










Services on Demand
Journal
Article
 text in
text in  English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista mexicana de ciencias pecuarias
On-line version ISSN 2448-6698Print version ISSN 2007-1124
Rev. mex. de cienc. pecuarias vol.11 n.1 Mérida Jan./Mar. 2020 Epub June 11, 2020
https://doi.org/10.22319/rmcp.v11i1.4998
Articles
Genetic relationships, biofilm formation, motility and virulence of Escherichia coli isolated from bovine mastitis
a Universidad de La Ciénega del Estado de Michoacán de Ocampo. Genómica Alimentaria. Avenida Universidad No. 3000. Col. Lomas de la Universidad, 59103, Sahuayo, Michoacán, México.
Escherichia coli is an environmental bacterium frequently implicated in bovine mammary infections. Although specific virulence factors are unknown, biofilm-forming bacteria are associated with persistent infections, and motile bacteria exhibit increased virulence. An analysis was done of the genetic relationship, biofilm formation capacity and motility of bovine mastitis-associated E. coli, as well as the in vivo virulence of representative bacterial isolates. Of the 34 isolates, 67.7 % belonged to phylogenetic group A, 17.6 % to group B1 and 14.7 % to group D. Genetic relationship analysis done with (GTG)5-PCR indicated that the analyzed bacteria are diverse, with only two isolates exhibiting 100 % similarity, and the remaining 32 being grouped into seven main clusters with more than 70 % similarity. Biofilm formation capacity ranged from strong to moderate and weak in 76.5 % of the isolates. The csgA and fimA genes were detected in 52.9 % of the biofilm-forming isolates. Most (70.6 %) of the isolates were motile. In vivo infection assays using Galleria mellonella showed the biofilm-forming bacteria to be more pathogenic than the non-biofilm-forming ones. The studied bovine mastitis-associated E. coli were genetically diverse. Biofilm formation capacity and motility were variable among the isolates, but the biofilm-forming bacteria were more pathogenic than the non-biofilm-forming ones.
Key words E. coli; Bovine mastitis; Diversity; Biofilm; Virulence
Escherichia coli es una de las bacterias ambientales que frecuentemente se ha visto implicada en infecciones de la glándula mamaria bovina. Aunque se desconocen los factores de virulencia específicos involucrados en dicho proceso, se ha observado que las bacterias formadoras de biopelículas se asocian con infecciones persistentes. De igual forma se ha planteado que las bacterias móviles pueden ser más patógenas. El propósito del trabajo fue analizar la relación genética, capacidad de formación de biopelículas y movilidad de E. coli aislada de casos de mastitis bovina, así como la virulencia in vivo de aislados representativos. El 67.7 % de las bacterias pertenece al grupo filogenético A, el 17.6 % al grupo B1 y el 14.7 % al D. El estudio de la relación genética mediante (GTG)5 reveló que solo dos bacterias son idénticas genéticamente, el resto se organizaron en siete grupos distintos con 70 % de similitud. El 76.5 % de las bacterias fue capaz de formar biopelículas de forma fuerte, moderada o débil. Además, los genes csgA y fimA se detectaron en el 52.9 % de las bacterias formadoras de biopelículas. El análisis de movilidad mostró que el 70.6 % fue móvil. Mientras que, el análisis de patogenicidad en larvas de Galleria mellonella reveló que bacterias formadoras de películas y móviles fueron capaces de matar un mayor número de larvas a las 24 h que las no formadoras. Los resultados indican que las E. coli causantes de mastitis bovina son muy diversas y pertenecen principalmente al grupo filogenético A y que las formadoras de biopelículas y móviles son más patógenas.
Palabras clave E. coli; Mastitis bovina; Diversidad; Biopelículas; Virulencia
Introduction
Bovine mastitis is the principal cause worldwide of economic losses in the dairy industry due to decreased milk production, treatment costs, and early disposal of cattle, among other factors1. Escherichia coli is one of the main environmental pathogens associated with bovine mastitis, and its infections can range from moderate to severe2,3. The specific virulence factors of bovine mastitis-associated E. coli have not been determined to date, but it may form part of a new pathotype known as Mammary Pathogenic E. coli (MPEC)4. More recent studies at the genomic level have revealed the presence of groups of genes that code for the type VI secretion systems, lipopolysaccharide biosynthesis, biofilm formation and iron uptake systems characteristic of bovine mastitis-associated E. coli (BMAEC)5-8. However, other studies suggest that evidence is still insufficient to assign BMAEC to a particular pathotype9.
