











 text new page (beta)
text new page (beta) English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkIntroduction
Throughout the world, large tracts of original natural habitat have been replaced by mosaics of diverse habitats with different levels of human intervention, including forest fragments, areas with secondary vegetation, croplands and induced grasslands for cattle (Mena 2010). In many of these landscapes, there are continuous, structurally contrasting features with respect to adjacent habitats, which connect with each other and with different vegetation remnants (Gurrutxaga and Lozano 2008; García and Abad 2014). These types of linear features, present in heterogeneous landscapes, can be natural, such as rivers, canyons, riparian zones and vegetation strips (Limpens and Kapteyn 1991; Van Geert et al. 2010), or artificial, such as roads, railways and live fences (Van der Zanden et al. 2013).
For many non-flying land mammals, linear components function as corridors that facilitate movement between patches (Bennet 1998; Gurrutxaga and Lozano 2008; Huber et al. 2012). Similarly, some bats use linear landscape features as flight paths and foraging areas (Brandt et al. 2007; Loram et al. 2011). The linear features most used by bats are hedgerows, rivers and live fences, as well as other landscape components that have continuity such as forest edges and stone walls (Downs and Racey 2006; García and Santos 2014). In the case of insectivorous species, it has been suggested that linear features could be widely used because they offer greater food availability, protection against wind and predators, as well as spatial references that facilitate navigation (Limpens and Kapteyn 1991; Toffoli 2016). In some cases, the use of these spatial components may decrease the energetic cost of searching for foraging sites (Kusch et al. 2004; Barragán et al. 2010).
The structural characteristics of linear features could be related to the frequency with which they are used by different bat species. For example, it has been observed that the most commonly used live fences are those with the highest heterogeneity levels, height and shrub cover (Fuentes-Montemayor et al. 2013). On the other hand, the intensity of use of these landscape components appears to be related to the flight patterns and echolocation of the different bat species. In particular, it has been observed that insectivorous species that emit intense, low frequency echolocation pulses (dominated by constant frequency components) tend to commute in open areas, while insectivorous species that produce weak, high frequency pulses (dominated by modulated frequency components) are those that use the landscape’s linear features with greater intensity (Limpens and Kapteyn 1991; Bouhey et al. 2011).
The common vampire bat, Desmodus rotundus (Chiroptera, Phyllostomidae), is relatively abundant in cattle-ranching landscapes in Latin America (Escobar et al. 2015). This species, which feeds mainly on the blood of large herbivorous mammals, leaves its roost every night to search for prey during the periods of greatest darkness (Wimsatt 1969). Unlike insectivorous species, D. rotundus may depend on its vision to orient itself in space (Greenhall et al. 1983; Eklof 2003). Because its visual field is limited by its tendency to fly very close to the ground, D. rotundus is likely to rely heavily on spatial references to orient itself in the landscape. In addition, its limited ability to detect small or distant objects through echolocation (Joermann 1984; Rodriguez and Allendes 2016) would force it to fly close to objects that serve as a reference. According to this hypothesis, D. rotundus could be expected to fly very close to linear features in cattle-ranching landscapes dominated by open areas. To test this prediction, we evaluated the use of the different linear features of the landscape by D. rotundus in an agroecosystem of the municipality of Tacotalpa, in the state of Tabasco, Mexico. The objectives of the present study were: 1) to estimate the flight height and flight distance relative to linear landscape features by D. rotundus; and 2) to compare the use that D. rotundus makes of live fences, forest edges and small streams as flight routes. The results of this work allow us to assess the importance of linear landscape features for the spatial navigation of D. rotundus, as well as to identify sites where cattle could be more vulnerable to attacks by this predator.
Materials and methods
Study area. This study was conducted in the vicinity of the Xicoténcatl rural settlement, in the municipality of Tacotalpa, Tabasco. The region’s characteristic climate is warm and humid, with year-round rainfall, average monthly temperatures ranging from 22 to 28 °C, and average annual rainfall of between 2,000 and 4,000 mm (INEGI 1998). In the low mountainous elevations, vegetation consists of fragments of medium-height semi-evergreen tropical forest and tall evergreen tropical forest with different levels of disturbance, while in the low, slightly undulating plains, induced grasslands predominate (Martínez et al. 2013). Large tracts of grassland are crossed by numerous live fences and some rivers, shallow valleys and streams. As in the entire municipality of Tacotalpa, the main economic activity is extensive cattle ranching (Secretaría de la Reforma Agraria 2012).
