










Services on Demand
Journal
Article
 text in
text in  English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista mexicana de ciencias forestales
Print version ISSN 2007-1132
Rev. mex. de cienc. forestales vol.9 n.50 México Nov./Dec. 2018
https://doi.org/10.29298/rmcf.v9i50.231
Articles
Vertical epiphyte distribution and its phylogeny in a mesophilic cloud forest of Tamaulipas
1Instituto de Ecología Aplicada, Universidad Autónoma de Tamaulipas. México.
2Facultad de Ciencias Forestales, Universidad Autónoma de Nuevo León. México.
The mesophyllic cloud forest in El Cielo Biosphere Reserve is close to its boreal distribution limit. Epiphyte plants are an important component of the biodiversity of these forests, although they are little studied. Vascular epiphytes were studied in an altitudinal gradient of a cloud forest, determining the preference of epiphytic species for a host or phorophyte. There were differences in the distribution of epiphytes over the phorophytes, as well as in the vertical distribution of each epiphytic species on its host. A total of 33 epiphytic species were recorded, of which 29 showed at least one association with a host species. There were 24 species of trees, with which the epiphytes showed at least one association with 16 of them. The number of epiphytic species was greater at lower altitudes. According to the number of individuals in each of the sections of the phorophyte, epiphytes showed preference to zones III and IV, corresponding to the basal and middle parts of the branches of the host trees. Phylogeny was not related with epiphyte species distribution in host tree zones or in the studied elevations. The total number of species found is small compared to similar studies carried out in more southern latitudes, perhaps because of a general decrease in species richness, associated with the distance from the equator.
Key words: Phylogenetic analysis; cloud forest; phorophytes; El Cielo biosphere reserve; species richness; habitat zoning
El bosque mesófilo de montaña en la Reserva de la Biósfera El Cielo se localiza cerca de su límite de distribución boreal. Las plantas epífitas son un componente importante de la biodiversidad de estos bosques, aunque son poco estudiados. En este trabajo se estudiaron las epífitas vasculares en un gradiente altitudinal, y se determinó su preferencia por un hospedero; también se observó que existen diferencias en su distribución sobre los forofitos, así como su distribución vertical sobre su hospedero. Se registró un total de 33 especies epífitas, de las cuales 29 se asociaron, mínimo con un taxón de hospedero. Los taxa arbóreos fueron 24; en 16 de ellas se observó la presencia de al menos una epífita. El número de especies epífitas fue mayor en altitudes menores. De acuerdo con la cantidad de individuos en cada una de las secciones del forofito, se registró una preferencia hacia las zonas III y IV, correspondientes a las partes basal y media de las ramas. La filogenia no estuvo relacionada con la distribución de los taxones en las zonas de los hospederos ni con las altitudes estudiadas. El total de taxa es pequeño, en relación a estudios similares llevados a cabo en latitudes más sureñas, quizá como resultado de un decremento general en la riqueza de especies, asociado con la distancia respecto al Ecuador.
Palabras clave: Análisis filogenético; bosque de niebla; forofitos; Reserva de la Biósfera El Cielo; riqueza de especies; zonificación de hábitat
Introduction
The mountain mesophylic forest or cloud forest (BMM, for its acronym in Spanish) is one of the ecosystems that hosts the greatest diversity of flora and fauna species, although it covers less than 1 % of the national territory (Challenger, 1998). It is estimated that 2 500 to 3 000 taxa of vascular plants live exclusively and preferably in this type of vegetation (Rzedowski, 1993). Its distribution coincides with a temperate climate of high humidity, in which precipitation and fog occur continuously or very frequently, which thus gives its name of cloud forest or humid mountain forest (Rzedowski, 1978; Villaseñor, 2010).
An important component of the BMM are the epiphytic plants, which grow on other plants without obtaining their nutrients, so the host or phorophyte only has the support function (Benzing, 1990). Epiphytes contribute significantly to the biodiversity of the tropics, as they constitute up to 40 % of the flora of a tropical zone, and 10 % of all vascular plants in the world (Gentry and Dodson, 1987). They provide habitat and food to many invertebrates and birds (Barthlott et al., 2001) and actively participate in the dynamics of nutrients and water retained in the canopy, which are reincorporated into the environment through water runoff or by the fall and death of epiphytes (Nadkarni and Matelson, 1991).
