











 text in
text in  English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkIntroducción
Las plantaciones de Pinus caribaea Morelet var. caribaea W. H. Barret et Golfari son la superficie que mejor representa el patrimonio forestal en la provincia de Pinar del Río, donde el manejo y el aprovechamiento maderable inciden sobre la biodiversidad; las aves, en particular, son el grupo de vertebrados más numeroso tanto en los bosques naturales como en las plantaciones.
Los estudios faunísticos en el Parque Nacional Viñales han sido pocos con respecto a la fauna y su relación con la vegetación; no obstante, desde el siglo XIX, este lugar ha llamado la atención de los naturalistas foráneos y nacionales por constituir un sitio de gran riqueza y variedad de especies (Corvea et al., 2014).
En Cuba se tienen registradas 371 especies silvestres, que pertenecen a 208 géneros, 63 familias y 21 órdenes. Los taxa terrestres suman 217; 69 se asocian con aguas dulces y 83 con aguas marinas; de ellas, 28 son endémicas, las que constituyen 13.1 % de la avifauna cubana (Garrido y Kirkconnell, 2011).
Varios investigadores se han dedicado a investigar las comunidades de aves que habitan los ecosistemas de pinar en diferentes regiones: Vicente (1991) estudió la estructura de 10 sistemas avifauna - vegetación de tres pinares; López y Moro (1997) se enfocaron en las aves de plantaciones de Pinus halepensis Miller en el sureste de España, en relación con la composición y estructura de la vegetación; Canterbury et al. (2000) evaluaron el hábitat asociado a las aves y desarrollaron indicadores ambientales a nivel de comunidad, en 197 cuadrantes en bosques de pino de Georgia a Virginia (Estados Unidos de América), a través de un gradiente de perturbación antropogénica.
A pesar de que P. caribaea var. caribaea conforma una buena parte de la vegetación del Caribe, es poco lo que se ha trabajado en relación a su avifauna (O'Brien, 2005). Un trabajo pionero es el de Cruz (1988), quien estudió el uso del recurso para las especies de aves en una plantación de Pinus caribaea Morelet en Puerto Rico, y recomendó para la mejora de sus hábitats el mantenimiento de un sotobosque diverso y de los árboles nativos.
En este contexto, en Cuba se han llevado a cabo varios trabajos en los pinares. Huerta et al. (1984) realizaron 13 conteos de dichos animales con el fin de determinar la abundancia de cada taxon y su diversidad en la asociación forestal de sabanas blancas en la Isla de la Juventud.
García et al. (1989) en 24 ocasiones cuantificaron las existencias de aves entre el 7 y el 15 de marzo de 1988, en la Zoilita, Sierra de Cristal. La abundancia relativa para bosques de pinos fue de 54.3 individuos ha-1, mientras que en sitios ocupados por vegetación de alta variabilidad ecológica se observaron 70 ejemplares ha-1. Hernández et al. (1998) hicieron un estudio de la estructura de las comunidades de aves que habitan en 216 ha de bosque de P. caribaea, de 25 años de edad, en el que aplicaron el método de Itinerario de Censo. Detectaron 28 especies, 22 residentes permanentes (78.5 %) y seis migratorias (21.4 %); las agruparon en 17 gremios; de las especies residentes permanentes, cinco son endémicas (22.7 %) y nueve consideradas endémicas a nivel subespecífico (40.9 %).
González et al. (1999) estudiaron las comunidades de aves terrestres en 34 áreas de muestreo de 10 localidades de Cuba; para ello utilizaron los métodos de Transecto Lineal, Parcela de Conteo y captura con redes ornitológicas. Determinaron la estructura y composición de las formaciones vegetales mediante el método de parcela de vegetación; sus resultados indican que las variables estructurales que más influyeron en la ordenación de los hábitats y en las poblaciones de las aves son la cobertura del dosel, la cobertura del suelo y el volumen del follaje.
Mereck (2004) realizó una investigación en cinco ecosistemas (Bosque de Galería, Semideciduo de Mogote, Xerófilo de Mogote, Pinar y Encinar) en el Valle de San Andrés, perteneciente a la EFI La Palma; se usó el método de Recuento por Punto sin Estimación de Distancia propuesto por Wunderle (1994) para las aves del Caribe. En los pinares se identificó un total de 50 especies de estos organismos, incluidas en 10 órdenes y 63 familias; ocho son endémicas a nivel específico y 13 a nivel subespecífico.