Several phylogenetic analyses of E. coli have classified this bacterium into four main groups: A, B1, B2 and D. Non-pathogenic commensal bacteria belong mainly to groups A and B1, although these groups also include important pathogens. Group B2 and a smaller proportion of D contain strains associated with extra-intestinal infections9,10. Bovine mastitis-associated E. coli (BMAEC) have been assigned mainly to groups A and B111,12,13. In addition, genetic and genomic analyses have revealed that BMAEC are quite diverse; indeed, the E. coli associated with both persistent and transient bovine mastitis exhibit both phenotypic and genotypic diversity14. Genomic analysis of various E. coli isolates which cause bovine mastitis and commensals has shown that none can be classified into a particular phylogenetic group and that in both cases they apparently originated from different lineages9.
The ability to colonize and invade the bovine mammary epithelium helps bacteria evade the immune response and infect persistently. The E. coli behind persistent mastitis efficiently invades mammary epithelial cells, and, although the mechanisms it uses are not well known, the bacteria exhibiting greater motility also have greater virulence15. The extracellular polymer matrix of biofilm-forming bacteria makes them more resistant to various chemical substances with antimicrobial activity produced by cells, as well as protecting them from the innate immune system16,17.
Bacterial virulence is generally studied using various animal models, although many of these can be expensive and present some ethical problems. A viable and increasingly popular model uses larvae of the greater wax moth Galleria mellonella to measure the virulence of Gram positive and negative pathogenic bacteria, as well as fungi18,19. For example, G. mellonella is a suitable model for study of the pathogenic variants of both intestinal and extraintestinal E. coli20,21,22.
The present study objective was to analyze the genetic diversity, identify phylogenetic groups, and quantify biofilm formation capacity and motility of E. coli isolated from cases of subclinical bovine mastitis, and evaluate the in vivo virulence of representative isolates using a G. mellonella larvae model.
Material and methods
DNA extract preparation and phylogenetic group identification
The studied E. coli were 34 antibiotic-resistant strains isolated from cases of subclinical bovine mastitis in western Michoacán state, Mexico23. Total DNA extracts were prepared from these isolates24. An established protocol was used to identify the phylogenetic groups of each strain10. The primers used for fragment amplification were:
ChuA.1 (5'-GACGAACCAACGGTCAGGAT-3');
ChuA.2 (5'-TGCCGCCAGTACCAAAGACA-3');
YjaA.1 (5'-TGAAGTGTCAGGAGACGCTG-3');
YjaA.2 (5'-ATGGAGAATGCGTTCCTCAAC-3');
TspE4C2.1 (5'-GAGTAATGTCGGGGCATTCA-3');
TspE4C2.2 (5'-CGCGCCAACAAAGTATTACG-3').
The PCR reactions were run using a 25 µl volume containing: 1X PCR Master mix (Promega, Madison, WI, USA), 1 µM of each primer and 2 µl bacterial DNA extract (50 ng). Fragment amplification was done in a C1000 thermocycler (Bio-Rad, Mexico City, Mexico) under the following conditions: initial denaturation at 94 °C for 5 min; 30 cycles as follows, denaturation at 94 °C for 30 s, alignment at 55 °C for 30 s and extension at 72 °C for 30 s; and final extension at 72 °C for 5 min. The amplified products were separated on 1.5% agarose gels and viewed with a Universal Hood II Gel Doc (Bio-Rad).
