











 text new page (beta)
text new page (beta) Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkIntroducción
Los venados temazates rojo (Mazama temama) y café (M. pandora), son dos de las cinco especies de cérvidos que habitan en México (Gallina y Mandujano 2009). Ambas especies se distribuyen en las áreas tropicales del país y en general la información sobre ellas es escasa (Mandujano 2011). La UICN (Unión Internacional para la Conservación de la Naturaleza) considera al M. temama como especie con datos insuficientes (Bello et al. 2008), mientras que el M. pandora lo incluye en la categoría vulnerable (Weber et al. 2008). En México ninguna de las dos especies se incluye en alguna categoría de riesgo (SEMARNAT 2010a), pero a partir de 2014 se les considera como especies prioritarias de conservación (SEMARNAT 2014). En México el aprovechamiento de ambas especies es posible dentro del esquema de cacería de subsistencia y con restricciones dentro del esquema de Unidad de Manejo para la Conservación de la Vida Silvestre (UMA; Weber y González 2003).
La importancia principal de los temazates en México se debe a que forman parte de la dieta de los pobladores rurales, razón por la cual son intensamente cazados dentro del esquema de cacería de subsistencia (Naranjo y Bodmer 2007; Reyna-Hurtado y Tanner 2010; Santos-Fita et al. 2012; Weber 2014). Asimismo, las especies de Mazama mexicanos son valorados como recurso cinegético (García-Marmolejo et al. 2008), ya que son trofeos de caza mayor y forman parte de competencias internacionales de cacería deportiva (Villarreal et al. 2008).
Las características de hábitat que determinan la presencia/ausencia de temazates son dependientes de la escala a la que se midan (García-Marmolejo et al. 2015). Esto obedece a que un mismo proceso ecológico puede generar patrones diferentes a distintas escalas espaciales al estar regulado por mecanismos distintos en cada escala (Levin 1992). La escala de paisaje o macrohábitat debe entenderse como aquella que permite evaluar la respuesta de los organismos frente a condiciones y recursos ambientales que son heterogéneos en el espacio, condicionando en gran medida el funcionamiento de los organismos a dicha heterogeneidad espacial (Fortin y Dale 2005).
La presencia de M. temama se relaciona con la precipitación, la elevación sobre el nivel del mar, la pendiente del terreno, la isotermalidad y la orientación de laderas (Ortíz-García et al. 2012; Pérez-Solano et al. 2012; Pérez-Solano y Mandujano 2013; Serna-Lagunes et al. 2014). En escala de paisaje, se ha determinado que la abundancia y uso de hábitat de los temazates tiene relación con las variables del paisaje natural, pero se encuentra más relacionada a variables de origen humano, principalmente la presión de cacería (Urquiza-Haas et al. 2009; 2011; Thornton et al. 2011; García-Marmolejo et al. 2013; 2015).
Mazama pandora utiliza los hábitats en relación a su disponibilidad por lo que se le ha considerado un generalista (Weber et al. 2008). En el caso del M. temama, se ha considerado un especialista pues prefiere sitios conservados (Weber et al. 2008; Bello-Gutiérrez et al. 2010). No obstante, la especie comúnmente se ha registrado en acahuales, zonas fragmentadas y sitios de cultivo (Medellín y Equihua 1998; Tejeda-Cruz et al. 2009; Bello-Gutiérrez et al. 2010; Thornton et al. 2011; Garmendia et al. 2013; García-Marmolejo et al. 2015). En el caso del microhábitat, se ha observado que la condición para la presencia de M. temama es que el sitio ofrezca una densa cobertura de protección vertical que le permita escapar de sus depredadores (Bello-Gutiérrez et al. 2010) y especies vegetales importantes para su alimentación (García-Marmolejo et al. 2015).