The main linear features present in the study area are live fences, streams, and forest edges. In this work, we consider forest edges as linear features, based on the classification of landscape components proposed by Downs and Racey (2006). In the study area, live fences represent the boundaries of the pastures, and are made up of small trees, known locally as cocoíte (Gliricidia sepium), linked with barbed wire. Initially, the cocoítes are planted as small trunks, but later branch again to form live trees arranged in a line. The region’s streams are made up of shallow, conspicuous depressions that form in unstable terrain. Although the presence of flowing water is seasonal, they generally maintain small accumulations of water or mud in which hydrophilic grasses and forbs can grow. Some of these streams form small, relatively deep valleys bordered by steep slopes. Forest fragments are generally restricted to the steepest hills, so their edge is often delineated by the base of slopes and some stone walls. Due to deforestation, some forest edges have scattered trees, so the boundaries may be blurred.
Sampling design. In total, we selected 24 sampling sites, spaced at least 150 m, at forest edges (n = 9), live fences (n = 10) and streams (n = 5). The sites were selected in such a way that they were not in the immediate vicinity (at least 60 m around) of other linear features that might interfere with the evaluated site. The first sampling stage was carried out between February 28 and April 29, 2014. A second set of data was collected between February 2016 and June 2017, at 24 sites different from those of the first sampling period. In this second stage, 12 sites were located in the El Paraíso ranch and 12 in the neighboring Poaná ejido; of the total number of sites, 19 corresponded to live fences, 4 to forest edges and 1 to a stream. The data from the second stage are derived from a parallel study carried out by our research group, whose objective depended on the collection of D. rotundus data throughout an annual cycle.
Data collection. Considering that D. rotundus reduces its activity during periods of greatest moonlight intensity (Flores-Crespo et al. 1972), we avoided sampling in the 10 days of most intense moonlight. We monitored each site for two non-consecutive nights per visit (one visit in 2014; six visits in 2016 to 2017). At each site, we placed a mist net perpendicular to the axis of the linear feature, positioning the first supporting pole as close as possible (< 30 cm) to it. During the 2014 sampling, we placed 24 continuous m of net (combinations of 6 and 12 m nets aligned and adjacent, leaving no gaps between them), so the total sampling area covered from the vicinity of the linear feature to the interior of the open areas. We did not attempt to extend the nets beyond 24 m for two reasons: to avoid approaching adjacent linear features, especially in small pastures, and to reduce damage to nets caused by cattle. In addition, available information suggests that D. rotundus is rarely captured in open pastures (e. g., Medina et al. 2007). Because the sampling effort increased significantly (spatially and temporally) during the second stage, only 12 m of nets were placed at each site in 2016 to 2017. In both stages, nets were placed at dusk and left open for seven hours (stage 2014) or five hours (stage 2016 to 2017). During the first stage, we made a total sampling effort of 8,064 m-net per hour, of which 3,024 m-net per hour was carried out at forest edges, 3,360 m-net per hour at live fences and 1,680 m-net per hour at streams. During the second stage, a total sampling effort of 25,920 m-net per hour was made.
We used a flexometer to measure the height from the ground and distance from the linear feature of each individual found in the net. We identified the captured animals according to the criteria suggested by Medellín et al. (2008). To avoid counting newly released specimens twice, we fitted each individual with a plastic necklace containing small beads representing a color code. After obtaining routine morphometric data, the individuals were released at the same capture point.
Data analysis. Chi-square goodness of fit tests were used to assess whether D. rotundus tends to fly more frequently at a particular distance from linear landscape features, separately for each type (live fences, forest edges and streams) and together. For this, the number of captures was grouped according to the distance to the linear feature in 2 m intervals and compared with the number of captures that would be expected if the individuals were flying homogeneously in the landscape (equal capture frequency in each distance interval). This analysis was performed separately for the 2014 and 2016 to 2017 samples. To determine whether D. rotundus uses all three types of linear features with the same frequency, a non-parametric analysis of variance (Kruskal-Wallis) was performed using the number of captures per site as the unit of analysis (Durán et al. 2005). Since only during the first sampling could we include an adequate number of sites corresponding to each type of linear feature, this last analysis was restricted to the 2014 sample.
Results
During the samplings carried out in 2014, we obtained a total of 50 captures corresponding to the species D. rotundus. Of the total number of individuals, 24 were caught at live fences, 20 at forest edges and six at streams. Only one individual was recaptured once. From the samplings conducted in 2016 and 2017, we obtained a total of 58 data on flight height and distance from the linear features. Results from both periods show that D. rotundus tends to fly very close to the ground (average height: 77 cm in 2014 and 69 cm in 2016 to 2017). Also, 88 % of the 2014 captures and 82 % of the 2016 to 2017 ones were obtained at heights equal to or less than 100 cm. However, no individual was captured more than 160 cm above ground level.
