










Servicios Personalizados
Revista
Articulo
 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista mexicana de ciencias forestales
versión impresa ISSN 2007-1132
Rev. mex. de cienc. forestales vol.11 no.58 México mar./abr. 2020 Epub 17-Jun-2020
https://doi.org/10.29298/rmcf.v11i58.549
Scientific article
Arboreal vegetation of the Cerro Tres Puntas de Pilasca, (Salas-Motupe), Lambayeque, Perú
1Facultad de Ciencias Biológicas, Universidad Nacional Pedro Ruiz Gallo, Lambayeque, Perú.
2Laboratorio General de Biotecnología, Universidad Nacional Pedro Ruiz Gallo, Lambayeque, Perú.
From March to June, 2018 the seasonally dry forest Cerro Tres Puntas of Pilasca (Salas-Motupe, Lambayeque, Perú) was studied, in order to obtain information on its structure and its floristic composition. In a linear transect, about 11.4 ha, specimens of woody plants with DBH ≥ 5.0 cm were assessed. 410 individuals representing 17 species, 17 genera and 10 families were recorded. The most abundant species were Vachellia macracantha (Fabaceae 154 individuals) and Celtis iguanaea (Cannabaceae 55 individuals). The families with the highest number of species were Fabaceae (7) and Malvaceae (2). The basimetric area was 343.86 m2 ha-1 highlighting Ficus obtusifolia with 139.23 m2 ha-1 and Beilschmiedia sulcata with 120.90 m2 ha-1. The Importance Value Index for the species reached the main values in F. obtusifolia (49.34), V. macracantha (46.75) and B. sulcata (41.57), while the Family Value Index was widely higher in the Fabaceae family (111.86). While these results show that this forest is a precarious version of the region’s, F. obtusifolia individuals between 35-39.9 m high and 210-219.9 cm DBH were found. This study will help to accomplish an efficient conservation and reforestation program in order to improve the Cerro Tres Puntas forest, located in one of the poorest areas of the nation.
Key words Basimetic area; seasonally dry forest; diametric distribution; floristic diversity; floristic structure; Lambayeque
De marzo a junio de 2018 se estudió el bosque estacionalmente seco en Cerro Tres Puntas de Pilasca (Salas-Motupe, Lambayeque, Perú) con la finalidad de obtener información sobre su estructura y composición florística. En un transecto lineal de 11.4 ha, aproximadamente, se evaluaron especímenes de plantas leñosas con DAP ≥ 5.0 cm. Se registraron 410 individuos que representan a 17 especies, 17 géneros y 10 familias. Los taxones más abundantes fueron Vachellia macracantha (Fabaceae 154 individuos) y Celtis iguanaea (Cannabaceae 55 individuos) y las familias mejor representadas corespondieron a Fabaceae, con siete especies y Malvaceae, con dos. El área basal total fue de 343.86 m2 ha-1, en la que destacan Ficus obtusifolia con 139.23 m2 ha-1 y Beilschmiedia sulcata con 120.90 m2 ha-1. El Índice de Valor de Importancia alcanzó los mayores valores en F. obtusifolia (49.34), V. macracantha (46.75) y C. ruizii (41.57); en tanto que el Índice de Valor de la Familia fue más alto para Fabaceae, con 111.86, seguida de Moraceae (46.74) y Lauraceae (45.33). Si bien, los resultados muestran que este bosque es una versión precaria de los existentes en la región, todavía se identificó la presencia de individuos de F. obtusifolia entre 35 - 39.9 m de altura y DAP de 210 a 219.9 cm. El presente estudio contribuirá a desarrollar un programa de conservación, y de reforestación eficaz para mejorar el bosque del Cerro Tres Puntas, ubicado en una de las áreas más pobres del país.
Palabras clave Área basal; bosque estacionalmente seco; distribución diamétrica; diversidad florística; estructura florística; Lambayeque
Introduction
The seasonally dry forest (BES, for its acronym in Spanish)) in the Neotropics are distributed from northern Mexico to southern Brazil and make up 66.7 % of this type of vegetation in the world (Miles et al., 2006). In Perú, BES are threatened and under-studied ecosystems and comprise several subtypes (Linares-Palomino, 2006; Linares-Palomino and Pennington, 2007). The mountain and plain BES of the north coast are very fragile; they extend from the Peninsula Santa Elena, in southern Ecuador, to several regions of northwestern Perú, among which are Tumbes, Piura, Lambayeque and northern La Libertad, as well as the lower layer of the Marañón Valley. These two areas communicate through the Porculla Pass, although they are distinct phytogeographic units (Brack and Mendiola, 2004). It is also known as the Tumbesina endemism region, which is shared by Ecuador and Perú, and it is considered one of the most important areas worldwide for this reason (Aguirre et al., 2006).
Some studies have revealed the large number of endemic species present in the BES of northern and northwestern Perú, with about 26 % of them disseminated in various communities, such as Prosopis limensis Benth. (carob) and Colicodendron scabridum Seem. (zapote) (Sagástegui et al., 1999). But not only the levels of diversity and endemism refer to plants but also to vertebrates such as Tremarctos ornatus Cuvier, 1825 (Spectacled bear), Lagidium Perúanum Meyen, 1833 (Vizcacha), Penelope albipennis Taczanowski, 1878 (Pava aliblanca) and Bothrops barnetti Parker, 1994 (Macanche), among others (Aguilar, 1994).
The works on the BES, although not very abundant, have been significant, among which those carried out by Weberbauer (1922; 1930; 1936; 1945) are particularly interesting, as he contributed floristic and phytogeographic aspects of the flora of Perú and elaborated a map of vegetation, hitherto unsurpassed, with emphasis on BES. In the past decade, the annotated list of woody species and plant formations in the BES of the Tumbesian Endemic Region of Ecuador and Perú (Aguirre et al., 2006) as well as woody plants as a new online tool for taxonomic ecological and biodiversity studies stand out (Linares-Palomino and Pennington, 2007).
Based on data from forest inventories, the structure and diversity patterns of the BES of two conservation areas of the Northwest Biosphere Reserve (Perú), the Cerros de Amotape National Park and the Tumbes Reserved Zone were analyzed and discussed (Leal-Pinedo and Linares-Palomino, 2005). Also addressed were the spatial distribution patterns of tree species such as Eriotheca ruizzii (K. Schum.) A. Robyns (pasayo), Bursera graveolens Triana & Planch. (palo santo), Caesalpinia glabrata Kunth (charán) and Cochlospermum vitifolium (Willd.) Spreng. (polo polo), in a BES of the Cerros de Amotape National Park, in northwestern Perú (Linares-Palomino, 2005). In the surroundings of Jaén (Cajamarca, Perú), in relicts of the seasonally dry tropical forest, eight inventories were made on woody vegetation, endemisms and conservation status (Marcelo-Peña, 2008).
On the other hand, in order to design maps and classify vegetation in seasonal ecosystems of the dry forests of Piura (Perú), with data from floristic inventories, six groups of vegetation were identified, of which five coincided with types of vegetation described 100 years ago by Weberbauer (1922; 1930; 1936; 1945) (La Torre-Cuadros and Linares-Palomino, 2008). In the Ayabaca province (Piura), the edaphic characteristics and floristic composition of La Menta and Timbes BES were evaluated (Rasal et al., 2011).