Even though 800 species of epiphytes have been recognized in the BMM of Mexico, only in some the floristic composition of these plants and their contribution to the total flora is known in detail (Sánchez-Ramos and Dirzo, 2014). This is mainly due to the difficulty involved in sites, usually inaccessible (Ingram and Lowman, 1995), although in recent years new methods of promotion have improved this situation.
In any case, the knowledge of the distribution of epiphytes based on biotic and abiotic conditions is important to understand the patterns of diversity in different ecosystems, especially if they are threatened, as is the case of BMM (Gentry and Dodson, 1987). In the present work, the composition of epiphytic species was determined in one of the most boreal distribution points of the cloud forest. The possible association between epiphytes and phorophytes is emphasized, as well as the preference of the former to grow in sections of the trunk or canopy of their host tree and the relationship between the distribution of species and their phylogeny.
Materials and Methods
Study Area
El Cielo Biosphere Reserve is located in the southwest of the state of Tamaulipas, in the foothills of the Sierra Madre Oriental. It is limited by the parallels 23°12'and 23°03'N and the meridian 99°18' W (Puig and Bracho, 1987). According to Conabio (2010), El Cielo includes the most northern fragments of BMM distribution in the American continent. The largest portion of BMM in Tamaulipas is located in the reserve (approximately 16 142.86 ha), and it also has a high degree of conservation (Sánchez-Ramos et al., 2005).
The altitudinal limits of the mesophilic mountain forest in El Cielo are 800 and 1 500 m, covering an approximate area of 100 km2. The predominant soils are lithosols (black), rendzinas (black) and chromic luvisols (black) (Puig and Bracho, 1987). The semiwarm subhumid climate prevails in most of the mesophilic forest; rain precipitations are greater than 2 000 mm annually, and they are located in heights between 700 and 1 400 masl (Casas and Requena, 2005).
In places without disturbance there is a dense forest in the tree stratum, which in turn differs according to the height in four sub-strata. The canopy has a height between 18 and 30 m; is composed of Liquidambar styraciflua L., Clethra pringlei S. Watson, Quercus germana Schltdl. & Cham. and Quercus sartorii Liebm., in addition to Fagus mexicana Martínez in some areas. Other species are distributed between 12 and 18 m, such as Carya ovata (Mill.) K. Koch, Magnolia tamaulipana A. Vázquez, Ostrya virginiana (Mill.) K. Koch, Podocarpus reichei J. Buchholz & N.E. Gray and Tilia houghi Rose. In a third substratum (8 to 12 m), Meliosma oaxacana Standl. and Turpinia occidentalis (Sw.) G. Don grow; and in the fourth, Eugenia capuli (Schltdl. & Cham.) Hook. & Arn., Rapanea myricoides (Schltdl.) Lundell and Ternstroemia sylvatica Schltdl. & Cham. (Puig et al., 1983).
Selection of sites and sampling
Field work began in April 2011, with monthly trips of one week, in a period of six months. Sites located in the altitudes of 800, 1 000, 1 300 and 1 500 masl were selected, according to the distribution of the mountain mesophylic forest in the Reserve (Puig and Bracho, 1987).
Based on the methodology proposed by Gradstein et al. (2003), in each altitudinal elevation five mature trees were chosen, with a minimum distance of 25 m between each individual. The largest ones were chosen, because they are presumed to be the oldest, and therefore, subject to greater colonization by epiphytes (Gradstein et al., 1996).
Around each mature tree a quadrant of 20 × 20 m was delimited, in order to sample the neighboring specimens with the presence of epiphytes. The location of each sampled arboreal individual was recorded by means of a Garmin e-trex GPS and its diameter at breast height (DAP) was measured with a 10 m diameter tape (Forestry suppliers); and its height, with a clinician PM5 / 360PC Suunto.
For the registration of the epiphytic species, the sampled tree (both mature and those of the quadrants) was subdivided into the five zones proposed by Johansson (1974) (Figure 1). The epiphytes were considered by arboreal individual, from the base of the trunk to the outer branches.
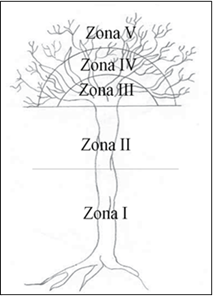
Source: Bøgh, 1992.