En México durante la temporada 2009-2010 se autorizó la cacería deportiva de 78 ejemplares de venados temazates (incluyendo ambas especies) en ocho UMA, todas ellas en el estado de Campeche (SEMARNAT 2010b). No obstante, existen evidencias de que de 21 UMA evaluadas en el estado, solo dos tienen las condiciones ambientales para mantener sus poblaciones viables (Escalante y Martínez-Meyer 2013) y en 19 no se ha garantizado la sostenibilidad ambiental de la explotación (Escalante y Martínez-Meyer 2013). A esta cantidad de ejemplares autorizados para cacería deportiva deben sumarse los cazados para subsistencia en Campeche, cuyo número es difícil de determinar, pero diferentes estudios indican que M. temama y M. pandora son de las especies silvestres más cazadas por los pobladores rurales en la Península de Yucatán (Hernández y Segovia 2010; Reyna-Hurtado y Tanner 2010; Santos-Fita et al. 2012). Es así que determinar las características del hábitat para los temazates en el estado de Campeche, es una tarea imprescindible para el manejo exitoso de la especie.
Para M. temama, se ha propuesto que la escala ideal para entender la respuesta de la especie a los procesos de cambios ambientales es la escala de paisaje (García-Marmolejo et al. 2015). En la actualidad se cuenta con información de ambas especies de temazates y su relación con el paisaje para San Luis Potosí (García-Marmolejo et al. 2015), Chiapas (Tejeda-Cruz et al. 2009), Quintana Roo (Urquiza-Haas et al. 2009) y Guatemala (Thornton et al. 2011). En Campeche no se conocen las características del hábitat a nivel paisaje que determinan la presencia de ninguna de las dos especies. Este estudio tiene como finalidad conocer las características del hábitat a nivel de paisaje, que determinan la presencia de las especies de Mazama , en la zona oeste del estado de Campeche donde se encuentran las Unidad de Manejo para la Conservación de la Vida Silvestre que tienen su aprovechamiento cinegético y de cacería de subsistencia.
Materiales y Métodos
Área de estudio. El estudio se realizó en un área de aproximadamente 3,500 km2 en los municipios de Champotón, Escárcega y Carmen en el oeste de Campeche, México (19° 15' N, -91º 20´ W; Figura 1). El clima es cálido subhúmedo, con una precipitación media anual

Figura 1 Mapa de localización del área de estudio en el oeste de Campeche, México. En la parte inferior de la figura se muestra la delimitación de las seis zonas en las que se dividió el área de estudio para determinar la presencia /ausencia de Mazama sp. en el oeste de Campeche, México. Los números corresponden a cada una de las regiones 1.- Chekubul-Cristalina; 2.- Chenkan, 3.- Chicbul-Km 74; 4.- Pixtun-Felipe Carrillo Puerto; 5.-Miguel Colorado; 6.- Centenario-Rigueña.
La población humana en el área de estudio se concentra en las poblaciones de Escárcega (29,500 hab), Champotón (30,000 hab) y Sabancuy (7,300 hab; INEGI 2013). Adicionalmente, en el área se encuentran 30 localidades entre 200 y 1,500 habitantes (INEGI 2013). El área se encuentra principalmente bajo el régimen de tierras ejidales con excepción de algunas propiedades hacía la costa donde es posible encontrar áreas privadas con extensiones que van de 2,000 a 5,000 ha (INEGI 2013).
Delimitación del área de estudio. El área de estudio se subdividió en seis regiones: 1) Chekubul-Cristalina; 2) Chenkan; 3) Chicbul-Km 74; 4) Pixtún-Felipe Carrillo Puerto; 5) Miguel Colorado; 6) Centenario-Rigueña. La delimitación se basó en las diferencias en las características ambientales regionales de uso de suelo y estatus de conservación de los ecosistemas naturales de cada una de las regiones (Tabla 1). Sin embargo, por practicidad los límites extremos de cada una de las regiones se ubicaron en las principales carreteras pavimentadas del área de estudio (Figura 1).
Tabla 1 Características de tamaño y uso de suelo de las seis regiones en las que se dividió el área de estudio. El uso de suelo se obtuvo a partir de la fotointerpretación de una serie de imágenes ASTER de 2008-2009 del área de estudio. Selvas tropicales conservadas (STC), Manejo agrícola y pecuario (MAyP), Manglares (M), vegetación costera inundable (VCI).