In the Lambayeque region (Perú), some studies have also been carried out at the BES. In the Chaparrí Ecological Reserve, which is part of the Tumbes-Chocó-Magdalena hot spot of biodiversity, a structural pattern and floristic inventory was developed, from which a fairly impoverished forest became evident, in relation to other BES in the region (Linares-Palomino and Ponce-Álvarez, 2009). In this same Chaparrí Private Ecological Reserve, the phenology (vegetative development, flowering and fruiting) of 17 plant species that feed the white wing turkey (pava aliblanca), an endemic species seriously threatened (Martos et al., 2009), was assessed.
Based on the above, in addition to the fact that the natural forests of Perú have been poorly studied, that the anthropic pressure that leads to their predation is increasing, that climate change continues to alter the patterns of plant distribution, among other factors (Steffen et al., 2011), the present work was carried out with the aim to determine the floristic composition of the BES of Cerro Tres Puntas with the purpose of facilitating the elaboration of reforestation, conservation and utilization plans and, thus, contributing to the knowledge of the natural ecosystems of the country.
Materials and Methods
Study area
The study area is located between 6°14'16.23" S - 79°35'34.81" W and 6°12'46.46" S - 79°33'59.58" W, and includes Cerro Tierra Blanca (1 986 masl), Cerro Gato (1 545 masl), both belonging to the Salas district, and Cerro Tres Puntas (1 590 masl), from the Motupe district, in the province and region of Lambayeque, Perú (Figures 1a -b); these three hills cover around 2 975 ha.
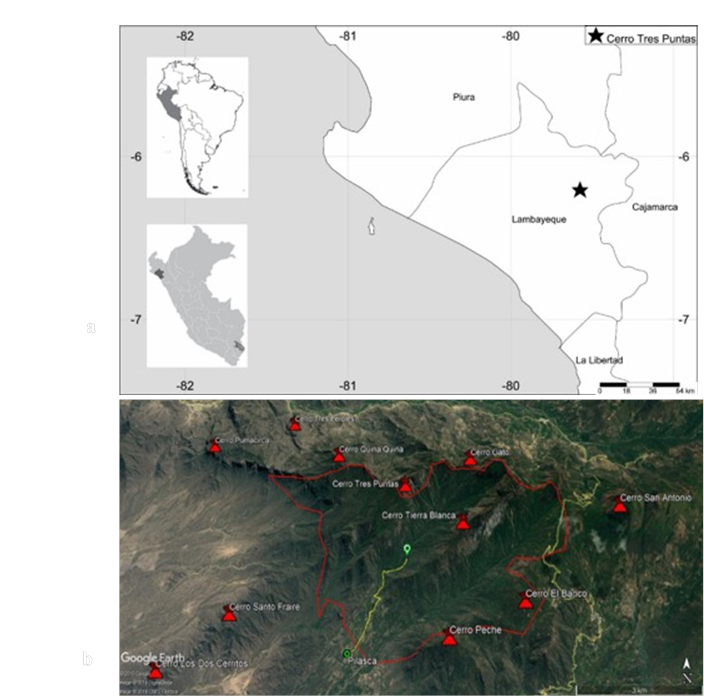
Source: Google Earth (2019).
Figure 1 a) Map of the Lambayeque region where Cerro Tres Puntas is located and the village of Pilasca is indicated with a star. b) Area comprising the Tierra Blanca, Cerro Gato and Cerro Tres Puntas of the BES (Salas-Motupe, Lambayeque, Perú).
The Pilasca village is one of the poorest in Lambayeque (Gobierno Regional de Lambayeque, 2013) where the maize agriculture (Zea mays L.), yucca (Manihot esculenta Crantz) and fruit trees, as well as cattle and goat breeding is for subsistence; it grows in small clearings in the forest, where there is no seed selection, fertilizers or control of pests and diseases. The peasants of Pilasca have used the forest species since ancient times for the manufacture of furniture, construction of rustic houses and pens as well as firewood as fuel, which is why Loxopterygium huasango Spruce ex Engl. (hualtaco), Handroanthus chrysanthus (Jacq.) S. O. Grose (yellow guayacán), Colicodendron scabridum (Kunth) Seem. (zapote), Prosopis limensis Benth. (carob tree) and Cedrela sp. (cedar) practically no longer exist.
In the geological aspect, the study area corresponds to the litho-stratigraphic unit Salas Group (Oi) of the Paleozoic or Salas Formation, (Pi-s) Lower Ordovician (480 to 470 million years) (Gobierno Regional de Lambayeque, 2013; López, 2013).
The area in which the research was conducted is a seasonally dry semi-dense forest ecosystem of hill and mountain (Bes-cm) (Minam, 2019). The data was collected in a place between 373 and 720 masl; the extreme point A corresponds to the entrance of the Pilasca village (373 m), where the evaluation began, to the extreme point E (720 masl), where it ended, whose topography is rugged with huge boulders and deep ravines, dry and scarce stony clearings. The temperature, relative humidity and precipitation data were taken from the Jayanca CO-Meteorological Station (LA Vic) (78 masl), the closest to Cerro Tres Puntas de Pilasca, from which the following data were recorded: maximum and minimum temperature and relative humidity of March 2018, 34.0 and 19.0 ºC and 78.4 %, respectively, while in June 2018, 26.2 and 15.0 ºC and 79.1 %, respectively. In these four months of evaluation, the precipitation was 0.0 mm day-1 (Senamhi, 2018).
Sampling
Field work was carried out from March to June 2018 and the method proposed by Gentry (1982; 1995a) was used for the sampling of woody plants, with substantial modifications from the difficult access and dangerous geography of the land. A linear sampling was applied taking as a baseline a relatively safe trail that the locals use to occasionally access the top of Cerro Tres Puntas de Pilasca (Figure 2).

Four sampling units were established in a total area of 3 801 m, designated with points A (373 masl) -B (456 masl), BC (565 masl), CD (677 masl) and DE (720 masl), which covered 958 m, 961 m, 968 m and 914 m, respectively. From the center of the trail were measured 15 m to the right and 15 m to the left, making a total of 30 m which allowed to evaluate about 114 030 m2, with a GPS Garmin 60CSX and a measuring tape. All woody individuals whose DBH (1.3 m from the base of the trunk) was ≥ 5.0 cm were recorded, taken as a measure of the basimetric area with a measuring tape.
For individuals with branched trunks, the diameter of each branch was measured at a height of 1.3 m from the base of the trunk, with a diametric tape Forestry Suppliers, Inc. (Modelo 283D/5M). As the species evaluated were known, only botanical samples were taken for registration and deposit in the Herbarium of the Universidad Nacional Pedro Ruiz Gallo de Lambayeque (HPR Herbarium). Likewise, herbaceous species, present in the collection area, were picked up and pressed for subsequent taxonomic identification. For families, the phylogenetic postulates of the Phylogeny Group of Angiosperms were followed (APG, 2009; 2016).
Information processing and analysis
To determine the abundance or density of the sampled species, the classification of Maleheiros and Rotta (1982) was used, which classifies as rare or scarce species with 1-6 individuals, not very abundant with 7-25 and abundant with more than 25 specimens. The horizontal structure of the forest was described from the distribution of the number of trees by diameter and height class (Hawley and Smith, 1972). Phytosociological analyzes were done through the FITOPAC 2.1 freeware (Shepherd, 2010). With this support, the following parameters were calculated: Relative Density (DER), Relative Frequency (FER), Relative Basimetric Area (AB) or Relative Coverage (COB), Importance Value Index (IVI) and Importance Value for the Family (VIF), formulas that were described by Rangel and Velásquez (1997).