Biofilm production
Biofilm production was grown according to an established protocol25 with some modifications. Bacteria were seeded in Petri dishes with trypticase soy agar (TSA, BD, Mexico City, Mexico). These cultures were used to inoculate tubes with 2 ml trypticase soy broth (TSB) containing 0.25% glucose, which were incubated overnight at 37 °C under constant agitation. In each tube the cultures were diluted to the 0.5 MacFarland standard with TSB containing 0.25% glucose. From these bacterial dilutions 200 μl were deposited in triplicate in 96-well ELISA plates and incubated for 24 h at 37 °C without stirring. Non-inoculated culture medium was used as a negative control, and the biofilm-forming bacterium Pseudomonas aeruginosa ATCC 27853 was used as a positive control. After incubation the culture medium was discarded, the wells washed three times with sterile saline, and the plate allowed to dry at 60 °C for 1 h to fix the cells. One percent (1%) violet crystal (200 µl) was added to the wells and the plate left at room temperature for 20 min. It was washed with running water until no color evolution was observed and allowed to dry at room temperature. The violet crystal was solubilized by adding 200 µl 96% ethanol to each well and stirring, and transferring the supernatant to a microcentrifuge tube. This process was repeated, tube content completed to 1 ml and optical density (OD) of the suspension read at 570 nm with a SmartSpec Plus spectrophotometer (Bio-Rad). This test was repeated four times in triplicate and average OD calculated from the results. The optical density cutoff (ODc) was calculated using the average OD of the negative control and increasing the value by three times its standard deviation, producing a value of 0.22. The biofilm-forming bacteria were classified into four groups: strong= OD> 4xODc; moderate= 2xODc<OD≤4xODc; weak= ODc<OD≤2xODc; and negative= OD≤ODc25.
Detection of genes associated to biofilm formation
The presence of two genes associated with biofilm formation, fimA (main subunit of type I fimbria) and csgA (main subunit of curli fimbria), was analyzed by PCR. A 119 bp fimA fragment was amplified with fimA-F (5'-CTCTGGCAATCGTTGTTCTGTCG-3') and fimA-R (5'-GCAAGCGGCGTTAACAACTTCC-3'), and a 178 bp csgA fragment was amplified with csgA-F (5'-GATCTGACCCAACGTGGCTTCG-3') and csgA-R (5'-GATGAGCGGTCGCGTTGTTACC-3')26. The reactions were run in a 25 µl volume containing 1X PCR Master mix (Promega), 1 µM of each primer, and 2 µl total DNA extract. Amplification conditions were: initial denaturation at 95 °C for 5 min; 30 cycles of 94 °C for 30 s, 55 °C for 30 s and 72 °C for 30 s; and a final extension cycle at 72 °C for 10 min. The amplified products were separated on 2% agarose gels and viewed as described above.
Determination of swarming type motility
Bacterial motility was measured following an established protocol15. Briefly, swarming motility was determined by allowing the cultures to grow overnight in Lysogeny Broth (LB), and adding 5 µl swarming agar (LB + 0.5% agar) containing 0.5% glucose. After incubating the cultures for 12 h at 37 °C, bacteria displacement diameter was measured. Three measurements were taken and averaged.
Rep-PCR of bovine mastitis-causing E. coli
Genetic diversity was analyzed by purifying genomic DNA according to standard protocols27. The purified DNA was amplified in 25 µl reaction mixture containing 12.5 µl 2X Master mix (Promega), 3 mM MgCl2, 5% DMSO, 0.16 µg/µl bovine serum albumin, 100 ng DNA and 2 µM of the primer (GTG)5 (5'-GTGGTGGTGGTGGTG-3'). Amplification was done in a C-1000 thermocycler (Bio-Rad) under the following sequence: initial denaturation 95 °C for 2 min; 30 cycles as follows, denaturation (94 °C/3 s and 92 °C/30 s), alignment (40 °C/1 min) and extension (65 °C/8 min); and final extension at 65 °C for 8 min28. The amplified products were separated on 1.5% agarose gels and viewed with a Universal Hood II Gel Doc (Bio-Rad).
Band patterns were analyzed with the GelJ ver. 2 software29. Band size was standardized using a 1 kb DNA ladder (Promega) with fragments between 250 and 10,000 bp as a reference standard. The similarity coefficients were generated by the Dice method with a 2 % tolerance between rows, and the dendrogram built using the UPGMA method.