Zona = Zone Zone 1 = 0.22-3 m from the ground; Zone 2 = 3 m-first main branches; Zone 3 = Large branches; Zone 4 = Middle and thin branches; Zone 5 = The thinnest branches
Figure 1 Zoning of the host tree (Johansson, 1974).
In order to have access to the canopy, climbing equipment, harness, hammers, elevators, stirrups, carabiners and descent device were used (Benzing, 1990). In addition, observations were made from the ground, with the help of binoculars (Eagle Optics Ranger ED). A total of 188 trees of 24 species, 20 genera and 17 families were sampled: 40 at an elevation of 1 500 m, 50 to 1 300 masl, 46 to 1 000 masl and 52 to 800 masl.
Species of vascular epiphytes, both adult and juvenile specimens, were included in the sampling. They were taken as vegetative units: individuals, rosettes and leaves, (Hietz and Hietz-Seifert, 1995). For the first category, the Peperomia genus was considered; for the second, most of the bromeliaceae taxa; and for the third, the ferns and most of the genera of orchids that form large aggregates, in which it is not reliable to distinguish one individual from another. Copies of all the taxa were collected, which were herborized, identified and deposited in the herbarium of the Facultad de Ciencias Forestales de la Universidad Autónoma de Nuevo León (School of Forest Science of the Autonomous University of Nuevo León).
Statistical Analysis
The diversity of the epiphytes was calculated with the Shannon index. Phenograms were elaborated to know the degree of similarity that exists between the zones of the tree, according to the epiphytic species contained. These also provide the value of the Sørensen similarity index. A χ2 test was applied to determine the preference of the epiphytes towards the hosts, as well as to know the association between the epiphytes and each one of the zones of the tree proposed by Johansson (1974). The phylogenetic distance between the species, measured in millions of years from a common ancestor was detected with Timetree (Hedges et al., 2006), and the differences between phylogenetic distances were analyzed with an ANOVA.
Results
Abundance and species richness
Ten families, 23 genera and 33 species of epiphytes were identified (Table 1). The total area sampled was 0.8 ha (8 000 m2). The Orchidaceae family represented 43 % of the total taxa, followed by Polypodiaceae (15 %) and Bromeliaceae (12 %).
Table 1 Floristic inventory of epiphytes identified in the mesophilic mountain forest of El Cielo Biosphere Reserve.
| Angiosperms | |||
|---|---|---|---|
| Famiy | Species | Altitude (masl) | Zones of the phorophyte |
| Agavaceae | Agave celsii Hook. | 800, 1 500 | 1-3 |
| Begoniaceae | Begonia sp. | 1 300 | 1 |
| Bromeliaceae | Tillandsia bartramii Elliott | 800-1 500 | 2-5 |
| Tillandsia deppeana Steud. | 800-1 500 | 1-5 | |
