










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista mexicana de ciencias forestales
Print version ISSN 2007-1132
Rev. mex. de cienc. forestales vol.2 n.6 México Jul./Aug. 2011
Artículos
Impacto de Phellinus pini (Brot.) Bondartsev & Singer en bosques de Pinus greggii Engelm. del estado de Nuevo León
Impact of Phellinus pini (Brot.) Bondartsev & Singer in Pinus greggii Engelm. forests of Nuevo León state
José G. Marmolejo1*, Heriberto Méndez Cortés1 y César Cantú A.1
1 Facultad de Ciencias Forestales. Universidad Autónoma de Nuevo León. *Correo-e: jmarmole@gmail.com
Fecha de recepción: 16 de mayo de 2011
Fecha de aceptación: 4 de agosto de 2011
RESUMEN
Phellinus pini es un hongo ampliamente distribuido en el hemisferio norte, el causante de la pudrición roja anular y el más importante que afecta a los árboles en pie de las pináceas. En años recientes se ha detectado la muerte del muchos ejemplares de Pinus greggii en el municipio de Santiago, N. L., lo que se atribuye a la acción de este organismo, con el consecuente deterioro de la masa arbolada del Parque Nacional Cumbres de Monterrey. Por esto, y debido al valor de los bosques de pino en el Estado, se plantearon como objetivos de estudio la determinación del impacto de Phellinus pini , su asociación con algunas variables silvícolas y la estimación del turno patológico. Se establecieron parcelas de muestreo circulares en varias localidades; se evaluó la presencia de pudrición mediante la toma de muestras por medio del taladro de Pressler y su posterior cultivo en cajas Petri en laboratorio. Se tomaron las variables silvícolas de todos los árboles muestreados para determinar los factores que favorecen la incidencia del agente causal y, por último, el turno patológico. La edad fue la variable de mayor correlación, que se confirma con los individuos de edad avanzada, sobre los cuales se advirtió el mayor daño. La incidencia y severidad fue de 11.66 y 12.3% respectivamente; el turno patológico propuesto es de 50 años.
Palabras clave: Incidencia, Phellinus pini , Pinus greggii, pudrición blanca, turno patológico, variables silvícolas.
ABSTRACT
Phellinus pini is widely distributed in the Northern Hemisphere, causes the red ring rot and is the most important agent that affects softwood trees. In recent years the death of many Pinus greggi examples has been detected in Santiago Municipalitiy, Nuevo Leon State, which has been linked to it, with the resulting tree mass deterioration of the Monterrey Heights National Park. Thus, and due to what forests are worth in the State, the following objectives were proposed for this study: to determine the impact of Phellinus pini, its association with some forestry variables and the pathological rotation age. Circular sampling lots were placed in various locations; rottenness was assessed through samples taken by a Pressler boarer, and its eventual laboratory cultivation in Petri boxes. Forestry variables were taken from all the sampled trees to determine the factors that favor the incidence of this pathogen and, finally, the pathological rotation age. Age was the highest correlation variable, which was confirmed with the eldest examples, over which the most severe damages were observed. Incidence and severity were 11.66 and 12.3%, respectively; the pathological rotation age was 50 years.
Key words: Disease incidence, Phellinus pini, Pinus greggii, white rot, pathological rotation age, forestry variables.
INTRODUCCIÓN
Los bosques de coníferas son frecuentemente atacados por organismos patógenos que se convierten en plagas y causan enfermedades que reducen el rendimiento de la producción de madera en pie. Entre ellos, los hongos inciden sobre la raíz, el fuste, las ramas y las hojas, provocando graves problemas.
Aun cuando muchas especies de ellos están adaptadas a vivir en pinos vivos, Phellinus pini (Brot.) Bondartsev & Singer (syn: Fomes pini (Thore) P. Karst.), está ampliamente distribuido en el hemisferio norte y es el responsable de la pudrición roja anular, y de la de árboles en pie en las pináceas (Haddow, 1938).