Virulence in Galleria mellonella
In vivo virulence was measured for representative bacteria exhibiting the highest biofilm formation rates, as well as non-biofilm-forming isolates. The G. mellonella larvae weighed 150 to 200 mg, were free of apparent damage, exhibited no melaninization (necrotic spots) (Petmmal, Cuautitlán Izcalli, Mexico), and were stored at 30 °C in darkness until use. The infection assays were done using bacterial cultures grown in LB broth until the mean exponential growth phase (OD 600 nm= 0.5). These were centrifuged at 12,000 rpm/min, washed twice with 1 ml 10 mM MgSO4, and resuspended in 1 ml 10 mM MgSO4. Serial dilutions were made of this cell suspension until attaining a bacterial concentration of approximately 1x105 CFU/ml, which was confirmed by CFU count on LB agar.
Ten G. mellonella larvae were used for each bacterium to be studied. Using an insulin syringe with a 29G gauge needle, 10 μl bacterial suspension (1 x 105 CFU/ml) were injected into the final pro-leg of each larva. The negative control was ten larvae injected with 10 mM of bacteria-free MgSO4, and an additional ten without inoculation. The inoculated and non-inoculated larvae were placed in sterile Petri dishes and incubated at 30 °C in darkness for 96 h. Survival percentages were measured at 24, 48, 72 and 96 h; larvae unresponsive to touch were recorded as dead21. The virulence assay for each bacterium was run at least twice independently.
Results
Phylogenetic groups and genetic relationship of bovine mastitis- associated E. coli
Amplification of the chuA (279 bp), yjaA (211 bp) and TspE4.C2 (152 bp) fragments (Figure 1) showed the 34 isolated BMAEC strains to belong to three phylogenetic groups. Most (67.7 %; 23 isolates) belonged to group A, followed by 17.6 % (6 isolates) in group B1 and the remaining 14.7 % (5 isolates) in group D (Figure 1B).
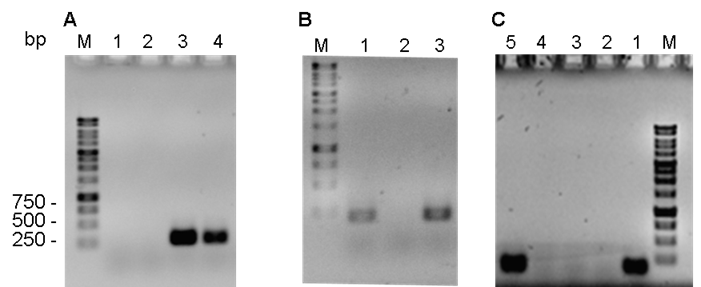
Figure 1 Agarose gels showing PCR amplification results for A) chuA (279 bp), B) yjaA (211 bp) and C) TspE.C2 (152 bp)
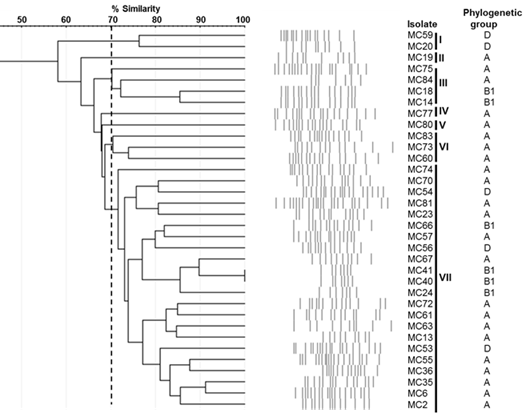
The genetic relationship analysis by rep-PCR using the (GTG)5 oligonucleotide yielded 32 different patterns between 7 and 21 bands, with sizes ranging from 250 to 5,000 bp (Figure 2). The dendrogram based on the Dice similarity coefficient and generated by the UPGMA method indicated that the lowest similarity among the isolates was 58 % while the highest was 100 % (in 2 isolates) (Figure 3). Considering a 70 % genetic similarity level (dotted line), seven groups of bacteria were identified; the largest was group VII with 22 isolates (64.7 %), followed by group III with 4 isolates (11.8 %), group VI with 3 isolates (8.8 %), group I with 2 isolates (5.8 %), and groups II, IV and V with a single isolate each (2.9 %).