Peraza y Berovides (2007) realizaron un trabajo sobre la dinámica espacio-temporal de la abundancia, diversidad y usos de recursos (sustratos y estratos de la vegetación) de dos ensambles de aves en pinares, localizados en la Reserva Florística Manejada San Ubaldo Sabanalamar, Pinar del Río.
Hasta el momento no se ha abordado la posible asociación entre las características dasométricas de Pinus caribaea y las comunidades de aves en Cuba. Por lo tanto, el objetivo general del estudio que se describe a continuación fue evaluar la relación de las aves con la altura, la densidad y el área basal, en una plantación de esta especie y, específicamente, determinar grupos funcionales (gremios tróficos), abundancia y diversidad de las aves.
Materiales y Métodos
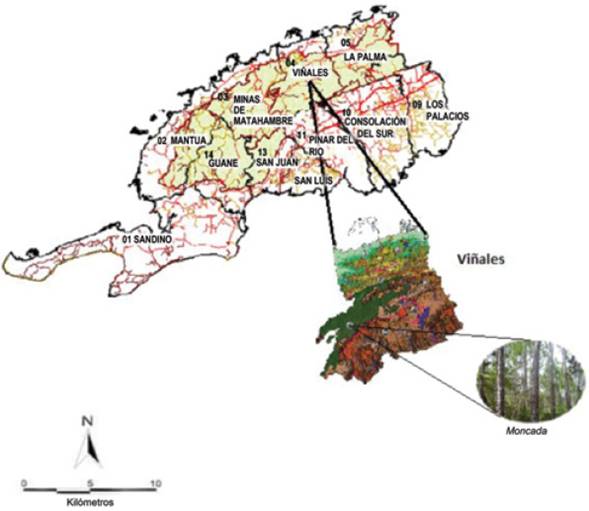
La zona de estudio se ubica en el Parque Nacional Viñales (PNV), municipio Viñales, Provincia Pinar del Río; comprende 1 510 ha, de las cuales 1 120 ha pertenecen a las áreas núcleo y 3 890 ha corresponde a la zona de amortiguamiento, ocupa la porción centro-oriental de la Sierra de los Órganos. Se extiende de NE a SO, con un ancho máximo de 8 km y un mínimo de 2.5 km, y abarca un total de 15 010 ha, sin incluir 920 ha pertenecientes a los cayos adyacentes, lo cual representa 6 % del total de la Provincia.
La investigación se realizó en una plantación de Pinus caribaea, en la localidad del Moncada a 23 km del pueblo de Viñales, en elevaciones donde la vegetación que predomina la conforman los pinos, que se desarrollan sobre suelos muy pobres y erosionados (Figura 1).
El régimen de temperaturas varía de 22 °C a 24 °C (temperatura media en el período invernal) y de 25 °C a 27 °C (temperatura media en el período estival). El acumulado promedio de lluvia es de 1 600 a 1 800 mm año-1. La humedad relativa media anual oscila entre 90 y 95 % (07:00 h) y 65-70 % (13:00 h).
El inventario de aves se llevó a cabo con el método de Parcelas Circulares de radio fijo; se definieron 30 puntos de conteo con diámetro fijo de 25 m a una distancia de 150 m uno de otro y 10 min de observación (Hutto et al., 1986) en las primeras horas de la mañana, entre las 07:30 y 1:30 h, de enero a mayo de 2015. Las aves censadas se agruparon en órdenes y familias, en correspondencia con los criterios de Llanes et al. (2002) y la clasificación por grupos tróficos, de acuerdo a los de Kirkconnell et al. (1992).
El muestreo de la vegetación se hizo en abril y mayo en las mismas parcelas utilizadas para el conteo de aves, para determinar la relación entre las variables de la vegetación y de las aves detectadas.