Results
Floristic composition
In the 114 030 m2 studied, 410 individuals of trees and shrubs with DAP≥5.0 cm were inventoried, corresponding to 17 species, 17 genera and 10 families, registered in 47 sampling units (Table 1). To these arboreal and shrub species were added numerous herbaceous plants with different growth habits such as vines, epiphytes, sub-shrubs, flowers, among others, collected with flowers and fruits in May 2018, among which can be mentioned: Salvia occidentalis Sw. (Lamiaceae), Psittacanthus chanduyensis Eichler (Loranthaceae), Croton pavonis Müll. Arg. (Euphorbiaceae), Scutia spicata (Wild.) Weberb. (Rhamnaceae), Alternanthera pubiflora Kuntze (Amaranthaceae), Dalechampia scandens L. (Euphorbiaceae), Cordia macrocephala (Desv.) Kunth (Boraginaceae), Plumbago scandens L. (Plumbaginaceae), Heliotropium angiospermum Murray (Boraginaceae), Waltheria ovata Cav. (Malvaceae), Wedelia latifolia DC. (Asteraceae), Cardiospermum corindum L. (Sapindaceae), Tessaria integrifolia Ruiz & Pav. (Asteraceae), Pseudogynoxys cordifolia (Cass.) Cabrera (Asteraceae), Tetramerium nervosum Nees (Malvaceae), Scoparia dulcis L. (Scrophulariaceae)´and Cestrum auriculatum L’Hér. (Solanaceae).
Table 1 Main tree mensuration features of the species present at the assessed area of BES Cerro Tres Puntas (Salas-Motupe, Lambayeque, Perú).
| Family/Species | Common name | Growth hab. | Num. indiv./% | Sampling unit | DBH (cm) | Total height (m) | Crown diam. (m) |
|---|---|---|---|---|---|---|---|
| Anacardiaceae | |||||||
| Loxopterygium huasango Spruce ex Engl. | Hualtaco | Tree | 20/ 4.9 | 4 | 27.79 | 12.83 | 8.84 |
| Boraginaceae | |||||||
| Cordia lutea Lam. | Overo | Tree/⁓ Shrub | 29/ 7.1 | 3 | 1.83 | 5.97 | 6.52 |
| Burseraceae | |||||||
| Bursera graveolens (Kunth) Triana & Planch. | Palo santo | Tree | 4/ 1.0 | 2 | 22.45 | 11.00 | 6.88 |
| Fabaceae | |||||||
| Albizia multiflora (Kunth) Barneby & J.W. Grimes | Angolo | Tree | 8/ 2.0 | 4 | 33.39 | 14.75 | 11.43 |
| Caesalpinia glabrata Kunth | Charán | Tree | 5/ 1.2 | 1 | 11.92 | 7.00 | 6.92 |
| Erythrina smithiana Krukoff | Frejolillo | Tree | 32/ 7,8 | 3 | 36.03 | 11.69 | 9.14 |
| Pithecellobium excelsum (Kunth) Mart. | Chaquiro | Tree/⁓ Shrub | 21/ 5.1 | 4 | 11.04 | 7.33 | 9.64 |
| Prosopis limensis Benth. | Algarrobo | Tree | 9/ 2.2 | 1 | 37.31 | 16.56 | 12.89 |
| Senna atomaria (L.) Irwing & Barneby | Chapa | Tree | 15/ 3.7 | 2 | 9.83 | 5.47 | 4.23 |
| Vachellia macracantha (Willd.) Seigler & Ebinger | Faique | Tree/⁓ Shrub. | 154/ 37.6 | 4 | 16.34 | 8.54 | 8.99 |
| Lauraceae | |||||||
| Beilschmiedia sulcata (Ruiz & Pav.) Kosterm | Palta de zorro | Tree | 1/ 0.2 | 1 | 124.10 | 12.00 | 22.10 |
| Malvaceae | |||||||
| Eriotheca ruizii (K. Schum.) A. Robyns | Pasayo | Tree | 6/ 1.5 | 1 | 31.3 | 10.58 | 7.37 |
| Guazuma ulmifolia Lam. | Guazumo | Tree | 11/ 2.7 | 3 | 27.42 | 12.83 | 10.52 |
| Moraceae | |||||||
| Ficus obtusifolia Kunth | Higuerón | Tree | 17/ 4.2 | 4 | 120.06 | 19.35 | 24.00 |
| Olacaceae | |||||||
| Ximenia americana L. | Ciruelillo | Tree | 17/ 4.2 | 3 | 11.40 | 10.06 | 5.9 |
| Sapindaceae | |||||||
| Sapindus saponaria L. | Choloque | Tree | 6/ 1.5 | 3 | 17.6 | 10.00 | 6.45 |
| Cannabaceae | |||||||
| Celtis iguanaea (Jacq.) Sarg. | Palo blanco | Tree | 55/ 13.4 | 4 | 38.48 | 14.62 | 12.99 |
| Total | 410 | 47 | 26.20 | 10.45 | 9.81 | ||
Growth hab. = Growth habit; Num. indiv. = Number of individuals; DBH=Diameter at l Breast Height; Crown diam. = Crown diameter.
Structure
Of the 17 inventoried species, few specimens of Cordia lutea Lam., Vachellia macracantha (Willd.) Seigler & Ebinger and Pitecellobium excelsum (Kunth) Benth. showed bush growth habit with more than 10 shafts per individual, in some cases. Of the total recorded species, only six - L. huasango, V. macracantha, P. excelsum, Albizia multiflora (Kunth) Barneby & J.W. Grimes, Ficus obtusifolia Kunth and Celtis iguanaea (Jacq.) Sarg.-were found in the four sampling units, while Caesalpinia glabrata Kunth, P. limensis, E. ruizii and Beilschmiedia sulcata (Ruiz & Pav.) Kosterm. were only found in one of them.
The largest number of the 410 inventoried individuals corresponded to the third sampling unit (565-677 masl) with 194 (47.32 %), which is attributed to the fact that it is a very rugged and stony territory, where it is not possible to develop agricultural activities ; in contrast, in the second sampling unit (456-565 masl) the lowest number was counted, 42 individuals (10.24 %), which can be explained because it is the area with the greatest agricultural vocation, which has led to a dramatic depredation of this part of the forest (Table 2).