a Áreas cubiertas de selva mediana subperennifolia y baja inundable, además de acahuales en estado sucesional avanzado.
b Áreas donde la vegetación original ha sido eliminada y que en la actualidad están cubiertos por pastizales inducidos o áreas agrícolas.
c Áreas cubiertas por vegetación de pantano y sabanas naturales inundables.
d Áreas urbanas y áreas cubiertas por agua.
Detección de la especie. El uso de huellas y rastros para detectar la presencia de temazates ha sido un método eficiente para determinar la presencia en áreas extensas (e. g. Reyna-Hurtado y Tanner 2005; García-Marmolejo et al. 2015). La distribución geográfica de ambas especies, convergen en la zona de estudio (Gallina et al. 2007), lo que recientemente se ha confirmado por medio de cámaras trampa (Juárez-López 2015), asimismo, no se ha evidenciado diferenciación aparente entre las huellas de las especies del género Mazama (Angeli et al. 2014), por tanto, para este estudio no se diferenció entre especies y los datos se agruparon únicamente como Mazama sp.
Se realizó la búsqueda de huellas de temazates en 263 transectos (500 m de largo y 3 m de ancho), localizados en caminos madereros y senderos poco transitados entre agosto de 2008 y noviembre de 2010 en terrenos de 24 comunidades y ranchos privados del área de estudio. Cada transecto se georreferenció y separó de otro por al menos 500 m (Conroy 1996) y fue revisado solo una vez. La búsqueda de rastros se realizó caminando a pie a una velocidad aproximada de 4 km/h. Cada muestreo fue realizado por 5 personas en un horario de las 6 am a 6 pM. Se consideró que el transecto tenía presencia de temazates cuando se observó al menos una secuencia de huellas para lo cual se siguió lo propuesto por Mandujano (2005) para el venado cola blanca (Odocoileus virginianus). No se consideraron las secuencias de huellas encontradas fuera de transectos. Las huellas de Mazama sp. se distinguieron de las de otros ungulados de la región (e. g. Odocoileus virginianus, Tayassu pecari y Pecari tajacu) por su forma y tamaño (Aranda 2000).
Generación de capas de información geográfica y variables del paisaje. Se ha encontrado que la presencia de temazates se ve afectada por diversas variables a nivel paisaje de origen humano y natural (García-Marmolejo et al. 2013; 2015), por lo que en este estudio se decidió incluir ambos tipos de variables obtenidas a partir de diferentes fuentes (Tabla 2). La primera etapa del análisis fue generar con la ayuda de un Sistema de Información Geográfica (SIG, ArcGis 9.2) un mapa vectorial que contenía el punto medio de cada uno de los transectos muestreados (con o sin presencia de temazates), con el objetivo de definir el punto específico a partir del cual se generarían los valores de las variables del paisaje. Una vez determinados estos puntos, en el SIG se combinaron con mapas de uso de suelo y vegetación, carreteras y poblados del área de estudio. Esto nos permitió obtener las primeras cuatro variables del paisaje incluidas en el análisis.
Tabla 2 Variables utilizadas en el análisis de las características del hábitat a nivel de paisaje para el venado temazate en el oeste de Campeche, México.

El mapa de uso de suelo y vegetación se obtuvo a partir de un mosaico de siete imágenes de Satélite ASTER (15 x 15 m por pixel), tomadas entre marzo de 2008 y abril de 2009 (Japan Space Systems 2012). A partir de 350 puntos de verificación distribuidos en el área de estudio en los que se reconoció el uso de suelo y vegetación, se realizó una clasificación supervisada del mosaico de imágenes a partir de la clasificación de la reflectáncia captada y almacenada por el sensor ASTER en sus diferentes bandas multiespectrales (bandas visibles e infrarrojas) utilizando el algoritmo de máxima verosimilitud (MaxLike) en el software ENVI 4.5 (Exelis Visual Information Solutions 2008). La clasificación de la imagen permitió distinguir siete categorías de uso de suelo y vegetación, 1) agua, 2) vegetación costera inundable (que incluyó sabanas naturales y vegetación de pantano), 3) selvas tropicales conservadas (que incluyó selva mediana subperennifolia y baja inundable, además de acahuales en estado de sucesión avanzado), 4) manglar, 5) acahuales jóvenes (áreas agrícolas en estados de sucesión tempranos y medios), 6) áreas de manejo agrícola y pecuario (conformado por áreas donde la vegetación original ha sido eliminada y que en la actualidad están cubiertos por pastizales inducidos o áreas agrícolas) y 7) áreas urbanizadas.