Esta especie es muy ubicua y tiene presencia cosmopolita (Blanchette, 1980), aunque Fisher (1994) asegura que no existe en Norteamérica. Se le ha identificado en asociación con más de 100 especies de aciculares (Larsen y Melo, 1996).
P. pini debe su éxito de colonización a su capacidad para degradar y mineralizar la lignina, prefiriendo las traqueidas de otoño por ser las más lignificadas (Blanchette, 1980); también lo hacen con la celulosa y la hemicelulosa en etapas avanzadas (Blanchette et al ., 1987), para lo cual estos hongos poseen un complejo de enzimas oxidasas y peroxidasas que usan para romper esas uniones complejas (Leandro et al., 2003), ocasionando con ello una oxidación de los azúcares debido a la ruptura de estas moléculas (Rodríguez, 1998).
Este tipo de hongos se desarrolla con más frecuencia en árboles viejos (Basham y Morawsky, 1964); sin embargo, se registraron pudriciones en Pinus banksiana Lambert a los diecisiete años de edad (Blanchette, 1982). Esto se explica por la formación de duramen existente en el árbol, que varía según la especie; en algunas se inicia en los primero años, mientras que en otras sólo se produce cuando alcanzan mayor edad o diámetros de cierta envergadura.
Por otra parte, la variación del diámetro del duramen aumenta desde la base del árbol hacia el DAP, para luego decrecer con la altura del fuste, lo que no siempre es posible advertir (Díaz-Vaz, 2003). La degradación del duramen puede ser tanto hacia arriba como hacia abajo de donde fue el punto de entrada del hongo, mientras que la pudrición en la albura es relativamente limitada (Basham, 1975).
García y Montero (1998) hicieron notar la escasez de trabajos sobre los factores que influyen sobre la incidencia de P. pini, y mencionan la edad, la altura, el área basal y el índice de sitio como los más relacionados con este tipo de pudrición.
En México, aunque se reconoce la importancia de P. pini , ningún estudio se ha hecho que detalle los síntomas, signos, impacto y manejo de los rodales afectados. Los hasta ahora conocidos solo se han limitado al conocimiento de su distribución, teniendo registros bibliográficos en los estados de Hidalgo, Morelos, Nuevo León, Valle de México y Veracruz, aunque su distribución pudiera ser más amplia (Bandala et al ., 1993).
En los últimos años se ha detectado la muerte del arbolado de Pinus greggii Engelm. en el municipio de Santiago, N L con los consecuentes graves daños a la masa arbolada, que es muy significativo porque esta es una especie de gran importancia ecológica en el Parque Nacional Cumbres de Monterrey. La causa de la mortalidad se atribuye a este hongo. Es por esto, y debido al valor de los bosques de pino en el estado, que los objetivos del estudio fueron la determinación del impacto de Phellinus pini , su asociación con algunas variables silvícolas y la estimación del turno patológico.
MATERIALES Y MÉTODOS
Para este estudio se delimitaron 120 parcelas circulares siguiendo el método propuesto por García y Montero (1998); que considera un diámetro de la parcela variable, pero que incluya 20 árboles. Cada una de ellas fue georreferenciada con la ayuda del GPS Margellan Triton 400, además se anotó la altura sobre el nivel del mar, exposición, fecha y pendiente con equipo convencional de medición.
De cada árbol se tomó el diámetro a la altura del pecho (DAP), altura e inspección visual de su condición física, y se anotó la presencia de basidiocarpos, puntas rotas o muertas, bifurcadas, ramas muertas, cicatrices y resinaciones.
Dentro de cada parcela se sortearon al azar cinco árboles de cada uno de los cuales se obtuvieron dos muestras con el taladro de incremento (de Pressler Haglöf de 5.15mm/450 mm). Una de ellas fue tomada a 0.3 m y la otra a la altura del pecho (1.3 m); con este material se determinó la edad y el nivel de pudrición.