Table 2 Distribution per sampling unit of the individuals from the species found in the assessed area of the BES Cerro Tres Puntas (Salas-Motupe, Lambayeque, Perú).
| Species | Sampling units | |||||||
|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | |||||
| No | % | No | % | No | % | No | % | |
| Beilschmiedia sulcata (Ruiz & Pav.) Kosterm | 0 | 0.0 | 0 | 0.0 | 0 | 0.0 | 1 | 1.56 |
| Bursera graveolens Triana & Planch. | 2 | 1.82 | 0 | 0.0 | 2 | 1.03 | 0 | 0.0 |
| Caesalpinia glabrata Kunth | 0 | 0.0 | 0 | 0.0 | 5 | 2.58 | 0 | 0.0 |
| Celtis iguanaea (Jacq.) Sarg. | 9 | 8.18 | 7 | 16.67 | 23 | 11.86 | 16 | 25.00 |
| Cordia lutea Lam. | 10 | 9.10 | 2 | 4.76 | 17 | 8.76 | 0 | 0.0 |
| Erythrina smithiana Krukoff | 0 | 0.0 | 1 | 2.38 | 26 | 13.40 | 5 | 7.81 |
| Eriotheca ruizii (K. Schum.) A. Robyns | 0 | 0.0 | 0 | 0.0 | 6 | 3.09 | 0 | 0.0 |
| Ficus obtusifolia Kunth | 1 | 0.91 | 1 | 2.38 | 3 | 1.55 | 12 | 18.75 |
| Guazuma ulmifolia Lam. | 4 | 3.64 | 5 | 11.90 | 2 | 1.03 | 0 | 0.0 |
| Loxopterygium huasango Spruce ex Engl. | 2 | 1.82 | 4 | 9.52 | 12 | 6.19 | 2 | 3.13 |
| Pithecellobium excelsum (Kunth) Mart. | 11 | 10.0 | 4 | 9.52 | 5 | 2.58 | 1 | 1.56 |
| Albizia multiflora (Kunth) Barneby & J.W. Grimes | 2 | 1.82 | 2 | 4.76 | 2 | 1.03 | 2 | 3.13 |
| Prosopis limensis Benth. | 9 | 8.18 | 0 | 0,0 | 0 | 0.0 | 0 | 0.0 |
| Sapindus saponaria L. | 1 | 0.91 | 3 | 7.14 | 2 | 1.03 | 0 | 0.0 |
| Senna atomaria (L.) Irwing & Barneby | 9 | 8.18 | 6 | 14.29 | 0 | 0.0 | 0 | 0.0 |
| Vachellia macracantha (Willd.) Seigler & Ebinger | 47 | 42.73 | 7 | 16.67 | 81 | 41.75 | 19 | 29.69 |
| Ximenia americana L. | 3 | 2.73 | 0 | 0.0 | 8 | 4.12 | 6 | 9.38 |
| Total = 410 individuals | 110 | 26.83 | 42 | 10.24 | 194 | 47.32 | 64 | 15.61 |
The upper canopy consisted of dominant trees of F. obtusifolia and P. limensis, although with very dispersed individuals. The tallest trees were F. obtusifolia with a height of 19.35 m followed by P. limensis (with 16.56 m), P. excelsum (with 14.75 m) and C. iguanaea, (with 14.62 m) in a distribution of heights that is not quite an inverted "J", as expected for natural forests and regular forests.
The crown diameter was larger in F. obtusifolia (with 24.00 m) followed by B. sulcata (with 22.10 m) (Table 3). In a similar way, in the horizontal structure of the forest, it was determined that in transect 4, individuals of greater height (13.59 m) and diameter (48.99 cm) were registered, which is much greater than the diameter recorded in transect 1 with 20.11 cm, which is half of its size (Table 4).
Table 3 Distribution of individuals by diametric class, height class and density of the species present in the assessed area of the BES Cerro Tres Puntas (Salas-Motupe, Lambayeque, Perú).
| Diametric class (cm) | Individuals (Number) | Diametric class (m) | Individuals (Number) | Density | Species (Number) |
|---|---|---|---|---|---|
| 5 - 9.9 | 86 | 0 - 4.9 | 9 | Abundant | 4 |
| 10 - 19.9 | 128 | 5 - 9.9 | 205 | Little abundant | 8 |
| 20 - 29.9 | 77 | 10 - 14.9 | 129 | Rare o scarce | 5 |
| 30 - 39.9 | 53 | 15 - 19.9 | 44 | ||
| 40 - 49.9 | 29 | 20 - 24.9 | 15 | ||
| 50 - 59.9 | 17 | 25 - 29.9 | 5 | ||
| 60 - 69.9 | 4 | 30 - 34.9 | 2 | ||
| 70 - 79.9 | 2 | 35 - 39.9 | 1 | ||
| 80 - 89.9 | 2 | ||||
| 90 - 99.9 | 0 | ||||
| 100 - 109.9 | 1 | ||||
| 110 - 119.9 | 2 | ||||
| 120 - 129.9 | 3 | ||||
| 130 - 139.9 | 1 | ||||
| 140 - 149.9 | 0 | ||||
| 150 - 159.9 | 0 | ||||
| 160 - 169.9 | 1 | ||||
| 170 - 179.9 | 1 | ||||
| 180 - 189.9 | 0 | ||||
| 190 - 199.9 | 1 | ||||
| 200 - 209.9 | 0 | ||||
| 210 - 219.9 | 2 | ||||
| Total | 410 | 410 | 17 |
Table 4 Horizontal structure of the forest from the distribution of tree species by height and diameter class of the species present at the transects of the assessed area of the BES Cerro Tres Puntas (Salas-Motupe, Lambayeque, Perú).
| Transect | Number of individuals | Height (m) |
Diameter (cm) |
|---|---|---|---|
| Mean ± sd | Mean ± sd | ||
| 1 | 110 | 10.21 ± 4.35 | 20.11 ± 12.28 |
| 2 | 42 | 10.33 ± 4.99 | 21.65 ± 12.52 |
| 3 | 194 | 9.58 ± 3.74 | 23.11 ± 24.25 |
| 4 | 64 | 13.59 ± 6.71 | 48.99 ± 44.92 |
Density
The species with the highest number of individuals and, thus, with the highest relative density (DER) was V. macracantha, con 154 (37.6 %), followed by C. iguanaea with 55 (13.4 %) and Erythrina smithiana Krukoff. with 32 (7.8 %); therefore, Fabaceae (59.4 %) and Cannabaceae (13.1 %) accomplished the highest values, while fue B. sulcata (0.2 %) had the lowest (Tables 1 and 4). Relative frequency (FER) achieved the greatest value (8.51) in several species, in addition to V. macracantha, en L. huasango, P. excelsum, A. multiflora, F. obtusifolia and C. iguanaea. Four species, V. macracantha, C. lutea, E. smithiana and C. iguanaea have been considered abundant, eight little abundant and five, rare or scarce, such as B. graveolens, C. glabrata, E. ruizii, C. iguanaea and B. sulcata, the latter just with one specimen (Table 3).
Basimetric area
The maximum value of DAP corresponded to B. sulcata with 124.10 cm followed by F. obtusifolia, (120.06 cm), C. iguanaea (38.48 cm) and P. limensis (37.31 cm), although B. sulcata only recorded one individual and P. limensis, eight (Table 1). With the exception of the 5-9.9 cm DAP class with 86 individuals, the analysis of the diametric distribution indicated that trees in the transects were in the 10-19.9 cm DAP (205) and 20-29.9 cm DAP classes (129), whose proportion decreased as the diameters increased (Table 3). The accumulated value of the basimetric area was 343.86 m2 ha-1 from B. sulcata 139.28 m2 ha-1 and F. obtusifolia (120.90 m2 ha-1) The largest relative coverage (COB) or wider relative basal area coincides with the same species (B. sulcate with 39.19 and F. obtusifolia with 36.68), followed very distantly, by C. iguanaea (3.77), P. limensis (3.54) and E. smithiana (3.30), and at the family level, Lauraceae (39.19), Moraceae (36.68) and Fabaceae (11.28) were outstanding (Table 5).