| Tillandsia utriculata L. | 800, 1 000, 1 500 | 2-5 | |
| Tillandsia sp. | 1 300, 1 500 | 1,3,4 | |
| Cactaceae | Rhipsalis baccifera (Sol.) Stearn | 800 | 1-4 |
| Selenicereus sp. | 800 | 1,2 | |
| Crassulaceae | Sedum sp. | 1 300 | 3 |
| Orchidaceae | Arpophyllum laxiflorum Pfitzer | 1 500 | 4 |
| Catasetum integerrimum Hook. | 800, 1 000 | 1-4 | |
| Desconocida 1 | 1 500 | 2,4 | |
| Desconocida 2 | 1 500 | 2 | |
| Encyclia alata (Bateman) Schltr. | 800, 1 000 | 1-3 | |
| Encyclia parviflora (Regel) Withner | 800 | 1-4 | |
| Isochilus unilateralis B.L. Rob. | 800-1 500 | 1-5 | |
| Laelia anceps Lindl. | 1 300 | 3 | |
| Oncidium sphacelatum Lindl. | 800 | 1-4 | |
| Prosthechea cochleata (L.) W.E. Higgins | 800 | 1-4 | |
| Prosthechea lívida (Lindl.) W.E. Higgins | 800 | 2-4 | |
| Stanhopea tigrina Bateman ex Lindl. | 800-1 500 | 1-4 | |
| Trichocentrum ascendens (Lindl.) M.W. Chase & N.H. Williams | 800 | 1-4 | |
| Trichocentrum cosymbephorum (C. Morren) R. Jiménez & Carnevali | 800 | 1-3 | |
| Piperaceae | Peperomia quadrifolia (L.) Kunth | 800-1 500 | 1-4 |
| Peperomia sp. | 800 | 1,2 | |
| Pteridophytes | |||
| Lycopodiaceae | Huperzia dichotoma (Jacq.) Trevis. | 800-1 500 | 1-4 |
| Aspleniaceae | Asplenium cuspidatum Lam. | 800, 1 300-1 500 | 1-5 |
| Asplenium sp. | 1 300 | 2-5 | |
| Polypodiaceae | Campyloneurum angustifolium (Sw.) Fée | 800-1 500 | 1-5 |
| Pecluma alfredii (Rosenst.) M.G. Price | 800, 1 000 | 1-4 | |
| Pleopeltis polylepis (Roem. ex Kunze) T. Moore | 800-1 500 | 1-5 | |
| Polypodium polypodioides (L.) Watt | 800-1 500 | 1-5 | |
| Polypodium sp. | 1 300 | 1-4 | |
115 069 vegetative units were recorded. Six species have the highest percentage of abundance of individuals: Pleopeltis polylepis (Roem ex Kunze) T. Moore (Polypodiaceae, 40.65 %), Isochilus unilateralis B.L. Rob. (Orchidaceae, 16.52 %); Polypodium polypodioides (L.) Watt (Polypodiaceae, 9.74 %); Asplenium cuspidatum Lam. (Aspleniaceae, 9.22 %), Tillandsia deppeana Steud. (Bromeliaceae, 4.87 %); and Tillandsia bartramii Elliott (Bromeliaceae, 4.81 %). The rest presented a low abundance, lower than 1 % (Figure 2).
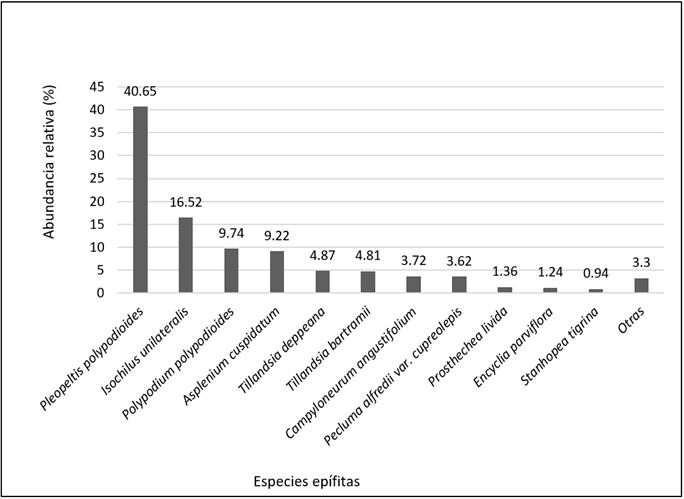
Especies epífitas = Epiphyte species; Abundancia relative = Relative abundance; Otras = Others.
Figure 2 Relative abundance (% of individuals) of the epiphyte species.
Quercus germana was the phorophyte with the highest percentage of epiphytes (38.08 %), followed by Q. sartorii (21.61 %) and Clethra pringlei (13.31 %)
Phorophyte -epiphyte association
Table 2 shows the results of the χ2 test. Epiphytes associated with phorophytes were not phylogenetically more distant from phorophytes than non-associated epiphytes (F = 0.51, P = 0.43).
Table 2 χ2 values that show association between the most frequent epiphytes and the phorophytes.