El mapa de carreteras pavimentadas se obtuvo a partir de un mapa vectorial de carreteras de Campeche (INEGI 2013). El mapa de caminos no pavimentados, así como el de localidades urbanas se obtuvo a partir de la digitalización de los caminos y los polígonos de las localidades observados en las bandas visibles del mosaico de imágenes ASTER del área de estudio.
En base al mapa de uso de suelo y vegetación se obtuvieron las variables porcentaje de uso de suelo conservado y distancia a áreas agrícolas y pecuarias mayor a 10 ha. El porcentaje de uso de suelo conservado se obtuvo a partir de trazar en el SIG un circulo de radio de 500 m a partir del punto medio del transecto. Dentro de cada uno de los círculos generados, se cuantificó el área cubierta por tipos de uso de suelo y se obtuvo la proporción de cada uno de ellos, al dividirlo entre el área total del círculo de 500 m de radio. Para el análisis de uso de suelo conservado se consideró así a la vegetación costera inundable conservada, selvas tropicales y manglar. Una vez cuantificada la proporción de cada uso de suelo, las proporciones de uso de suelo conservado se obtuvieron al sumar cada categoría. La distancia a áreas agrícolas y pecuarias se generó utilizando la herramienta Nearest Features de ArcGis y en ella se obtuvo la distancia entre el punto medio del transecto y el límite del área de manejo agropecuario más cercana mayor a 10 ha. Se decidió por 10 ha, debido a que en el área de estudio el tamaño promedio de las parcelas que son permanentemente deforestadas y dedicadas a actividades pecuarias se encuentra entre 8 y 20 ha. En la región se ha observado que áreas menores son comúnmente utilizados como áreas de agricultura de roza tumba y quema y las áreas menores de agricultura son abandonadas una vez agotadas (Schmook et al. 2013). La variable distancia a localidad urbana más cercana se obtuvo midiendo la distancia entre el punto medio del transecto y el límite de la localidad urbana más cercana utilizando la herramienta Nearest Features de ArcGis. La distancia a la carretera pavimentada y camino no pavimentado más cercano se obtuvieron a partir de medir la distancia del punto medio del transecto y la carretera pavimentada más cercana y el camino no pavimentado más cercano utilizando la herramienta Nearest Features de ArcGis (Tabla 2).
En el área de estudio existen grandes extensiones de terrenos cercanos a la costa que se caracterizan por presentar largos periodos de inundación estacional. Esta característica ambiental podría estar afectando la presencia/ausencia de temazates, por lo que se decidió cuantificar esta variable y ocuparla en el análisis. Se generó un mapa de potencialidad de inundación a partir de un Modelo Digital de Elevación del área de estudio obtenido a partir del Earth Resources Observatory and Science Center (EROS 2008). En el mapa se consideró que el que todas aquellas áreas planas y cercanas a la costa que se encontraban entre -2 y 0 m fueran clasificadas como potencialmente inundables (Tabla 2). En el SIG se intersectó el mapa de inundación con el punto medio de los transectos y se determinó si el transecto era inundable o no inundable si se sobreponían espacialmente con estas categorías del mapa.
En el área de estudio la densidad de ganado varía en función del coeficiente de agostadero, el uso de suelo y la presencia de áreas inundables. La densidad de ganado se ha considerado como un indicador de la actividad humana (Lambin y Meyfroidt 2011) e influye de forma importante sobre la presencia de temazates en un área en particular (García-Marmolejo et al. 2015). Se utilizó el valor de densidad de ganado en toda el área de estudio obtenido a partir del mapa de Densidad Global de Ganado ver. 2.01 (Robinson et al. 2014; Tabla 2). El valor de densidad de ganado para cada uno de los transectos se obtuvo al extraer el valor del raster de densidad de ganado en el punto central del transecto para la búsqueda de temazates.