Table 5 Ecological variables of the species present in the assessed area of the BES Cerro Tres Puntas (Salas-Motupe, Lambayeque, Perú).
| Family/Species | Number of species | Number of individuals | DER | FER | Basimetric area (m2) | Relative basimetric area (COB) | IVI | VIF |
|---|---|---|---|---|---|---|---|---|
| Anacardiaceae | ||||||||
| Loxopterygium huasango Spruce ex Engl. | 20 | 4.88 | 8.51 | 6.06 | 1.97 | 15.35 | ||
| Total Family | 1 | 20 | 4.88 | 8.51 | 6.06 | 1.97 | 15.35 | 12.73 |
| Boraginaceae | ||||||||
| Cordia lutea Lam. | 29 | 7.07 | 6.38 | 0.92 | 0.30 | 13.74 | ||
| Total Family | 1 | 29 | 7.07 | 6.38 | 0.92 | 0.30 | 13.74 | 13.27 |
| Burseraceae | ||||||||
| Bursera graveolens (Kunth) Triana & Planch. | 4 | 0.98 | 4.26 | 3.95 | 1.28 | 6.53 | ||
| Total Family | 1 | 4 | 0.98 | 4.26 | 3.95 | 1.28 | 6.53 | 8.16 |
| Fabaceae | ||||||||
| Albizia multiflora (Kunth) Barneby & J.W. Grimes | 8 | 1.95 | 8.51 | 8.75 | 2.84 | 13.30 | ||
| Caesalpinia glabrata Kunth | 5 | 1.22 | 2.13 | 1.11 | 0.36 | 3.71 | ||
| Erythrina smithiana Krukoff | 32 | 7.80 | 6.38 | 10.19 | 3.30 | 17.49 | ||
| Pithecellobium excelsum (Kunth) Mart. | 21 | 5.12 | 8.51 | 0.96 | 0.31 | 13.94 | ||
| Prosopis limensis Benth. | 9 | 1.95 | 2.13 | 10.93 | 3.54 | 7.62 | ||
| Senna atomaria (L.) Irwing & Barneby | 15 | 3.66 | 4.26 | 0.76 | 0.25 | 8.16 | ||
| Vachellia macracantha (Willd.) Seigler & Ebinger | 154 | 37.56 | 8.51 | 2.09 | 0.68 | 46.75 | ||
| Total Family | 7 | 244 | 59.40 | 40.50 | 34.79 | 11.28 | 110.97 | 111.86 |
| Lauraceae | ||||||||
| Beilschmiedia sulcata (Ruiz & Pav.) Kosterm | 1 | 0.24 | 2.13 | 120.90 | 39.19 | 41.57 | ||
| Total Family | 1 | 1 | 0.24 | 2.13 | 120.90 | 39.19 | 41.57 | 45.33 |
| Malvaceae | ||||||||
| Eriotheca ruizii (K. Schum.) A. Robyns | 6 | 1.46 | 2.13 | 9.74 | 2.49 | 6.80 | ||
| Guazuma ulmifolia Lam. | 11 | 2.68 | 6.38 | 6.53 | 1.91 | 10.98 | ||
| Total Family | 2 | 17 | 4.16 | 8.52 | 16.27 | 4.40 | 17.78 | 20.32 |
| Moraceae | ||||||||
| Ficus obtusifolia Kunth | 17 | 4.16 | 8.51 | 139.28 | 36.68 | 49.34 | ||
| Total Family | 1 | 17 | 4.16 | 8.53 | 139.28 | 36.68 | 49.34 | 46.74 |
| Olacaceae | ||||||||
| Ximenia americana L. | 17 | 4.15 | 6.38 | 0.92 | 0.33 | 10.86 | ||
| Total Family | 1 | 17 | 4.15 | 6.38 | 0.92 | 0.33 | 10.86 | 10.38 |
| Sapindaceae | ||||||||
| Sapindus saponaria L. | 6 | 1.46 | 6.38 | 2.69 | 0.79 | 8.63 | ||
| Total Family | 1 | 6 | 1.46 | 6.38 | 2.69 | 0.79 | 8.63 | 8.15 |
| Cannabaceae | ||||||||
| Celtis iguanaea (Jacq.) Sarg. | 55 | 13.41 | 8.51 | 13.38 | 3.77 | 25.70 | ||
| Total Family | 1 | 55 | 13.41 | 8.51 | 13.38 | 3.77 | 25.70 | 23.08 |
| Total | 17 | 410 | 100 | 100 | 343.86 | 100 | 300 | 300 |
Importance Value Index per Species (IVI) and Importance Value Index per Family (VIF)
The IVI analysis of the 17 assessed speciesEl análisis del IVI de las 17 especies highlighted F. obtusifolia (49.34) over V. macracantha (46.75) and C. ruiziana (41.57) y, and so, Fabaceae (110.97), Moraceae (49.34) and Lauraceae (41.57) (Table 5). These same families recorded the highest VIF (Fabaceae, 111.86; Moraceae, 46.74 and Lauraceae, 45.33), while those ewith the loest numbers were Burseraceae (8.16) and Sapindaceae (8.15) (Table 5).
Discussion
The sampling area is classified as seasonally dry mountain forests on the north coast that covers the BES over 500 masl, and covers the western flanks of the Andes in the Tumbes, Piura and Lambayeque regions to La Libertad, including part from Cajamarca (Linares-Palomino and Pennington, 2007). However, as indicated by La Torre-Cuadros and Linares-Palomino (2008), this altitudinal limit is quite variable, debatable and depends on several factors. As sampling began at 373 masl, it engulfed a small area of the plain BES on the north coast that covers the BES below 500 masl along the western flanks of the Andes from Tumbes to La Libertad (Linares- Palomino and Pennington, 2007). This would be the reason why, under the conditions of Pilasca, there was little presence of carob (P. limensis) and only a single specimen of zapote (C. scabridum) and others of vichayo (Beautempsia avicennifolia (Kunth) Gaudich.); the latter outside the sampling area.
These authors also pointed out, that the dry mountain BES at the north coast as those of the interandine system of the Marañón river, are the richest in woody species with 193 and 184, respectively. Such information has been confirmed by the study conducted in the seasonally dry tropical forest (BTES) of the Marañón valley, in northern Perú, in which 440 woody species, and 143 (33 %) endemic, as well as two endemism centers were identified within the valley, with 56 % of species and 78 % of genera found in Peruvian BTES (Marcelo-Peña et al., 2016).
In the analysis of species diversity patterns ─4660 woody plant species─ of BTES from Latin America and the Caribbean, it was determined that the outstanding floristic rotation observed implies the need to establish multiple conservation areas to protect such a great flora diversity (DRYFLOR et al., 2016).
Even though an annotated list of the BES of Ecuador and other related studies (Svenson, 1946; Gentry, 1995b; Aguirre et al., 2006) indicated that the BES of Ecuador and Perú, especially those of the coast, formed a single geographical unit, with a high number of endemic species (Tumbesino Endemic Center), the few species (17), genera (17) and families (10), inventoried in the Cerro Tres Puntas de Pilasca, would demonstrate a high degree of depredation of the original flora. This would confirm what has already been observed in other BES such as Chaparrí in Lambayeque (Linares-Palomino and Ponce-Álvarez, 2009; Martos et al., 2009) and La Menta and Timbes in Piura (Rasal et al., 2011).
On the other hand, other factors such as the diversity and density of species and precipitation gradients, which decrease as the BES extend south, should be considered, including the Lambayeque and La Libertad regions.