| Epífita/Forofito | Pleopeltis polylepis | Isochilus unilateralis | Polypodium polypodioides | Asplenium cuspidatum | Tillandsia bartramii | Campyloneurum angustifolium | Pecluma alfredii var. cupreolepis | Sedum sp. | Arpophyllum laxiflorum |
|---|---|---|---|---|---|---|---|---|---|
| Quercus germana | N.S. | 20.1* | 6.82* | N.S. | 5.36* | 25* | N.S. | N.S. | N.S. |
| Quercus sartorii | N.S. | N.S. | N.S. | N.S. | 6.99* | N.S. | N.S. | N.S. | N.S. |
| Clethra pringlei | N.S. | N.S. | N.S. | N.S. | N.S. | 4.95* | N.S. | N.S. | N.S. |
| Quercus canbyi | N.S. | N.S. | N.S. | 5.72* | N.S. | N.S. | N.S. | N.S. | N.S. |
| Sapindus saponaria | N.S. | N.S. | N.S. | N.S. | N.S. | N.S. | 6.78* | N.S. | N.S. |
| Liquidambar styraciflua | 7.9* | N.S. | 5.21* | N.S. | N.S. | N.S. | N.S. | N.S. | N.S. |
| Decatropis bicolor | N.S. | N.S. | N.S. | N.S. | N.S. | 0.36 | 4.8* | N.S. | N.S. |
| Turpinia occidentalis | 2.86 | N.S. | N.S. | 2.94 | N.S. | 6.35* | N.S. | N.S. | N.S. |
| Rapanea myricoides | N.S. | N.S. | N.S. | N.S. | N.S. | N.S. | N.S. | 14.3 | 18.8 |
| Quercus affinis | N.S. | N.S. | N.S. | N.S. | N.S. | N.S. | N.S. | N.S. | 6.2 |
Tabulated χ2= 3.84, p< 0.05); N.S. = Non- significant; * = Significant differences (p<0.05).
Similarity between phorophytes by epiphytic load
The phenogram to analyze the similarity between tree species, with respect to the epiphytes they harbor (Figure 3) shows that the species with the greatest similarity were: 1) Nectandra sanguinea Rol. ex Rottb., Meliosma oaxacana and Abies vejarii Martínez (Index of Similarity = 1.0); 2) Quercus germana and Quercus sartorii (Similarity Index = 0.945); 3) Podocarpus reichei and Magnolia tamaulipana (Similarity Index = 0.880); and 4) Turpinia occidentalis and Rapanea myricoides (Similarity Index = 0.875).
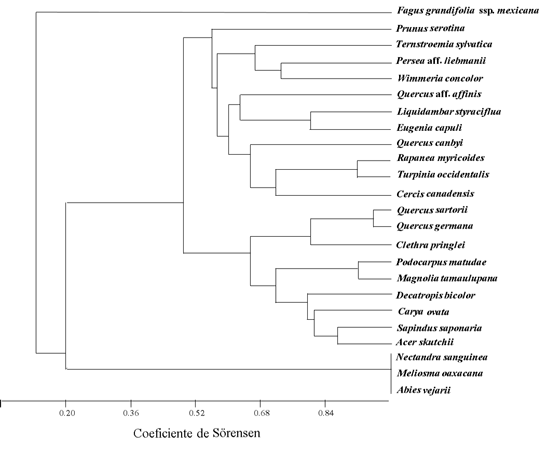
Proportion of species by families in altitudinal elevations
At 1 500 masl, 15 species and six families were identified, Orchidaceae had the highest proportion of species (33 %), followed by Bromeliaceae (26 %) and Polypodiaceae (20 %). At 1 300 masl, 16 species and seven families occurred, including Polypodiaceae (29 %), Orchidaceae and Bromeliaceae (18 % and 17 %, respectively). Two families are restricted to this altitudinal floor: Crassulaceae and Begoniaceae, although their representation within the quadrants is of few individuals. At the level of 1 000 masl, 12 species and five families were recorded, Orchidaceae was the best represented (34 %), followed by Bromeliaceae and Polypodiaceae, both with 25 %. At 800 masl there were 24 species and eight families, of which Orchidaceae had the highest proportion of species (41.67 % of the total), Polypodiaceae (16.67 %) and Bromeliaceae (12.50 %). The highest number of epiphytes was obtained at 800 masl, which coincides with the Shannon Index by altitudinal elevation (Table 3). The phylogenetic distance between epiphytic species was similar within and between altitudes (F = 1.41, P = 0.18).
Vertical distribution of epiphytes
The vertical distribution of the epiphytes in each of the sections of the phorophyte shows its marked preference towards zones III and IV of the host, according to the Johansson classification (Figure 4; χ2 = 31 700.00, p <0.001), sites that correspond to the basal and middle part of the branches; there are more epiphytes than would be expected in a random distribution.