Análisis estadístico. Para conocer si existían variables de paisaje autocorrelacionadas, se realizó un análisis de correlación de Spearman con las variables originales (Zar 1999). En los casos en que se observó que existía correlación significativa entre las variables (P < 0.05). Se seleccionó la variable considerada con mayor importancia para explicar la presencia del temazate, esto, basados en la experiencia previa y en la información disponible sobre los temazates.
Se realizó un Análisis de Componentes Principales (ACP) en el programa InfoStat versión 2008 (Di Rienzo 2008), en el que se incluyeron las variables descriptoras del hábitat de los temazates a nivel paisaje, con el fin de identificar gradientes (eigenvectores) formados por las variables (características del hábitat), y a partir de ellos poder discriminar entre sitios potencialmente utilizables por el temazate, para lo cual se usaron los datos donde se confirmó la presencia de temazate (presencias) y todos los datos de transectos en los que no se tuvo registro de los mismo (ausencias). El ACP se realizó con las variables estandarizadas, debido a la diferencia de escalas y unidades en que se presentaron las variables (McGarigal et al. 2000).
Para determinar la relación existente entre la variable de respuesta (presencia/ausencia de temazate) y las variables explicativas (las variables del hábitat a un nivel de paisaje; Tabla 2), se llevó a cabo un análisis de regresión logística utilizando Modelos Lineales Generalizados (GLM), ya que estos permiten expresar de forma cuantitativa dicha relación entre estos dos tipos de variables (Guisan et al. 1999; Crawley 2007). Utilizando un modelo binomial, se realizó una regresión por pasos hacia atrás (Backwards) esta forma permite aprovechar la mayor variabilidad en el proceso de análisis (Crawley 2007), todo esto a partir del modelo completo (modelo que contiene todos las variables). La selección de los mejores modelos se realizó mediante el criterio de información de Akaike y la D2 (Burnham y Anderson 2002). El nivel de significancia utilizado en todos los análisis fue de P < 0.05. Para la realización de los análisis de regresión múltiple, se utilizó el módulo Biodiversity R (Kindt y Coe 2005), incluido en el programa estadístico R (R Development Core Team 2007).
Resultados
Se obtuvieron registros de temazates en 33 de los 263 transectos (12.55 %) localizados en las regiones de Centenario-La Rigueña, Miguel-Colorado y Chenkan. En las otras tres áreas no se encontraron registros de la especie (Chekubul-Cristalina, Chicbul-Km 74 y Pixtún-Felipe Carrillo Puerto; Tabla 3; Figura 2). El área Miguel Colorado fue donde se localizaron más evidencias de la presencia de temazate, seguido por Centenario-La Rigueña y finalmente Chenkan con sólo un transecto con presencia (Tabla 3).
Tabla 3 Número de transectos recorridos y número de presencias registradas de temazate para cada una de las seis regiones de estudio en las que se buscó determinar la presencia de temazates durante 2008-2010 en el oeste de Campeche, México.

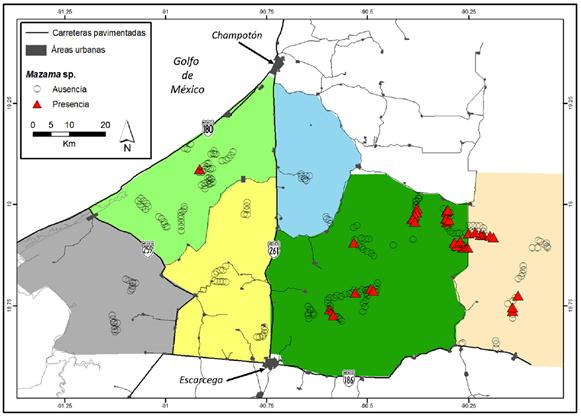
Figura 2 Mapa que muestra la localización de los transectos para determinar la presencia /ausencia de Mazama sp. en el oeste de Campeche, México. Los triángulos rojos indican los transectos donde se encontró la presencia de Mazama sp. y los círculos negros donde no se detectó su presencia.