Starting from the rugged and dangerous topography of the land, the linear sampling that was used in the present investigation made it possible to evaluate a large area that covered 11.4 ha, a sampling that has already been used in the rapid botanical inventories (IBR) in which it woody vegetation, endemisms and the conservation status of the BES of Jaén (Perú) were addressed (Marcelo-Peña, 2008), based on the methodology of Hawthorne and Abu-Juam (1995) and Gordon et al. (2004).
In spite of the interest in BES in recent years (Espinosa et al., 2012; Aguirre and Geada-López, 2017), in the few works carried out in Ecuador and Perú it was observed that the sampling methods were very variable between the use of rectangular transects and linear sampling, as well as in the size of the sample units, from 0.25 to 6.5 hectares and in the DAP of the sampled species, from ≥ 1.0 to 10 cm. The difference between the number of individuals, species, genera and families should be added, which is usually higher in protected natural areas such as the Cerros de Amotape National Park (Linares-Palomino, 2005) and very depressed in other historically depredated areas, as the Chaparrí Ecological Reserve (Linares-Palomino and Ponce-Álvarez, 2009).
From all of them, the closest to the described one here is that used in La Menta and Timbes in Piura (Rasal et al., 2011), where, despite the fact that woody species with only DAP ≥ 1.0 cm were registered, the number of inventoried individuals was significantly smaller, although there was quite similarity in the number of species, genera and families.
In their study at the Chaparrí Ecological Reserve, Linares-Palomino and Ponce-Álvarez (2009) pointed out the complication of establishing comparisons of inventories in the seasonally dry forests of the region, from the great variation between the sampling methodologies used, altitudinal floors, total sampling area, DAP of the sampled specimens, conservation status and depredation of the sampling area at the time of its realization, and even in the prioritization of the information collected, among other factors.
The previous arguments are confirmed by several studies in Perú such as those in the forests of Jaén, Cajamarca (Marcelo-Peña, 2008), La Menta and Timbes in Ayabaca, Piura (Rasal et al., 2011),Biosphere Northeaster Reserve of Perú (Cerros de Amotape National Park and Tumbes Reserved Zone) (Leal-Pinedo and Linares-Palomino, 2005), Piura coastal plains (La Torre-Cuadros and Linares-Palomino, 2008), Cerros de Amotape National Park (Linares-Palomino, 2005) and Chaparrí Ecological Reserve, Lambayeque (Linares-Palomino and Ponce-Álvarez, 2009); however, it has not been possible to establish rigorous patterns of comparison between all this valuable information and the study carried out at the Cerro Tres Puntas de Pilasca.
In the plant formations of the BES of Ecuador and Perú, lists of many woody species were made for the deciduous forest, semi deciduous dry forest mixed with evergreen species, deciduous forest and the dry thorny thicket (Aguirre et al., 2006; Aguirre and Kvist, 2009); however, in the actual work, only eight species were recorded: P. limensis, C. lutea, P. excelsum, C. glabrata, V. macracantha, E. smithiana, B. graveolens and L. huasango, which somehow, would indicate the strong pressure that Cerro Tres Puntas de Pilasca has suffered in recent years, even though it was possible to find sporadic individuals from Bougainvillea peruviana Nees & Mart., Bauhinia aculeata L., C. vitifolium and C. scabridum although they were outside the linear transect.
The trend of distribution of heights that does not fit exactly with the inverted “J”, described by Lamprecht (1990) and Günter et al. (2011), would exhibit the succession characteristic of the forest from the constant and selective disturbances exerted by the local inhabitants, keeping it in a secondary succession stage (Rasal et al., 2012). Although the referred experience did not include individuals with DAP <5.0 cm, although with a plant height <4.9 m, its results allow a thorough understanding of the regeneration process of the species present in the higher diameter and height classes, as recommended by García et al. (2010) studying the structure and floristic diversity of two natural forests in Cauca, Colombia.
In BES La Menta and Timbes of Piura (Perú), about 55.0 % of the sampled individuals had a DAP between 0-10.0 cm with some L. huasango and B. graveolens trees with DAP between 50-60 cm for La Menta, and Ceiba trischistandra Bakh. and E. ruizii with DAP between 80-90 cm for Timbes (Rasal et al., 2011). Regarding these five indicated species, at BES Cerro Tres Puntas only E. ruizii was found. Likewise, of the 410 registered individuals, 196 had DAP ≥ 20.0 cm, where three species, V. macracantha, C. iguanaea and E. smithiana contributed 58.8 % of the total.
Very few studies conducted in the dry forests of Perú have collected information or performed analyzes related to AB (Basimetric Area), IVI (Importance Value Index) and VIF (Importance Value for the Family); however, Linares-Palomino and Ponce-Álvarez (2009) calculated an AB of 11.74 m2, in which E. ruizii and L. huasango stood out with 4.40 and 4.12 m2, respectively in the Chaparrí Ecological Reserve (Lambayeque). In the dry forests of La Menta and Timbes (Piura), Rasal et al. (2011) estimated an AB of 128.86 m2 in La Menta, in which L. huasango (45.97 m2) and B. graveolens (34.69 m2) stood out, and in Timbes an AB of 196.63 m2 was obtained, with special contribution from E. smithiana (47.16 m2) and C. trischistandra (38.88 m2). In both cases, the AB was much lower than the one reported in the current study in which the AB was 343.86 m2 ha-1, which indicated that despite the fact that the Cerro Tres Puntas forest has suffered a strong anthropic pressure, it has not been as dramatic as what happened in Chaparrí and La Menta and Timbes, where even the Chaparrí forest was defined as an impoverished version of the seasonally dry forests of the region (Linares- Palomino and Ponce-Álvarez, 2009).
In this same community, four species recorded IVI numbers over 30.0: E. ruizii (58.22), L. huasango (44.93), C. lutea (33.76) and B. graveolens (30.08), which belong to the Malvaceae, Anacardiaceae, Boraginaceae and Burseraceae families, respectively (Linares-Palomino and Ponce-Álvarez, 2009); while in La Menta, B. graveolens (50.0), L. huasango (43.0) and C. lutea (36.0) had the highest values, and in Timbes, C. lutea (62.0), E. smithiana (40.0) and C. trischistandra (25.0) (Rasal et al., 2011); also, as mentioned above, in the work in the dry forests of the province of Loja, Aguirre et al. (2013). defined the five species with the highest IVI: C. trischistandra, Simira ecuadorensis (Standl.) Steyerm., Tabebuia chrysantha G. Nicholson, E. ruizii and Terminalia valverdeae A. H. Gentry, although no quantitative information was provided. When comparing these results with those of the present study, notorious differences are observed since the species that stood out were F. obtusifolia (49.34), V. macracantha (46.75) and B. sulcata (41.57), which were relevant in other seasonal forests. It is possible to hypothesize that the depredation of the species of these forests has not followed a definite pattern, and this would have been subject to the preference for the most valuable timber species and to the same protection that would have given them the local inhabitants, where, certainly, on the Cerro Tres Puntas, predation has been more selective.
The VIF results of the research described for the Cerro Tres Puntas de Pilasca, in which the families Fabaceae (111.86), Moraceae (46.74) and Lauraceae (45.33) stood out, disagree widely with those referred to Chaparrí (Linares-Palomino and Ponce- Álvarez, 2009), La Menta and Timbes (Rasal et al., 2011), since none of the families listed in these works has been recorded in the study carried out in the area of interest.