Los parámetros estructurales de la vegetación que se calcularon fueron:
Densidad de árboles (da = árboles ha-1) = Número de individuos por especie arbórea por estado fenológico
Diámetro de los árboles a la altura de 1.30 m (DAP m) = Todos los árboles se ubicaron por clases de diámetro
Altura del dosel (m) = Promedio de las alturas (m) de los árboles de la parcela
Para comparar la abundancia y riqueza entre parcelas y meses se utilizó la prueba no paramétrica de comparación de rangos de Kruskal Wallis, se determinó la correlación entre la riqueza de especies y las variables dasométricas mediante la matriz de correlación no paramétrica de Rho Spearman. Para identificar la incidencia de las variables dasométricas sobre la composición de las especies ornitológicas se realizó un análisis de correspondencia canónico. Se utilizó el paquete estadístico SPSS versión 15.0.
Resultados y Discusión
El total de especies de aves censadas fue de 41, las cuales se agruparon en nueve órdenes y 16 familias. En el mes de enero se observó el mayor número con 35 y en mayo el menor con 27.
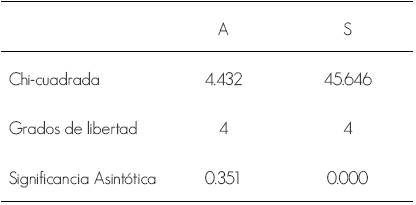
En las comparaciones entre los meses existió diferencia significativa en la riqueza (Cuadro 1), debido que el estudio se llevó a cabo durante los períodos invernal y primaveral comprendido de enero a mayo: los resultados coinciden con los hallazgos de Pérez (2003 y 2007) en investigaciones realizadas en la península de Guanahacabibes; con los de Mereck (2004), en la Empresa Forestal Integral (EFI) La Palma, y con los de Alonso (2009) en la EFI Minas de Matahambre.
Cuadro 1 Resultado de la Prueba de Kruskal-Wallis para la comparación entre los meses, abundancia (A) y riqueza (S).
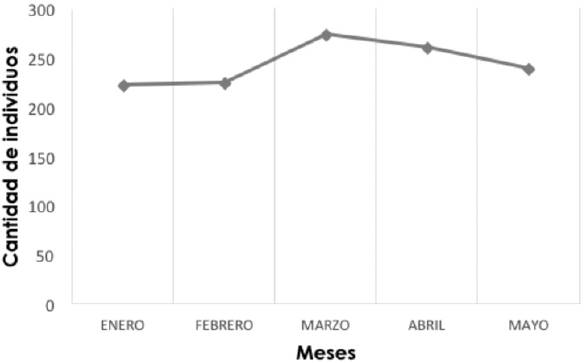
El menor avistamiento se realizó en los meses de enero y febrero con 222 y 224 individuos, respectivamente, y los más altos en marzo y abril con 273 y 261, cuando se observó a muchas de las especies de aves en el cortejo y apareamiento; en mayo se reconocieron varios nidos en el área (Figura 2).
El orden Passeriformes reúne a 50 % del total de las familias presentes, de las cuales las mejor representadas fueron Parulidae con 19.5 %, Columbidae con 14.6 % y Tyranidae con 9.7 %. Los valores concuerdan con los obtenidos en los estudios de Pérez et al. (2003); Mereck (2004); Toledo (2004); Pérez (2007); Peraza (2008) y Alonso (2009).
Las especies se agruparon en 21 gremios tróficos, lo que suma 62 % de los registrados por Kirkconnell et al. (1992); predomina el insectívoro de follaje por espigueo con 12.1 % y granívoro de suelo con 9.7 %. Acosta y Múgica (1988) citan porcentajes parecidos en ocho formaciones arbóreas del territorio nacional, y Sánchez (2007) en pinares naturales de Pinus tropicalis Morelet (Figura 3).