Se recolectaron los basidiocarpos que estaban creciendo sobre árboles vivos dentro de la parcela y se fotografiaron los signos y los síntomas.
Para tener la certeza de la presencia de P. pini, se analizaron las muestras obtenidas a 0.3 m, las cuales fueron debidamente etiquetadas y llevadas al Laboratorio de Patología y Micología Forestal de la Facultad de Ciencias Forestales de la UANL para su procesamiento. Por cada muestra (viruta) se extrajeron seis submuestras a lo largo de ésta y se aislaron en dos cajas Petri con extracto de malta agar al 2 %. Las cajas se mantuvieron en incubación por el tiempo necesario hasta la obtención de un desarrollo adecuado de las colonias. La identificación se realizó siguiendo las técnicas rutinarias en micología, tomando en cuenta las estructuras morfológicas, su crecimiento en medio de cultivo y la literatura especializada correspondiente (Gilbertson, 1974; Gilbertson y Ryvarden 1986, 1987; Marmolejo et al. , 1981). Dicha bibliografía se usó, igualmente, para determinar a los hongos recolectados en cambo, después de haberlos deshidratado y herborizado. Se hicieron cultivos de referencia en extracto de malta agar al 2 % para compararlos con los cultivos obtenidos.
La información conjunta se empleó para cuantificar la enfermedad mediante la incidencia y severidad. La incidencia se define como el número de unidades de la planta que están visiblemente enfermas, por lo general en relación al número total evaluado (Madden y Hughes, 1995); se determina mediante porcentajes o proporción de plantas enfermas en una muestra o población, partiendo de la evaluación de parámetros sintomatológicos a nivel individual, tales como: hojas cloróticas, desecación apical, presencia de basidiocarpos y la resinación producida, aplicando la siguiente fórmula:

La severidad determina el nivel de intensidad de una crisis fitopatológica con el fin de estimar la efectividad de las prácticas de manejo del problema para toma de decisiones (Nutter et al ., 1993). Esta severidad expresa la proporción del tejido afectado por la enfermedad y se obtiene mediante la observación de las muestras sacadas a 1.3 m. Con este dato y mediante una escala de severidad propuesta por García y Montero (1998), se conoció el grado en que estaban dañados esos árboles (Cuadro 1). Con estos datos, se aplicó la siguiente fórmula para calcular esa proporción:

Para determinar las variables independientes asociadas a la pudrición blanca, se tomaron en cuenta el diámetro, la altura, la edad y la calidad de sitio de las parcelas. La edad se estimó mediante el conteo de anillos de las muestras tomadas a 1.3 m. Los datos obtenidos fueron procesados y analizados estadísticamente por medio de análisis de regresión utilizando el programa Statistica (7.1) (StatSoft, 2005).
Las variables altura, diámetro y edad se analizaron mediante el modelo de regresión lineal simple, mediante el método de mínimos cuadrados para determinar el coeficiente de correlación, de determinación, error estándar absoluto y el porcentual. El modelo lineal simple utilizado fue el siguiente:
Y = α + β X
Donde:
Y = incidencia de la enfermedad
X = Variable de interés
α = Intercepción al origen
β = Pendiente de la recta
Se construyeron curvas anamórficas de índice de sitio con el objetivo de establecer la capacidad productiva de estos bosques y la relación que tiene con la pudrición blanca. Esta capacidad está en función de la edad y altura de los árboles y para ello se utilizaron cuatro modelos matemáticos no lineales descritos por Kiviste et al. (2002), y se eligió aquel que tuviera mayor ajuste con base al cuadrado medio residual, coeficiente de determinación, desviación estándar residual y coeficiente de variación. Los modelos fueron los siguientes:

Donde:
h = altura
t = años
exp = exponente
α + β = parámetros del modelo
El turno patológico se define como la edad a la cual el árbol pierde lo mismo por pudrición de la madera que lo que gana por incremento anual. Para calcularlo fue necesario tomar en cuenta la severidad presentada a nivel individual, con la finalidad de conocer la proporción de árboles con daño en las diferentes categorías de edad.