Conclusions
The study in Cerro Tres Puntas de Pilasca (Salas-Motupe, Lambayeque) contributes to the knowledge of the natural ecosystems of Perú, specifically that of seasonally dry forests, by determining their floristic structure and composition. In the context of biodiversity, the results obtained describe its plant richness, which is a valuable contribution to the conservation and management of these resources, and ecological data as a preliminary approach to the dynamics of the BES in the region.
It is suggested to extend the studies to other areas in the same Tres Puntas Hill, in adjacent geographical areas, in other BES of Lambayeque such as those of Tongorrape, Ñaupe and Tocmoche and even increase the altitude range of sampling. Also, conduct carbon capture assessments and identify the species that would most contribute to mitigate climate change and thus, to promote their large-scale multiplication for reforestation purposes.
Acknowledgements
The authors thank the Research Vice Chancellor of the Universidad Nacional Pedro Ruiz Gallo (UNPRG) de Lambayeque, Dr. Ernesto Hashimoto Moncayo, for the facilities to achieve the actual study, to the Administration staff of UNPRG: Héctor Valdiviezo Castillo and Melissa J. Saldarriaga Heros as well as to Br. Wendy Herrera Moncada, for the technical support in writing of the manuscript. Also, to the community members of Pilasca: Froilán Sánchez and Isabel Valencia, for their priceless guide services in the several ascents to the Tres Puntas Hill.
REFERENCES
Aguilar, P. 1994. Características faunísticas del norte del Perú. Arnaldoa 2(1):77-102. [ Links ]
Aguirre, Z., R. Linares P. y L. P. Kvist. 2006. Especies leñosas y formaciones vegetales en los bosques estacionalmente secos de Ecuador y Perú. Arnaldoa 13(2):324-350. ISSN: 1815-8242. [ Links ]
Aguirre, Z. y L. Kvist. 2009. Composición florística y estructura de bosques estacionalmente secos en el sur-occidental de Ecuador, Provincia de Loja, municipios de Macará y Zapotillo. Arnaldoa 16(2):87-99. Doi: 10.13140/2.1.1288.0008. [ Links ]
Aguirre, Z., Y. Betancourt y G. Geada. 2013. Composición florística y estructura de los bosques secos de la Provincia de Loja, Ecuador. Arnaldoa 20:117-128. ISSN: 1815-8242 [ Links ]
Aguirre, Z. y G. Geada-López. 2017. Estado de conservación de los bosques secos de la provincia de Loja, Ecuador. Arnaldoa 24(1):207-228. Doi:10.22497/arnaldoa.241.24107. [ Links ]
Angiosperm Phylogeny Group III (APG III). 2009. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: AGP III. Botanical Journal of the Linnean Society 161:105-121. Doi:10.1111/j.1095-8339.2009.00996.x. [ Links ]
Angiosperm Phylogeny Group IV (APG IV). 2016. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IV. Botanical Journal of the Linnean Society 181:1-20. Doi:10.1111/boj.12385. [ Links ]
Brack, A. y C. Mendiola. 2004. Ecología del Perú. PNUD. Asociación Editorial Bruño. Lima, Perú. 495 pp. [ Links ]
Dryflor, K. Banda-R, A. Delgado-Salinas, K. G. Dexter, R. Linares-Palomino, A. Oliveira-Filho, D. Prado, M. Pullan, C. Quintana, R. Riina, G.M. Rodriguez M., J. Weintritt, P. Acevedo-Rodriguez, J. Adarve, E. Álvarez, A. Aranguren B., J. Camilo Arteaga, G. Aymard, A. Castaño, N. Ceballos-Mago, Á. Cogollo, H. Cuadros, F. Delgado, W. Devia, H. Dueñas, L. Fajardo, Á. Fernández, M. A. Fernández, J. Franklin, E. H. Freid, L. A. Galetti, R. Gonto, R. González-M., R. Graveson, E. H. Helmer, Á. Idárraga, R. López, H. Marcano-Vega, O. G. Martínez, H.M. Maturo, M. MacDonald, K. Maclaren, O. Melo, F. Mijares, V. Mogni, D. Molina, N. P. Moreno, J. M. Nassar, D. M. Neves, L. J. Oakley, M. Oatham, A. R. Olivera-Luna, F. F. Pezzini, O. J. Reyes Dominguez, M. E. Rios, O. Rivera, N. Rodriguez, A. Rojas, T. Sarkinen, R. Sánchez, M. Smith, C. Vargas, B. Villanueva and R.T. Pennington. 2016. Plant diversity patterns in neotropical dry forests and their conservation implications. Science 353(6306): 1383-1387. Doi: 10.1126/science.aaf5080. [ Links ]
Espinosa, C., M. De la Cruz, A. L. Luzuriaga y A. Escudero. 2012. Bosques tropicales secos de la región Pacífico Ecuatorial: diversidad, estructura, funcionamiento e implicaciones para la conservación. Ecosistemas 21(1-2):167-179. [ Links ]
García, C., C. Suárez y M. Daza. 2010. Estructura y diversidad florística de dos bosques naturales (Buenos Aires, Dpto. Cauca, Colombia). Revista Biotecnología en el Sector Agropecuario y Agroindustrial (Universidad del Cauca) 8(1):74-82. [ Links ]
Gentry, A. H. 1982. Patterns of neotropical diversity. Evolutionary Biology 15:1-84. Doi: 10.1007/978-1-4615-6968-8_1. [ Links ]
Gentry, A. H. 1995a. Patterns of diversity and floristic composition in Neotropical Montane Forests. In: Churchill, S. P., H. Balslev, E. Forero and J. L. Luteyn (eds.). Biodiversity and Conservation of Neotropical Montane Forests. The New York Botanical Garden. New York, NY, USA. pp. 103-126. [ Links ]
Gentry, A. H. 1995b. Diversity and floristic composition of neotropical dry forests. In: Bullock, S. H., H. A. Mooney and E. Medina (eds.). Seasonally Dry Tropical Forests. Cambridge University Press, Cambridge, UK. pp. 146-194. [ Links ]
Gobierno Regional de Lambayeque. 2013. Estudio Geológico del Departamento de Lambayeque. Lambayeque, Perú. 60 p. [ Links ]
Google Earth. 2019. Salas, Perú. 17M657260.19 m E9312897.45 m S, Altitud del ojo 11.65 km. CNES/Airbus 2019. http://www.earth.google.com (2 de diciembre, 2019). [ Links ]
Gordon, J. E., W. D. Hawthorne, A. Reyes-García, G. Sandoval and A. J. Barrance. 2004. Assesing landscapes: a case study of tree and shrub diversity in the seasonally dry tropical fortest of Oaxaca, Mexico and southern Honduras. Biological Conservation 117:429-442. Doi:10.1016/j.biocon.2003.08.011. [ Links ]
Günter, S., M. Weber, B. Stimm and R. Mosandl (eds.). 2011. Silviculture in the Tropics. Tropical Forestry 8. Springer-Verlag. Berlin, Germany. 560 p. [ Links ]
Hawley, R. C. y D. M. Smith. 1972. Silvicultura Práctica. Ediciones Omega. Barcelona, España. 544 p. [ Links ]