We observed that D. rotundus shows a tendency to fly very close to the linear features of the landscape and to avoid open areas; 52 % of the 2014 catches and 62 % of the 2016 to 2017 ones were obtained in the first two meters, with the number of captures gradually decreasing as the distance from the linear feature increased (Figure 1). According to the Chi-square test, the use of the airspace covered by the nets adjacent to the linear features was not homogeneous (2014: X² = 132.59, P < 0.001; 2016 to 2017: X² = 92.57, P < 0.001).
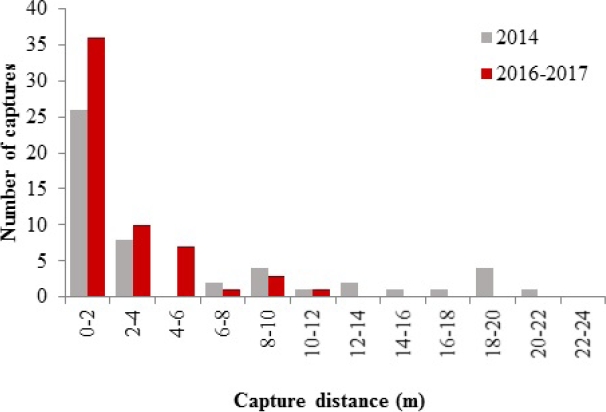
Figure 1 Number of D. rotundus individuals captured as a function of distance from linear landscape features (forest edges, live fences and streams) in cattle-ranching landscapes of Tacotalpa, Tabasco, for sampling periods 2014 and 2016-2017.
According to the Kruskal-Wallis test performed with the 2014 data, D. rotundus does not preferentially use one particular type of linear feature (H = 2.15, P = 0.342), regardless of the capture distance. The average number of captures per site was 2.2 individuals for forest edges, 2.4 for live fences and 1.2 for streams. However, the flight pattern was different for each type of linear feature (Figure 2). At the forest edges, (X² = 51.90, P < 0.001), 80 % of the captures were made in the first four meters, with the number of captures notably decreasing in the other distances. A similar pattern was observed in the streams and live fences (streams: X² = 8.62, P = 0.03; live fences: X² = 28.40, P < 0.001), although the percentage of captures in the first 4 m was lower than that of forest edges (Figure 2). Comparing the flight pattern in the three types of linear features, it can be observed that the tendency to fly near the linear features was more pronounced at the forest edges.
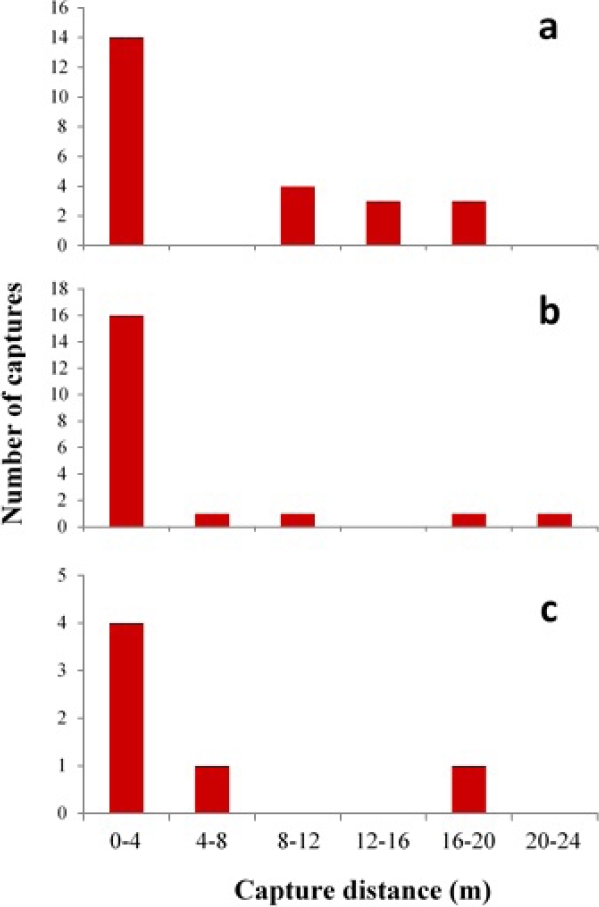
Discussion
Despite the extensive knowledge accumulated over several decades about the ecology of D. rotundus, this is the first study that describes its commuting patterns in agricultural landscapes. The flight height recorded in this study for D. rotundus is similar to that reported in a previous study on the attack behavior of this species, in which it was observed that individuals approached their prey at a height of between 50 and 150 cm (Greenhall et al. 1971). In the present study, the average flight height was around 70 cm, indicating that vampires tend to fly at low altitudes while searching for prey or while commuting to and from their roosts.