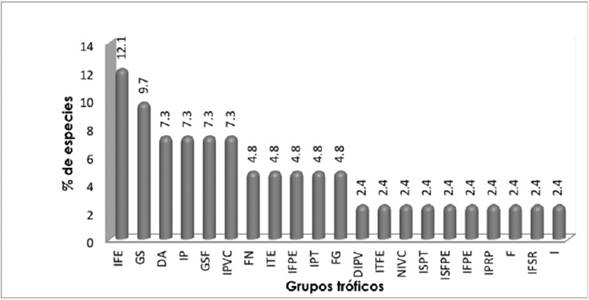
ITE) = Insectívoro de tronco por espigueo; (IFE) = Insectívoro de follaje por espigueo; (ITFE) = Insectívoro de trono y follaje por espigueo; (N-IVC) = Nectarívoroinsectívoro con vuelo cernido; (ISFPE) = Insectívoro de suelo y follaje, con picoteo y espigueo; (I-FP) = Insectívoro-frugívoro de percha; (IPT) = Insectívoro perforador de tronco; (GS) = Granívoro de suelo; (F) = Frugívoro; (GSF) = Granívoro de suelo y follaje; (DIPV) = Depredador de insectos y pequeños vertebrados; (IPRP) = Insectívoro de percha con revoloteo y persecución; (IP) = Insectívoro de percha; (IPVC) = Insectívoro de percha con vuelo colgado; (F-G) = Frugívorogranívoro; (I-FPE) = Insectívoro-frugívoro con picoteo y espigueo; (I ) = Insectívoro; (ISPT) = Insectívoro (I-FSR).
Figura 3 Representación del porcentaje de especies por grupos tróficos presentes en el área de estudio.
En la Figura 4 se observa el comportamiento medio de la abundancia y la riqueza por parcela en el período estudiado; la parcela 1 es la que presenta los valores más grandes, lo que puede explicarse por una menor densidad de pino y la existencia de un mayor número de otros taxa arbóreos como Abarema obovalis (A. Rich.) Barneby & J. W. Grimes con flores, las cuales son llamativos para los insectos, Davilla rugosa Poir; y especies de la familia Melastomataceae que estaban en fructificación.
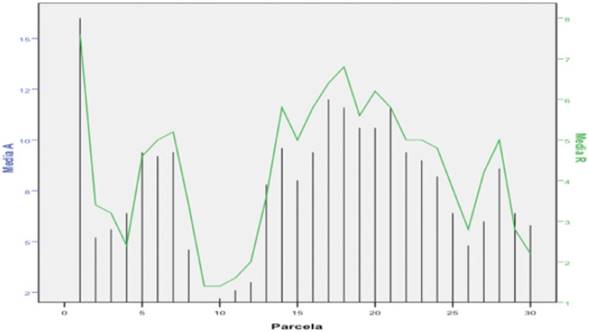
Figura 4 Comportamiento de las medias de la riqueza y la abundancia de especies de aves por parcela.
En las parcelas de la 8 a la 12 se registraron la abundancia y riqueza más bajas, lo que responde a una densidad superior de árboles de menor altura, con un sotobosque herbáceo compuesto, esencialmente, por los helechos Odontosoria sp. y Urechites luteus (L.) Britton, que no aportan frutos y en el que rara vez fueron avistadas las aves.
En contraste, en las parcelas de la 13 a la 23 la abundancia y la riqueza tienen un buen comportamiento, algo semejante a lo ocurrido en la parcela 1, pero en este caso, además de la baja densidadarbórea, con un gran número de especies vegetales en el sotobosque tales como Byrsonima coriacea (Sw.) DC., Quercus cubana A. Rich., Clidemia strigillosa (Sw.) DC., Conostegia xalapensis (Bonpl.) D. Don ex DC., Miconia aevigata (L.) D. Don, Matayba oppositifolia (A. Rich.) Britton., Clusia rosea Jacq. y Gouania poligama (Jacq.) Urb., que les brindan alimentos a las aves.
En los estadísticos de contraste, las comparaciones entre parcelas evidenciaron diferencias significativas de la abundancia y la riqueza, lo que está determinado por la densidad de los árboles, la altura promedio en las parcelas y la abundancia de especies vegetales (Cuadro 2). La información coincide con los registros de Peraza (2008).
Cuadro 2 Resultado de la Prueba de Kruskal-Wallis para la comparación entre las parcelas y abundancia (A) y riqueza (R).
En el Cuadro 3 se advierte que las correlaciones del área basal con la abundancia y la riqueza de aves es baja, pero significativa a un intervalo de confianza de 99 %, lo que está influenciado por la relación matemática entre la altura y el área basal, además de que muchas especies de aves se distribuyen en los estratos superiores del arbolado en diferentes épocas del año, como Tyrannus caudifaciatus d'Orbigny, 1839 y Turdus plumbeus Linnaeus, 1758, a pesar de que muchas veces forrajean en el suelo.