RESULTADOS Y DISCUSIÓN
Al hacer las comparaciones entre las colonias de las submuestras y aquellas que provenían de los basidiocarpos, se observó que Phellinus pini es el agente causal de la pudrición y mortandad de los árboles de Pinus greggii , y se determinó que las colonias fueron iguales morfológicamente. Se identificó un caso en el cual Phellinus chrysoloma (Fr.) Donk estaba generando una pudrición central, pero no representó un problema económico para esos bosques.
La incidencia fue diferente entre parcelas, de 11.66 %, en promedio. Los árboles menores a 30 años no presentaron ninguna manifestación de daños, sin embargo, conforme aumentó la edad, la incidencia fue mayor, pues a partir de 50 a 60 años o más, llegó a 25 % (Figura 1).
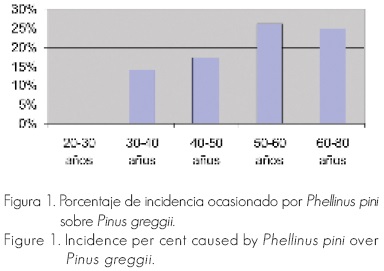
La severidad del ataque por Phellinus pini en árboles de Pinus greggii fue similar a la incidencia; esta severidad causó una pérdida de madera en pie de 12.38 % por el daño que infligió en el duramen de la madera de los anillos más internos. De igual manera, árboles menores a 30 años, no presentaron afectaciones, pero la destrucción de material estructural se incrementó con la edad, alcanzando valores de hasta 26 % en árboles de 60 a 80 años de edad (Figura 2).

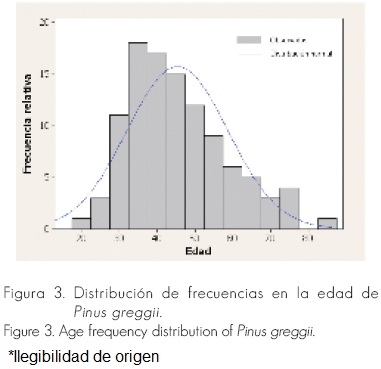
La mayor frecuencia de edades se observó en años menores a la media (Figura 3); si hubiera mayores frecuencias en edades avanzadas, tanto la incidencia como la severidad aumentarían considerablemente.
Es probable que uno o más factores estén interactuando con la pudrición blanca; sin embargo, en esta investigación no se consideran aquellos de tipo abiótico y se enfocó el interés en las variables dasométricas a nivel de rodal como altura, diámetro, edad y calidad de sitio, con una correlación positiva en todos los casos, siendo diferente en cada una de ellas.
La altura presentó una media de 13.43 m, con valores máximos y mínimos de 29.2 y 6.2 respectivamente, y una desviación estándar de 2.9; sin embargo, el valor de correlación fue de 63.77 % y el coeficiente de determinación de 40.67 % (Cuadro 2), lo que indica que la incidencia puede estar influida por otros factores.
El diámetro de los árboles presentó una media de 28 cm, 15 como valor mínimo y 71 cm como máximo y una desviación estándar de 8.28 %. No se tomaron en cuenta árboles menores a 15 cm de diámetro para efecto de este estudio, ya que están poco desarrollados o son muy jóvenes, y, por lo general, no portan manifestaciones de pudrición central. Esta variable presentó una mayor correlación que la altura y, por ende, un mayor coeficiente de determinación, con valores de 78.64 y 61.84 %, respectivamente (Cuadro 2).
La edad tuvo una influencia mucho mayor que las dos variables anteriores, ya que su correlación fue de 92.85 %, un coeficiente de determinación del 86.21 y un bajo error; por ello, se considera que la incidencia de la enfermedad está estrechamente relacionada con la edad, tal y como lo mencionaron García y Montero (1998) (Cuadro 2). El valor medio de la edad fue de 45 años, con intervalos de 20 y 26 años y una desviación estándar del 13 %.