Análisis de paisaje. El análisis de correlación indicó que las variables "distancia a la localidad urbana más cercana" y "distancia a caminos pavimentados", se encontraban autocorrelacionadas (coeficiente de correlación 0.83). Debido a que se consideró que localmente la distancia a caminos pavimentados podría tener mayor influencia en la presencia de temazates a nivel del área de estudio pues facilita el acceso a potenciales cazadores, además de que favorece la existencia de áreas perturbadas se decidió utilizar esta variable y eliminar del análisis distancia a localidad urbana más cercana. Las variables finales utilizadas en los análisis fueron las que se encuentran en la Tabla 2.
El ACP explicó en los primeros dos componentes el 64 % de la varianza, determinando que las variables de mayor peso en el primer componente fueron: porcentaje de selva conservada, densidad de ganado y distancia a potreros. En el segundo componente fueron la distancia al camino no pavimentado y porcentaje de inundación (Figura 3). Se observa en la gráfica de los componentes que el incremento en el porcentaje de selvas tropicales conservadas en los círculos de 500 m, se asocia a la presencia de temazates en el área de estudio. Por el contrario, únicamente se obtuvo una presencia en áreas con alta densidad ganadera, así como en áreas consideradas inundables. Factores como la presencia de carreteras pavimentadas o sin pavimentar no parecen tener un efecto tan importante sobre la presencia.

Figura 3 Representación gráfica de los valores para el Componente Principal 1 y 2 de cada uno de los transectos recorridos en el oeste del estado de Campeche, México en búsqueda de presencia de Mazama sp. Los cuadros rojos representan transectos en los que se registró presencia de las especies. La descripción y nombres completos de las variables se encuentran en Tabla 2.
La regresión logística para identificar el peso que las diferentes variables tenían sobre la presencia/ausencia de temazates en el área de estudio estaba fuertemente desbalanceada debido a que sólo en 33 transectos se obtuvo presencia, comparados con 230 con ausencia. El desbalance en los modelos estadísticos tiene efectos importantes sobre la prueba, provocando poca sensibilidad de la misma en caso de que existieran pequeñas desviaciones sobre el supuesto de varianzas iguales, además de reducir notablemente el poder de la prueba (Montgomery 2012). Para resolver este problema, se decidió balancear el experimento utilizando el mismo número de presencias y ausencias con el fin de reducir el peso que podrían tener las variables asociadas a las ausencias en el análisis. Para este fin, del conjunto de 230 transectos con ausencia de temazate se seleccionaron al azar 33 (el mismo número de transectos con presencia), los cuales fueron los únicos incluidos en la regresión logística de presencia/ausencia.
La regresión logística mostró que la utilización de todas las variables la D2 del modelo completo es de 21.08 %, mientras que el mejor modelo con las variables de porcentaje conservado y densidad de ganado variables la D2 es de 20.71 % (Tabla 4). El mejor modelo muestra que la presencia de temazates en el área de estudio es mayor cuando se incrementa el porcentaje de uso de suelo conservado alrededor del transecto, mientras disminuye al incrementarse la densidad de ganado (Tabla 4).
Tabla 4 Resultados de los modelos lineales generalizados entre las variables descriptoras del hábitat y los registros de presencia-ausencia de temazates en el oeste del estado de Campeche, México. Se muestra el mejor modelo que incluye las variables que determinan la presencia o ausencia de temazates. La descripción y nombres completos de las variables se encuentran en Tabla 2.

a Criterio de Información de Akaike.
b Diferencia en el criterio de información de Akaike.
c Presencia/Ausencia de Temazate = PORC_CON+GANADERÍA + DIST_PAVIME + DIST_POTRE + DIST_BRECH +INUNDA.
d Presencia/Ausencia de Temazate = -2.53 + 0.019(PORC_CON)-0.274(GANADERÍA).