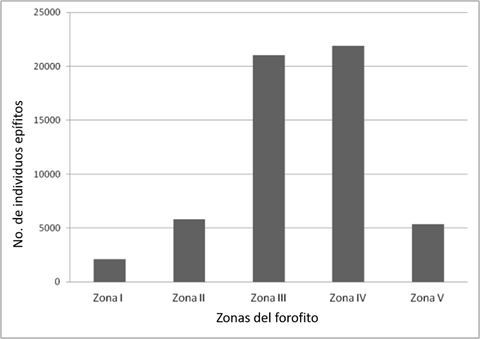
Zonas del forofito = Zones of the phorophyte; No. de individuos epífitos = Number of epiphytic individuals.
Figure 4 Number of epiphytic individuals in each of the areas of the phorophyte.
There were differences in the distribution per zones of all species, whose calculated χ2 values were higher than those of the tabulated χ2 (χ2= 9.48, p= 0.05). This means that the epiphytes, even those that are presente in the five zones of the phorophyte, are more common in one or two of them and it is there where the greatest abundance was recorded.
Discussion
Abundance and species richness
The predominance of the families Orchidaceae, Polypodiaceae and Bromeliaceae coincides with the classification made by Gentry and Dodson (1987) of these taxonomic groups, with respect to the total number of epiphytic species. It is worth mentioning that Orchidaceae, although it has the highest number of taxa, it has a low abundance. Isochilus unilateralis (Orchidaceae) had the greatest value in the fragment of cloud forest studied, perhaps because of the small space it occupies. The pattern of distribution of this family agrees with what is cited in the literature, regarding that orchids have a high diversity of species, although the main characteristic of many of them is their scarcity of individuals (Nieder et al., 1999; 2006).
It stands out for its abundance of Pleopeltis polylepis, which is explained by its tendency to form large conglomerates, which sometimes cover almost completely the branches of the phorophytes, without allowing the growth of other species in the vicinity.
Epiphyte-phorophyte association
The main function of the phorophyte is to provide support to the epiphytes. If its function were restricted to it, the distribution of these plants would be general, regardless of the arboreal species that supports it. Some correlate with the tree taxa by the χ2 test. That is, there are characteristics of the phorophytes that affect the distribution and abundance of epiphytes (Callawell et al., 2002, Krömer and Gradstein, 2003). This correlation was determined in the most abundant species, as in the most scarce; examples of the first are: Isochilus unilateralis, Campyloneurum angustifolium (Sw.) Fée, Polypodium polypodioides and Tillandsia bartramii; and of the latter: Sedum sp., Laelia anceps Lindl., Begonia sp., Arpophyllum laxiflorum Pfitzer and Piper sp. However, the rarity of these epiphytic species may also be the factor that affects the values of χ2, so that they suggest a correlation with the phorophyte. Increasing the number of collections would help to expand the last point raised.
Elements such as the size and shape of the tree determine the abundance and richness of epiphytic species. The predominance of Quercus germana as a preferred phorophyte, it is due to its abundance and physiognomy, since they have a very branched shape and a greater basal area, as recorded by Rosa-Manzano et al. (2017) in a pine-ecino forest in CBR. In addition, the almost horizontal orientation of its branches propitiates the accumulation of organic matter and the prolonged conservation of environmental humidity, which is ideal for the establishment of many epiphytes (Puig, 1993).
Other tree species very common in the mountain mesophilic forest, such as Liquidambar styraciflua, contain few plants of this type, although they have rough bark. However, its shaft is almost straight, its ramifications are thin and its canopy cover is small, so it offers little fixing surface for epiphytes (Puig, 1993).
Vertical distribution
In several works it is mentioned that epiphytes tend to be more abundant in zones III and IV of the phorophytes (Nieder et al., 1999; Krömer et al., 2007), as observed in the present study. According to Nieder et al. (1999), height preferences correspond to adaptations of physiological and mechanical type. Thus, for example, the genus Campyloneurum is more abundant in the first two zones of the phorophyte, which reflects its adaptation to a gloomy environment. In as much, Pleopeltis is distributed in the five areas of the phorophyte, although it is more abundant in zones III and IV. Also in those sites a large part of the orchids is concentrated, with the exception of Isochilus unilateralis, which can be found in zone V, but in smaller numbers, mainly due to the high insolation and the rapid desiccation prevailing in that area.
Comparison with similar studies in neotropical mountain forests
The total of identified taxa is lower than that documented in studies carried out in another mountain mesophilic forest. Martínez et al. (2006) recorded 44 species in an area of 0.04 ha in Chiapas; Hietz and Hietz-Seifert (1995) cite 53 at an altitude of 1 439 masl, within an area of 0.06 ha, with a total of 134 taxa for all their sampling sites. At present, 33 species are recorded in 0.6 ha.