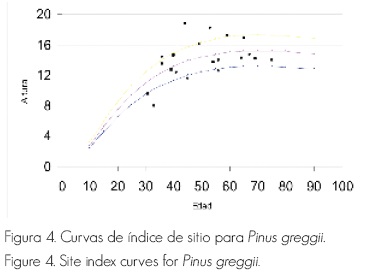
La relación existente entre incidencia e índice de sitio fue baja. El modelo utilizado fue el de Korsun, debido a que es el que mejor se ajustó a los datos y, con base en ello, se construyeron las líneas guías, a partir de tres calidades de sitio. Al analizar los datos, se observa que el índice de sitio no es un indicador de la presencia de Phellinus pini sobre Pinus greggii , debido a que la enfermedad se pudo constatar tanto en sitios con índices buenos, como en aquellos con un índice de sitio muy bajo (Figura 4).
Para determinar el turno patológico, se tomó en cuenta la severidad del daño que este hongo causa en madera en pie. En la Figura 5 se puede advertir que los porcentajes en pérdida de este material cambian conforme se surgen diferentes categorías de edad, siendo el punto más crítico los árboles mayores a 50 años, ya que a partir de entonces el valor es de 45.7 %, casi la mitad de lo que se gana en incremento. Es por esto que estos individuos son considerados los más susceptibles a ser atacados por P. pini , lo que asemeja a este turno al propuesto por Baudin (1963) sobre Pinus pinea L , en el que la pérdida de madera se incrementaba casi en 50%.

CONCLUSIONES
Phellinus pini es el causante de la mortalidad del arbolado de Pinus greggii en los bosques de pino estudiados del estado de Nuevo León, lo que se manifiesta en resinaciones, pudrición central, presencia de basidiocarpos, puntas muertas y clorosis.
La incidencia y la severidad estuvieron fuertemente influidas por las diferentes categorías de edad, la más intensa se registró en árboles de edad avanzada y los jóvenes menores a 30 años no mostraron problemas. Aunque la relación existente se dio en las tres variables (altura, diámetro y edad), la edad fue la que evidenció una mayor correlación con la incidencia de la enfermedad, lo que indica que el problema aumenta mientras la edad de los árboles avanza. La calidad de sitio no tuvo ningún efecto en la presencia de P. pini .
Se estimó un turno fitopatológico de 50 años.
AGRADECIMIENTOS
Se agradece a la CONAFOR (Proyecto: CONAFOR-2003-C03-10178) por el apoyo otorgado para la realización de este estudio.
REFERENCIAS
Bandala, V. M., G. Guzmán y L. Montoya 1993. Los hongos del grupo de los poliporáceos conocidos en México. Facultad de Ciencias Forestales de la UANL. Linares, N.L. Reporte Científico No. Esp. 13: 1-55; [ Links ]
Basham, J. T. and Z. J. R Morawski. 1964. Cull studies. The defects and associated basidiomycete fungi in the heartwood of living trees in the forests of Ontario. Canadian Department of Forestry, Publication 1072, Contribution 1043. Ottawa, Ontario, Canada. 67 p. [ Links ]
Basham, J. T. 1975. Heart rot of jack pine in Ontario. IV. Heartwood-inhabiting fungi, their entry and interactions within living trees. Can. J. For. Res. 5: 706-721. [ Links ]
Baudin, F. 1963. Transformación de las masas de Pinus pinea atacadas por el Trametes pini. Actas de la II Asamblea Técnica forestal. MAPA, Madrid 112-114. [ Links ]
Blanchette, R. A. 1980. Wood decomposition by Phellinus (Fomes) pini: a scanning electron microscopy study. Can. J. Bot. 58: 1496-1503. [ Links ]