It was observed that D. rotundus flies preferentially near linear landscape features and avoids open areas (grasslands with scattered trees), as has been reported for several species of insectivorous bats (Lumsden and Bennett 2005; Downs and Racey 2006; Hein et al. 2009). Although there was no statistically significant preference for any type of linear feature, the greatest number of captures occurred at live fences and forest edges. The low level of stream use is similar to that observed in insectivorous species (Brandt et al. 2007), which could be because these sites increase the risk of predation due to the lack of tree cover (Russ and Montgomery 2002). However, this hypothesis has little support considering that D. rotundus avoids commuting during nights or hours with high levels of moonlight as a strategy to decrease the risk of predation (Flores-Crespo et al. 1972). Alternatively, lack of tree cover is likely to reduce the effectiveness of streams as spatial reference features in predominantly open landscapes.
Heavy rains and strong winds have been reported to decrease the foraging activity of D. rotundus (Wimsatt, 1969). Therefore, flying close to linear features may offer the common vampire bat protection against extreme environmental conditions, as has been suggested for some insectivorous species (Limpens and Kapteyn 1991; Brandt et al. 2007). This may explain, in part, why this species was observed more frequently using those linear features that had some degree of tree cover (live fences and forest edges).
Although D. rotundus has a moderately developed visual capacity (Greenhall et al. 1983; Eklof 2003), its low flight height may limit its field of vision. In addition, the short detection range by echolocation would prevent it from detecting objects at great distances, which in turn would make commuting and orientation difficult in open areas (Joermann 1984). For this reason, we suggest that D. rotundus shows a greater tendency to fly near linear landscape features because they act as reference points for spatial orientation (for a discussion on the role of linear features in insectivorous species, see Limpens and Kapteyn 1991).
One factor that may influence the usefulness of linear landscape components as reference points for the spatial orientation of D. rotundus is the level of contrast between the linear feature and the open areas. If these features are well defined in space, they could facilitate the commuting of individuals along the landscape (Fuentes-Montemayor et al. 2013). This would explain the low level of use observed in streams, which had a more diffuse structure relative to the surrounding pastures, while forest edges and living fences were generally well defined linearly.
It has been suggested that food abundance is a factor promoting the use of linear features by insectivorous bats (Verboom and Huitema 1997; Lumsden and Bennett 2005; Fuentes-Montemayor et al. 2013; Kalda 2015). This theory, however, has not been explored in non-insectivorous species, including D. rotundus. In a study conducted in a cattle-ranching area of Colombia, it was found that, during the hottest time of the year, cattle explore areas with tree cover in search of a better food supply during the night and part of the early morning (Polania 2012). Therefore, there is a possibility that, at least in some regions and at some times of the year, D. rotundus would prefer to commute in the immediate vicinity of wooded areas because they have greater prey availability.
The flight patterns observed in this study suggest that the most vulnerable prey to hematophagous bat attacks would be those resting at night near linear landscape features. This would explain why domestic animals maintained near wooded areas and D. rotundus roosting sites are most often bitten by vampire bats (Novaes et al. 2010; Costa and Esbérard 2011; Hernández-Pérez et al. 2019). In addition, this hypothesis is supported by the results of various experimental studies which documented that D. rotundus has a short detection range of objects (including experimental “prey”) through the senses of echolocation (Joermann 1984), smell (Bahlman et al. 2007), thermoperception (Kürten et al. 1982), and passive hearing of sounds emitted by prey (Schmidt et al. 1991; Heffner et al. 2013). The role of sight is more uncertain (Greenhall et al. 1983; Eklof 2003), although the visual detection of prey by D. rotundus can be expected to decrease with increasing distance from its flight paths. An additional reason that might explain the preference of D. rotundus for linear landscape features (especially wooded ones) is the search for levels of darkness that maximize its ability to visually detect prey, while reducing the likelihood of being detected by them. The three hypotheses (need for spatial references, maximization of visual function in conditions of maximum darkness and minimization of the risk of being detected by prey) are not mutually exclusive and could occur simultaneously.
The flight patterns reported in this study for the common vampire bat are consistent in space and time. Considering that the number of recorded individuals decreases gradually as the distance from the linear features of the landscape increases, we can suggest (with a high level of confidence) that D. rotundus prefers to fly close to the linear features in predominantly open landscapes. The results reported herein have strong implications for livestock management in tropical regions, considering that the production animals most vulnerable to attacks by D. rotundus (and therefore most at risk of being infected with rabies) are those that rest near linear landscape features.