In more southern latitudes, such as Ecuador and Bolivia, the number of species increases, notably (Krömer and Gradstein, 2003). The high number and diversity in the equatorial zones is explained by the multiple niches and microclimates derived from orography (Küper et al., 2004). The lower number of taxa can be explained by the fact that as the latitudinal gradient increases, the diversity of epiphytes decreases (Gentry and Dodson, 1987).
Given that the El Cielo Biosphere Reserve is one of the sites that mark the boreal distribution limit of the mesophilic forest in Mexico (Conabio, 2010), it is understood the low number of species in relation to similar forests, but located in more latitudes. to the south.
Conclusions
There were 33 epiphytic species, which belong to 23 genera and 10 families of vascular plants, with a total of 115 069 individuals. There was an association between some taxa of epiphytes and their phorophytes; they are also more abundant in the basal and intermediate part of them. Phylogeny does not explain the distribution of epiphytes.
The mountain mesophilic forest of the El Cielo Biosphere Reserve shows a pattern of epiphytic distribution similar to that of similar environments in other localities.
Acknowledgements
This work began as the Master's thesis of Eng. Noé Hervé García Balcázar, who unfortunately died before obtaining his Master's degree. The article is dedicated to his memory. The study had partial support from PAICYT (UANL) and Conacyt (CB-2015-01 255453).
REFERENCES
Barthlott, W., V. Schmit-Neuerburg, J. Nieder and S. Engwald. 2001. Diversity and abundance of vascular epiphytes: a comparison of secondary vegetation and primary montane rain forest in the Venezuelan Andes. Plant Ecology 152: 145-156. [ Links ]
Benzing, D. H. 1990. Vascular epiphytes. Cambridge University Press. Cambridge, UK. 280 p. [ Links ]
Bøgh, A. 1992. Composition and distribution of the vascular epiphyte flora of an Ecuadorian montane rain forest. Selbyana 13: 25-34. [ Links ]
Callaway, R. M., K. Reinhart, G. W. Moore and S. Pennings. 2002. Epiphyte host preferences and host traits: mechanisms for species-specific interactions. Oecologia 132(2): 221-230. [ Links ]
Casas G., S. L. y G. N. Requena L. 2005. Generalidades geográficas. In: Sánchez-Ramos, G., P. Reyes-Castillo y R. Dirzo (eds.). Historia natural de la Reserva de la Biósfera El Cielo, Tamaulipas, México. Universidad Autónoma de Tamaulipas. Ciudad Victoria, Tamps., México. 732 p. [ Links ]
Challenger, A. 1998. Utilización y Conservación de los Ecosistemas Terrestres de México. Pasado, Presente y Futuro. Conabio. México, D.F., México. 847 p. [ Links ]
Comisión Nacional para el Conocimiento y Uso de la Biodiversidad (Conabio). 2010. El Bosque Mesófilo de Montaña en México: Amenazas y Oportunidades para su Conservación y Manejo Sostenible. Conabio. México D.F., México. 197 p. [ Links ]
de la Rosa-Manzano, E., A. Guerra-Pérez, G. Mendieta-Leiva, A. Mora-Olivo, J. G. Martínez-Ávalos and L.U. Arellano-Méndez. 2017. Vascular epiphyte diversity in two forest types of the “El Cielo” Biosphere Reserve, Mexico. Botany 95: 599-610. [ Links ]
Gentry, A. H. and C. H. Dodson. 1987. Diversity and biogeography of neotropical vascular epiphytes. Annals of the Missouri Botanical Garden 74: 205-233. [ Links ]
Gradstein, S. R., P. Hietz, R. Lücking, A. Lücking, H. J. Sipman, H. F. M. Vester, J. Wolf and E. Gardette. 1996. How to sample the epiphytic diversity in tropical rain forests. Ecotropica 2: 59-72. [ Links ]
Gradstein, S. R., N. M. Nadkarni, T. Krömer, I. Holz and N. Nöske. 2003. A protocol for rapid and representative sampling of vascular and non-vascular epiphyte diversity of tropical rain forests. Selbyana 24(1): 105-111. [ Links ]