Discusión
Los resultados del estudio muestran que solo en tres regiones se encontró presencia de temazates (Miguel Colorado, Centenario-La Rigueña y Chenkan). Los factores para la obtención de este resultado pueden ser metodológicos, antropogénicos o ambientales. Tres de las regiones donde no se encontraron rastros fueron las que contaron con el menor número de transectos, lo que podría tener efectos en la detectabilidad de la especie. Sin embargo, en concordancia con este estudio, otros estudios no han detectado temazates en la región de, Chicbul-Km 74 (Rangel-Negrin et al. 2014) ni en Pixtún-Felipe Carrillo Puerto (Juárez-López 2015). No se descarta la presencia de temazates en las tres regiones donde no fueron encontrados. La ausencia de registros pudiera ser debido a sesgos metodológicos, que redujeran las probabilidades de detectar especies en bajas densidades como ya ha ocurrido en las regiones de Chenkan y Miguel Colorado, sitios en los que intensos esfuerzos de muestreo con cámaras trampa ha permitido la detección de especies de mamíferos crípticas (e. g. Tayassu pecari, Mustela frenata o Panthera onca; Contreras-Moreno et al. 2015; Hidalgo-Mihart et al. 2014, 2015). Es así que es necesario que en futuros esfuerzos se conbienen métodos como huellas y cámaras trampa.
Los resultados de la regresión logística y el ACP mostraron que la presencia de selvas tropicales conservadas y una mayor distancia a áreas de potreros favorece la presencia de temazates, mientras que una mayor densidad de ganado (actividad asociada a presencia humana y transformación de hábitat natural a pastizales inducidos) disminuye la probabilidad de su detección. Las áreas Chicbul-Km 74, Chekubul y Pixtún-Felipe Carrillo Puerto son las tres áreas donde no se detectó su presencia y al mismo tiempo los sitios en las que la mayor parte de los hábitats originales han sido transformados (ya sea que se mantienen como áreas agrícolas, o han sido abandonadas y se encuentran como acahuales jóvenes). Es posible que la ausencia de hábitats favorables para Mazama en estas áreas pudiera estar determinando, ya sea la ausencia o las bajas densidades asociadas a su falta de detección.
Las regiones de Chekubul-Cristalina y Chenkan son áreas donde una proporción importante de los hábitats naturales se asocian a prolongados periodos de inundación que ocurren en el área (manglares y vegetación costera inundable). El ACP mostró que la condición de inundación de las áreas, tiene gran importancia para la determinación de la presencia de los temazates. Se obtuvo un registro en el área de Chenkan, mientras que para Chekubul y Cristalina no se detectó la especie. El registro en la región de Chenkan fue durante el mes de diciembre de 2008, y durante esa época la región se encontraba inundada. En la región de Calakmul, los temazates prefirieron el bosque inundable en las aéreas con cacería (Reyna-Hurtado y Tanner 2005). En Sudamérica se ha comprobado que los ungulados pueden cambiar su comportamiento cuando los sitios se encuentran inundados (Hofmann et al. 2015), y tomar tres tipos de decisiones, vivir de forma subacuática, mantener ámbitos hogareños muy grandes que le permitan migrar a sitios fuera de la inundación o quedarse en el sitio y resguardarse en islas de selva (Husson 1978), esto último se observó en el Amazonas donde M. americana y Pecari tajacu permanecieron durante la temporada de inundación, resguardándose en islas de selva a diferencia de Tapirus terrestris y Tayassu pecari que abandonaron la zona inundable (Bodmer 1990).
En estudios realizados en zonas inundables de Campeche y Yucatán por fototrampeo (Juárez-López 2015; Hernández-Pérez et al. 2015), así como para la determinación del uso de fauna por pobladores locales (León y Montiel 2008), no han registrado la presencia de especies de Mazama en este tipo de ambientes, por lo que es probable que la especie en caso de encontrarse, este presente en bajas densidades en estas áreas. Lo anterior marca la necesidad de determinar el efecto que tienen los largos periodos de inundaciones de la zona costera sobre su habilidad para sobrevivir en estas áreas. Esta necesidad se hace mayor ante el evidente cambio climático que sufren los ecosistemas actuales, en la Amazonía se ha observado en los últimos años, que las crecientes extremas, han afectado substancialmente poblaciones de mamíferos terrestres, ya que extremos climáticos ahora se producen en lapsos de tiempo tan cortos que las especies no tienen tiempo de recuperarse (Bodmer et al. 2014).