Hawthorne, W. D. and M. Abu-Juam. 1995. Forest protection in Ghana. With particular reference vegetation and plant species. Forest Inventory and Management Planning Brach. Forestry Department Kumasi, Ghana. IUCN/ODA/Forest Department, Republic of Ghana. https://www.iucn.org/es/content/forest-protection-ghana-particular-reference-vegetation-and-plant-species (20 de septiembre de 2019). [ Links ]
Lamprecht, H. 1990. Silvicultura en los trópicos. Deutsche Gesellschaft fur Technische Zusammenartbeit (GTZ). Alemania. 334 p. [ Links ]
La Torre-Cuadros, M. A. y R. Linares-Palomino. 2008. Mapas y clasificación de vegetación en ecosistemas estacionales: un análisis cuantitativo de los bosques secos de Piura. Revista Peruana de Biología 15(2):31-42. Doi: 10.15381/rpb.v15i1.1668. [ Links ]
Leal-Pinedo, J. M. y R. Linares-Palomino. 2005. Los bosques secos de la Reserva de Biosfera del Noroeste (Perú): Diversidad arbórea y estado de conservación. Caldasia 27(2):195-211. [ Links ]
Linares-Palomino, R. 2005. Spatial distribution patterns of trees in a seasonally dry forest in the Cerros de Amotape National park, nortwestern Peru. In: Weigend, M., E. Rodríguez and C. Arana. (comps.). Bosques Relictos del NO de Perú y SO de Ecuador. Revista Peruana de Biología 12:317-326. [ Links ]
Linares-Palomino, R. 2006. Phytogeography and floristics of seasonally dry tropical forests in Peru. In: Pennington, R. T., G. P. Lewis and J. A. Ratter (eds.). Neotropical Savannas and Seasonally Dry Forests: Plant Diversity, Biogeography and Conservation. CRC Press. Boca Raton, FL, USA. pp. 257-279. Doi: 10.1201/9781420004496. [ Links ]
Linares-Palomino, R. and S. I. Ponce-Álvarez. 2009. Structural patterns and floristics of a seasonally dry forest in Reserva Ecológica Chaparrí, Lambayeque, Perú. Tropical Ecology 50:305-314. [ Links ]
Linares-Palomino, R. y R. T. Pennington. 2007. Lista anotada de plantas leñosas en bosques estacionalmente secos del Perú - una nueva herramienta en internet para estudios taxonómicos, ecológicos y de biodiversidad. Arnaldoa 14(1):149-152. [ Links ]
López, M. 2013. Geomorfología y Geología Histórica de Lambayeque. https://www.academia.edu/14393809/Geomofología_de Lambayeque . (15 de enero de 2019). [ Links ]
Maleheiros, D. E. O. y E. Rotta. 1982. Levantamiento horizontal de uma mata de araucaria do primero planalto paranaense. Boletim de Pesquisa. Florianópolis 4:1-39. [ Links ]
Marcelo-Peña, J. L. 2008. Vegetación leñosa, endemismos y estado de conservación en los bosques estacionalmente secos de Jaén, Perú. Revista Peruana de Biología 15:43-52. Doi: 10.15381/rpb.v15i1.1669. [ Links ]
Marcelo-Peña, J. L., I. Huamantupa, T. Särkinen and M. Tomazello. 2016. Identifying conservation priority areas in the Marañón valley (Peru) based on floristics inventories. Edinburgh Journal of Botany 73:95-123. Doi:10.1017/S0960428615000281. [ Links ]
Martos, J. R., M. Scarpati, C. Rojas y G. E. Delgado. 2009. Fenología de algunas especies que son alimento para la pava aliblanca Penelope albipennis. Revista Peruana de Biología 15: 51-58. Doi: 10.15381/rpb.v15i2.1721. [ Links ]
Miles, L., A. C. Newton, R. S. DeFries, C. Ravilious, I. May, S. Blyth, V. Kapos and J. E. Gordon. 2006. A global overview of the conservation status of tropical dry forests. Journal of Biogeography 33:491-505. Doi: 10.1111/j.1365-2699.2005.01424.x. [ Links ]
Ministerio del Ambiente (Minam). 2019. Mapa Nacional de Ecosistemas del Perú. Lima, Perú. 60 p. [ Links ]
Rangel, J. O. y A. Velásquez. 1997. Métodos de estudio de la vegetación. In: Rangel, J. O., P. Lowy y M. Aguilar (eds.). Colombia Diversidad Biótica II. Instituto de Ciencias Naturales, Universidad Nacional de Colombia. Bogotá, Colombia. pp. 59-82. [ Links ]
Rasal, M., J. Troncos, C. Lizano, O. Parihuamán, D. Quevedo, C. Rojas y G. Delgado. 2011. Características edáficas y composición florística del bosque estacionalmente seco La Menta y Timbes, región Piura, Perú. Ecología Aplicada 10(2):61-74. Doi: 10.21704/rea.v10i1-2.414. [ Links ]
Rasal, M., J. Troncos, C. Lizano, O. Parihuamán, D. Quevedo, C. Rojas y G. Delgado. 2012. La vegetación terrestre del bosque montano de Lanchurán (Piura, Perú). Caldasia 34(1):1-24. [ Links ]
Sagástegui, A., M. O. Dillon, I. Sánchez, S. Leiva y P. Lezama. 1999. Diversidad Florística del Norte del Perú. Tomo I. Editorial Graficart. Trujillo, Perú. 228 p. [ Links ]
Shepherd, G. J. 2010. Fitopac 2.1.2: Manual do usuario. UNICAMP. Campinas, São Paulo, Brasil. 93 p. [ Links ]
Servicio Nacional de Meteorología e Hidrología (Senamhi). 2018. Estación CO-Meteorológica Jayanca (LA Vic). Lambayeque, Perú. 4 p. [ Links ]
Steffen, W., Å. Persson, L. Deutsch, J. Zalasiewicz, M. Williams, K. Richardson, C. Crumley, P. Crutzen, C. Folke, L. Gordon, M. Molina, V. Ramanathan, J. Rockström, M. Scheffer, H. J. Schellnuber and U. Svedin. 2011. The anthropocene: from global change to planetary stewardship. Ambio 40:739-761. Doi:10.1007/s1328 0-011-0185-x. [ Links ]
Svenson, H. K. 1946. Vegetation of the coast of Ecuador and Peru and its relation to the Galapagos Islands. I. Geographical relations of the flora. American Journal of Botany 33(5):394-426. Doi: 10.2307/2437128. [ Links ]
Weberbauer, A. 1922. Die Vegetationskarte der peruanischen Anden zwischen 5º und 17oS. Petermanns Geographische Mitteilungen 68:89-91. [ Links ]
Weberbauer, A. 1930. Die Pflanzendecke Nordperus im Departamento Tumbez una angrenzenden Teilen des Departamento Piura (3º30’ - 5os. Br.). Botanische Jahrbücher 63:29-48. [ Links ]
Weberbauer, A. 1936. Phytogeography of the Peruvian Andes. Field Museum of Natural History, Botanical Series 13:13-81. [ Links ]
Weberbauer, A. 1945. El mundo vegetal de los Andes peruanos. Ministerio de Agricultura, Dirección de Agricultura, Estación Experimental Agrícola de La Molina, Lima. Lima, Perú. 776 p. [ Links ]
Received: March 27, 2019; Accepted: February 10, 2020
 Este es un artículo publicado en acceso abierto bajo una licencia
Creative Commons
Este es un artículo publicado en acceso abierto bajo una licencia
Creative Commons