Blanchette, R. A. 1982. Phellinus (Fomes) pini decay associated with sweetfern rust in sapwood of jack pine. Can. J. For. Res 12: 304-310. [ Links ]
Blanchette, R. A., L. Otjen and M.C. Carlson. 1987. Lignin distribution in cell wall of birch wood decayed by white rot Basidiomycetes. Phytopathology, 77: 684 - 690. [ Links ]
Cartwright, K. S. G. and Findlay W.P.K. 1950. Decay of Timber and its prevention. Chemical Publ. Co., Brooklyn, NY, USA. 294 pp. [ Links ]
Díaz-Vaz, J. E. 2003. Anatomía de maderas. Facultad de Ciencias Forestales, Universidad Austral de Chile, Valdivia, Chile. 150 p. [ Links ]
Fischer, M. 1994. Pairing tests in the Phellinus pini group. Mycologia 86(4): 524-539. [ Links ]
García, G., C. y G. Montero 1998. Influencia de ciertas variables selvícolas en la pudrición provocada por Phellinus pini sobre Pinus pinea . Invest. Agr.: Sist. Recurs. For. 7(1, 2): 204-218. [ Links ]
Gilbertson, R. L. 1974. Fungi that decay Ponderosa Pine. The Universityof Arizona Press, Tucson, AZ USA.197p. [ Links ]
Gilbertson, R. L. and I. Ryvarden. 1986. North American Polypores. I. Abortiporus-Lindtneria. Fungiflora. Oslo, Norway. pp. 1-433 [ Links ]
Gilbertson R. L. and I. Ryvarden 1987. North American Polypores. II. Megasporoporia-Wrightoporia. Fungiflora. Oslo, Norway. pp. 434-835 [ Links ]
Haddow, R. 1938. On the classification, nomenclature, hosts and geographic range of Trametes pini (Thore) Fries. Transactions of the British Mycological Society 22: 182-93. [ Links ]
Kiviste, A., J. G. Álvarez G., A. Rojo y A. D. Ruiz G. 2002. Funciones de crecimiento de aplicación en el ámbito forestal. Monografías INIA: Forestal No 4. Madrid, España. 190 p. [ Links ]
Larsen, M. J. and I. Melo. 1996. Neotypification of Phellinus pini . Mycologia 88(5): 839 - 843. [ Links ]
Leandro P., V., L. Diorio y F. Forchiassin. 2003. Degradación de madera de álamo por Fomes sclerodermus : producción de enzimas ligninolíticas en aserrín de álamo y cedro. Revista Iberoamericana de Micología 20: 16-20. [ Links ]
Madden, L. V. and G. Hughes 1995. Plant disease incidence: distributions, heterogeneity and temporal analysis. Annu. Rev. Phytopathol. 33: 529-564. [ Links ]
Marmolejo J., G., J. Castillo y G. Guzmán. 1981. Descripción de especies de Teleforáceos poco conocidas en México. Boletín de Sociedad Mexicana de Micología 15: 9-66. [ Links ]
Melo, R. y J. Paz. 1978. Procesos de obtención de celulosa a partir de madera de Eucalyptus globulus Labill. Parte V. Proceso al Sulfito Neutro. Universidad de Concepción. Concepción, Chile. 46 p. [ Links ]
Nutter, F. W., M. L. Gleason, J. H. Jenco and N. C. Christians 1993. Assessing the accuracy, intra-rater repeatability, and inter-rater reliability of disease assessment systems. Phytopathology 83(8): 806-812. [ Links ]
Otjen, L. and Blanchette R. A. 1986. A discussion of microstructural changes in wood during decomposition by white rot basiodiomycetes. Can. J. Bot. 64: 905-911. [ Links ]
Rodríguez B., J. 1998. Patología de la madera. Fundación Conde del Valle de Salazar. Ediciones Mundi-Prensa. Madrid, España. 349 p. [ Links ]
StatSoft. 2005. Statistica (7.1). StatSoft, Inc. Tulsa, Oklahoma, USA. s/p. [ Links ]