Hedges, S., J. Dudley and S. Kumar. 2006. TimeTree: a public knowledge-base of divergence times among organisms. Bioinformatics 22: 2971-2972. [ Links ]
Hietz, P. and U. Hietz-Seifert. 1995. Composition and ecology of vascular epiphyte communities along an altitudinal gradient in central Veracruz, México. Journal of Vegetation Science 6: 487-498. [ Links ]
Ingram, S. W. and M. D. Lowman. 1995. The collection and preservation of plant material from the tropical forest canopy. In: Lowman, M. D. and N. M. Nadkarni (eds.). Forest Canopies. Academic Press. San Diego, CA USA. pp. 587-603. [ Links ]
Johansson, D. 1974. Ecology of vascular epiphytes in West African rain forests. Acta Phytogeografica Suecica 59: 1-129. [ Links ]
Krömer, T. and S.R. Gradstein. 2003. Species richness of vascular epiphytes in two primary forest and fallows in the Bolivian Andes. Selbyana 24: 190-195. [ Links ]
Krömer, T., M. Kessler and S. R. Gradstein. 2007. Vertical stratification of vascular epiphytes in submontane and montane forest of the Bolivian Andes: the importance of the understory. Plant Ecology 189: 261-278. [ Links ]
Küper W., H. Kreft, J. Nieder, N. Köster and W. Barthlott. 2004. Large-scale diversity patterns of vascular epiphytes in Neotropical montane rain forests. Journal of Biogeography 31:1477-1487. [ Links ]
Martínez M., N., M. A. Pérez F. y A. Flores P. 2006. Estratificación vertical y preferencia de hospedero de las epífitas vasculares de un bosque nublado de Chiapas, México. Revista de Biología Tropical 56(4): 2069-2086. [ Links ]
Nadkarni, N. M. and T. J. Matelson. 1991. Fine litter dynamics within the tree Canopo of a tropical cloud forest. Ecology 72: 2071-2082. [ Links ]
Nieder, J., S. Engwald and W. Barthlott. 1999. Patterns of Neotropical epiphyte diversity. Selbyana 20: 66-75. [ Links ]
Puig, H. 1993. Árboles y arbustos del bosque mesófilo de montaña de la reserva El Cielo, Tamaulipas, México. Instituto de Ecología A.C. Xalapa, Ver., México. 84 p. [ Links ]
Puig, H., R. Bracho y V. Sosa. 1983. Composición florística y estructura del bosque mesófilo en Gómez Farías, Tamaulipas. México. Biótica 8(4): 339-359. [ Links ]
Puig, H. y R. Bracho . 1987. El bosque mesófilo de montaña de Tamaulipas. Primera edición. Instituto de Ecología, A.C. México, D.F., México. 176 p. [ Links ]
Rzedowski, J. 1978. Vegetación de México. Limusa. México, D. F., México. 432 p. [ Links ]
Rzedowski, J. 1993. Diversity and origins of the fanerogamic flora of Mexico. In: Ramamoorthy, T. P., R. Bye, A. Lot and J. Fa (eds.). Biological diversity of Mexico: Origins and distribution. Oxford University Press. New York, NY USA. pp. 139-144. [ Links ]
Sánchez-Ramos, G., P. Reyes-Castillo y R. Dirzo (eds). 2005. Historia natural de la Reserva de la Biosfera El Cielo, Tamaulipas, México. Universidad Autónoma de Tamaulipas. Hong Kong, China. 693 p. [ Links ]
Sánchez-Ramos y R. Dirzo. 2014. El bosque mesófilo de montaña: un ecosistema prioritario y amenazado. En: Gual-Díaz, M. y A. Rendón-Correa (comps.). Bosques mesófilos de montaña de México: diversidad, ecología y manejo. Comisión Nacional para el Conocimiento y Uso de la Biodiversidad. México D.F., México. pp. 109-139. [ Links ]
Valverde R., I. 2006. Crecimiento de Barkeria lindleyana (Orchidaceae) en un hábitat suburbano de Costa Rica. Lankesteriana 6(2): 33-41. [ Links ]
Villaseñor, J. L. 2010. El Bosque Húmedo de Montaña en México y sus plantas vasculares: Catálogo Florístico-Taxonómico. Comisión Nacional para el Conocimiento y Uso de la Biodiversidad y Universidad Nacional Autónoma de México, México D.F., México. 40 p. [ Links ]
Received: February 13, 2018; Accepted: September 12, 2018
 Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