Cuadro 3 Representación de la matriz de correlación no paramétrica de Rho Spearman.
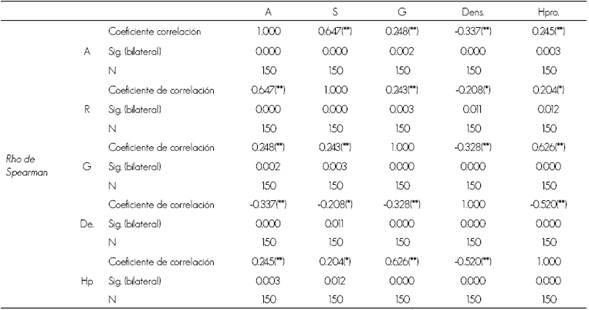
** La correlación es significativa al nivel 0.01 (bilateral); * La correlación es significativa al nivel 0.05 (bilateral); G = Área basal; De = Densidad; Hp = Altura promedio de la parcela; S = Riqueza y A = Abundancia.
Por su parte, la densidad de árboles muestra una correlación baja e inversa con la abundancia y riqueza de especies, lo cual se pudiera explicar por los taxa de aves que necesitan menor densidad de árboles, mayor espacio para efectuar su vuelo y la búsqueda de sus alimentos. La altura promedio tiene una correlación positiva, con la abundancia y riqueza de las aves y está determinada por algunas especies que se encuentran en árboles altos para su alimentación, como son los representantes del gremio trófico insectívoro de tronco y follaje por espigueo y los insectívoros perforadores de tronco.
En la Figura 5 se observa cómo la variable densidad muestra mejor asociación con las aves en el cuadrante 2 con las especies: Mniotilta varia Linnaeus, 1766 (bij trepa), Teretistris fernandinae Lembeye, 1850 (chillina), Cyanerpes cyaneus Linnaeus, 1766 (aparecid), Chlorostilbon ricordii Gervais, 1835 (zunzún), Setophaga palmarum J. F. Gmelin, 1789 (bij común), Tyrannus dominicensis J. F. Gmelin, 1788 (pitirre) y Myiarchus sagrae Gundlach, 1852 (bobito), en particular, las dos últimas; este resultado es atribuible a su forma de vuelo, que demanda más espacio para capturar el recurso alimento, y las otras, lo que necesitan es más densidad para capturar insectos.

Figura 5 Representación del diagrama de ordenación de análisis de correspondencia de las especies de aves con la densidad, altura y área basal en una plantación de Pinus caribaea Morelet.
En el cuadrante 3 en el que se relacionan el área basal y la altura, las aves que se vinculan con estas variables son Spindalis zena Linnaeus, 1758 (cabrero), Setophaga pityophila Gundlach, 1858 (biji pinar), Mniotilta varia (bij trepa), Priotelus temnurus Temminck, 1825 (tococoro), Melanerpes superciliaris Temminck, 1827 (carpi ja) y Xiphidiopicus percussus Temminck, 1826 (carpi ve). Los taxa que se estudiaron y mostraron asociación fueron mayoritariamente insectívoros.
Conclusiones
La composición taxonómica de las comunidades de aves asociadas al área de estudio estuvo compuesta por nueve órdenes, 16 familias y 41 especies. El orden Paseriformes y la familia Parulidae fueron los mejores representados. Los gremios tróficos más abundantes fueron el insectívoro de follaje por espigueo con 12. 1 % y el granívoro de suelo con un 9.7 %. Las aves presentan mayor relación con la densidad y el área basal y menor con la altura promedio del arbolado.
Conflicto de intereses
Los autores declaran no tener conflicto de intereses.
Contribución por autor
Sael Anoi Báez Pérez: trabajo de campo, levantamiento de las parcelas y punto de conteos, monitoreo de las aves, estudio de vegetación, procesamiento de los datos y análisis de los resultados y redacción del documento; Leyanis Pintado Martínez: trabajo de campo, levantamiento de las parcelas y punto de conteos, monitoreo de las aves, estudio de vegetación y caracterización del área de estudio; Fernando Hernández Martínez: trabajo de campo, levantamiento de las parcelas y punto de conteos, monitoreo de las aves, estudio de vegetación, clasificación taxonómica de las especies de aves.

