.
CONCLUSIONES
C. humboldtianum se desarrolló como un organismo zooplanctónico y especialista. Mientras C. carpio se manifestó como una especie plantófaga y generalista.
C. auratus se reportó como planctófaga además de generalista. En tanto que P. infans fue reportada como una especie especialista, planctófaga–bentónica.
En la trama trófica desaparecen G. Multiradiatus por la caída del zooplancton y C. idella por la ausencia de pastos.
La trama trófica se enfoca hacia el consumo de zooplancton (Bosmina y Mastigodiaptomus) y zoobentos (Corixidae).
LITERATURA CITADA
ÁLVAREZ, V. J. 1970. Peces Mexicanos (Claves). Serie de Investigación Pesquera. Estudio I. Instituto Nacional de Investigaciones Biológico Pesqueras. México. 166 p. [ Links ]
ANDERSON, A.; SAMUELSSON, K.; HAECKY P.;ALBERTSSON, J. 2006. Changes in the pelagic microbial food web due to artificial eutrophication. Aquat. Ecol. 40: 299–313. [ Links ]
APHA, AWWA & WPCF. 1992. Métodos normalizados para el análisis de aguas potables y residuales. Díaz de Santos, S. A. Madrid. España. 1134 pp. [ Links ]
BARBOUR, C. D. 1973. The Systematics and evolution of the genus Chirostoma Swainson (Pisces, Atherinidae). Tulane Studies in Zoology and Botany. 18(3):97–141. [ Links ]
BLANCAS, A. G.; FIGUEROA, L. G.; BARRIGA, S. I. A.; ARREDONDO, F.J. L. 2003. Aportaciones al cultivo del pez blanco, Chirostoma humboldtianum. (Pisces: Atherinidae). pp 155168. En: P. M. ROJAS C. y D. FUENTES C. (Comps.). Historia y avances del cultivo de pescado blanco. Instituto Nacional de la Pesca. Dirección General de Investigación en Acuacultura. [ Links ]
BIRÓ, P. 1995. Management of pond ecosystems and trophic webs. Aquaculture. 129: 373–386. [ Links ]
COHEN, J. E.; BRIAND, F.; NEWMAN, C. M. 1990. Community food webs: Data and theory. 554–555 pp. En: RICKLEFS, R. E.; G. L. MILLER, 2000. Ecology. 4ª ed. W. H. Freeman and Company. New York. 822 pp. [ Links ]
DROSSEL, B.,A.; MCKAEN, J.; QUINCE, C. 2004. The impact of nonlinear functional response on the long–term evolution of food web structure. Journal of Theoretical Biology. 229: 539–548. [ Links ]
GARCÍA, E. 1988. Modificación al Sistema de Clasificación Climática de Köppen (para adaptarlo a las condiciones de la República Mexicana). Instituto de Geografía. Universidad Nacional Autónoma de México. México, D. F. 220 p. [ Links ]
HOBSON, E. S. 1991. Trophic Relationships of Fishes Specialized to Feed on Zooplankters above Coral Reefs. 69–95 pp. En: SALE, P. F. (ed). The Ecology of Fishes on Coral Reefs. Academic Press. London. 754 pp. [ Links ]
INEGI. 2002. Instituto Nacional de Estadística Geografía e Informática. Carta Hidrológica, F–14C88. Escala 1:50 000. México, D. F. [ Links ]
KREBS, J. Ch. 1994. Ecological Methodology, Addison–Weley Educational Publishers Inc. U. S. A. 440–444 p. [ Links ]
LEGENDRE, L.; RIVKIN, R. B. 2002. Pelagic food webs: responses to enviromental processes and effects on the environment. Ecologycal Research. 17: 143–149. [ Links ]
MARTÍNEZ, N. D. 1993. Effects of resolution on food web structure. Oikos. 66: 412–413. [ Links ]
MOORE, J. C.; DE RUITER, P. C. 1991. Temporal and spatial heterogeneity of trophic interactions within bellow–grownd food webs. Agruculture, Ecosistems & Environment. 34: 371–397. [ Links ]
MUUS, B. J.; DAHLSTRÖM, P. 1970. Los peces de agua dulce de España y de Europa. Omega. Barcelona. 232 pp. [ Links ]
NAVARRETE, S. N.; CHÁZARO, O. S. 1992. Espectro trófico del charal Chirostoma humboldtianum Valenciennes (Atherinidae) del Embalse Tiacaque, México. Revista de Zoología. ENEP Iztacala UNAM. 3: 28–34. [ Links ]
NAVARRETE S. N.; ELÍAS, F. G.; CONTRERAS, R. G.; ROJAS, B. M. L.; SÁNCHEZ, M. R. 2004. Piscicultura y ecología en estanques dulceacuícolas. AGT. 180 pp. [ Links ]
NAVARRETE, S. N.; SORIANO, A. E.; CONTRERAS, R. G.; DUARTE, S. M. A.; SÁNCHEZ, M. R.; GUZMÁN, T. D. 2006. Alimentación de la carpa dorada Carassius auratus (Pisces: Cyprinidae) en el Embalse San Miguel Arco, Estado de México. Revista de Zoología. 17: 9–17. [ Links ]
NAVARRETE S. N.; AGUILAR, R. J.; GONZÁLEZ, D. J. M.; ELÍAS, F. G. 2007. Espectro trófico y trama trófica de la Ictiofauna del Embalse San Miguel Arco, Soyaniquilpan, Estado de México. Revista de Zoología. 18: 12–22. [ Links ]
ODUM, E. P.; SARMIENTO, F. O. 1998. Ecología. El puente entre ciencia y sociedad. McGraw–Hill Interamericana, México. 343 pp. [ Links ]
ORBE–MENDOZA, A. A.; ACEVEDO–GARCÍA. J.; LYONS, J. 2002. Lake Pátzcuaro fishery management plan. Reviews in Fish Biology and Fisheries. 12: 207–217. [ Links ]
ORTEGA, M. M. 1984. Catálogo de algas continentales recientes de México. Universidad Nacional Autónoma de México. Dirección General de Publicaciones. México. 566 p. [ Links ]
PARKER, S. M.; HURYN, A. D. 2006. Food web structure and function in two arctic streams with contrasting disturbance regimes. Freshwater biology. 51, 1249–1263. [ Links ]
PENNAK, R. W. 1989. Fresh–Water invertebrates of the United States. John Wiley and Sons. Inc. New York. 803 p. [ Links ]
PEREIRA, S.; ESPÍNDOLA, E. 2004. Hábitos alimenticios de nueve especies de peces del Emblase de Três Irmãos, São Paulo, Brasil. Universidad y Ciencia. 1: 33–38. [ Links ]
RICKLEFS, R. E.; MILLER, G. L. 2000. Ecology. 4a ed. W.H. FREEMAN AND COMPANY. New York. 822 pp. [ Links ]
STEVENS, H. H.; STEINER, C. E. 2006. Effects of predation and nutrient enrichment on a food web with edible and inedible prey. Freshwater Biology. 51, 666–671. [ Links ]