Las áreas de Miguel Colorado y Centenario-La Rigueña fue donde se obtuvieron el 97 % de los registros de temazate. Asimismo, es importante resaltar que sólo se registraron en alrededor del 20 % de los transectos recorridos en estas áreas, lo que podría ser un indicador de que la abundancia es relativamente baja en comparación con otras áreas como Calakmul (Reyna-Hurtado y Tanner 2005) o la Lacandona (Tejeda-Cruz et al. 2009). Los resultados en el ACP y la regresión logística indican que Miguel Colorado y Centenario-La Rigueña tienen gran importancia para las especies de Mazama en el oeste de Campeche, pues ambas áreas se caracterizan por tener una cubierta vegetal constituida en su mayor parte por selvas conservadas con baja densidad de ganado. La presencia de estos venados es mayor en áreas donde el grado de conservación de los ecosistemas naturales es relativamente alto (García-Marmolejo et al. 2015). Aunque la cacería podría estar influenciando su presencia en un sitio particular aún más que las variables del paisaje (Urquiza-Haas et al. 2007; 2009). En el caso particular del área de estudio la presencia de grandes áreas con selvas conservadas, favorecen la presencia de las especies de Mazama . El impacto que la cacería de subsistencia tiene en su presencia en estas regiones no se determinó, debido a que no se cuentan con estudios específicos sobre el tema. En este sentido, es probable que en selvas conservadas de Miguel Colorado y Centenario-La Rigueña, se extraigan un gran número de temazates como ocurre en Calakmul (Reyna-Hurtado y Tanner 2010); en comunidades del este de Campeche y Quintana Roo (Santos-Fita et al. 2012) y en el sur de Yucatán (Hernández y Segovia 2010). En el sur de México, una parte importante de la cacería de subsistencia se localiza en áreas cercanas a cultivos (Santos-Fita et al. 2012). Basado en este hecho, es posible que la ausencia de terrenos de cultivo en los grandes parches de bosque conservado, reduzca el efecto de la cacería de subsistencia. Pese a esto, en el área existen caminos madereros en uso o abandonados que permiten el libre acceso a cazadores de las comunidades cercanas con fines de comercialización local (F. Contreras obs. pers.).
La constante deforestación y perdida del hábitat en toda la región oeste de Campeche (Soto-Galera et al. 2010) apunta a una situación crítica para los temazates en la región. Estudios sobre deforestación en la región que rodea Laguna de Términos indican que de 1974 a 2001 se perdió aproximadamente el 41 % de la vegetación original, de la cual 62,000 ha correspondieron a selvas tropicales (Soto-Galera et al. 2010), lo que podría haber reducido en gran medida la presencia y abundancia de las poblaciones en el área. Es necesario considerar este hecho al momento de otorgar autorizaciones de cacería deportiva en la región, exigiendo a los proponentes estudios específicos para el cálculo de densidades y tasas de cosecha de ejemplares que impliquen un muestreo sistemático y robusto de las poblaciones de los temazate en las UMA. Si bien los planes de manejo actuales exigen estimaciones poblacionales, en muchos casos estas son realizadas por personas sin experiencia (Contreras-Moreno y Hidalgo-Mihart 2015), por lo que debe garantizarse que las estimaciones sean rigurosas y combinando métodos que garanticen la detección de la especie como el uso de cámaras trampa. Así mismo, es necesario que se generen alternativas de manejo comunitario que auto-regulen la cacería de subsistencia, para alcanzar un aprovechamiento sustentable de los recursos.
Es necesario determinar el efecto que tienen las áreas inundables costeras sobre los temazates. La escasa presencia en este tipo de ambientes, sumado a la ausencia de registros de la especie en estudios previos en ambientes costeros inundables de Campeche, indicaría que este tipo de ambientes son poco utilizados. Este hecho es importante desde el punto de vista de conservación y manejo regional de las especies, pues es posible que una importante sección de hábitats conservados de la costa del Golfo de México no tenga aptitud para sostener poblaciones importantes de estas especies.