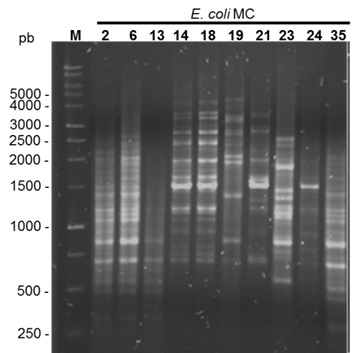
Figure 2 Agarose gel of products amplified by (GTG)5-PCR. The base pair size of some bands is indicated by the 1 kb marker (M)
Motility, biofilm formation and associated genes
Ten of the 34 E. coli isolates were not mobile (-) and the remaining 24 were mobile. Based on their displacement diameters, twelve of these were minimally mobile (+), six were moderately mobile (++), and six were highly mobile (+++) (Figure 4, Table 1).
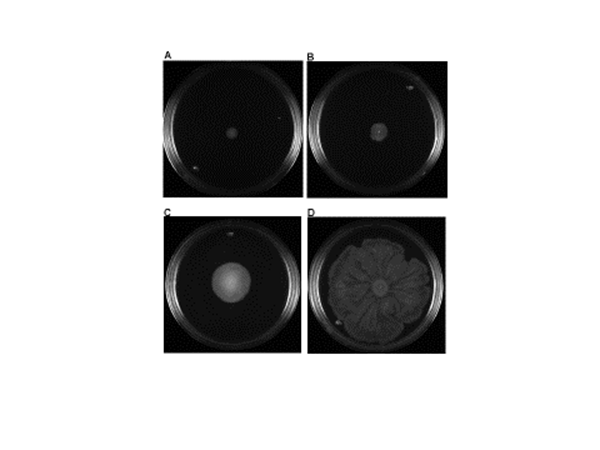
Figure 4 Representative examples of “swarming” type motility. A) Not mobile, B) Minimally mobile, C) Moderately mobile, D) Highly mobile
Table 1 Motility, biofilm formation and frequency of biofilm formation-associated genes of the 34 E. coli isolates from subclinical bovine mastitis cases
| E. coli | Motilitya | Biofilmb | Genotype csgA/fimA |
|---|---|---|---|
| MC75 | +++ | W | +/+ |
| MC80 | + | N | +/+ |
| MC81 | + | W | +/+ |
| MC83 | + | N | +/+ |
| MC13 | - | S | -/- |
| MC14 | - | W | +/+ |
| MC72 | +++ | M | +/+ |
| MC19 | ++ | S | -/+ |
| MC40 | +++ | W | +/+ |
| MC41 | + | N | +/+ |
| MC59 | - | W | -/- |
| MC73 | + | M | -/- |
| MC66 | - | W | +/+ |
| MC2 | - | S | -/- |
| MC6 | + | S | -/- |
| MC35 | ++ | S | -/+ |
| MC36 | + | S | -/+ |
| MC84 | - | W | -/- |
| MC24 | + | W | +/+ |
| MC56 | ++ | W | +/+ |
| MC67 | - | N | +/+ |
| MC74 | ++ | M | -/- |
| MC77 | ++ | N | -/- |
| MC55 | + | S | -/- |
| MC60 | + | W | -/- |
| MC54 | +++ | M | +/+ |
| MC18 | + | W | +/+ |
| MC23 | - | M | -/+ |
| MC57 | - | M | -/- |
| MC53 | +++ | N | +/+ |
| MC20 | + | N | +/+ |
| MC70 | +++ | W | +/+ |
| MC61 | ++ | N | +/+ |
| MC63 | - | M | -/- |
a(-) not mobile, (+) minimally mobile, (++) moderately mobile, (+++) highly mobile.
bN, negative; W, weak; M, moderate; S, strong.
Twenty-six (76.5 %) of the isolates formed biofilm to some degree (OD>0.22) while the remaining eight (23.5 %) did not form biofilm (OD≤0.22). Of the biofilm-forming isolates seven were classified as having a strong (OD>0.88) and moderate formation capacity (OD 0.45-0.88), and twelve as having a weak one (OD 0.23-0.44) (Table 1). Twenty-two (64.7 %) of the 34 isolates had either csgA or fimA, whereas of the 26 biofilm-forming isolates 15 had one or both of these genes. Both csgA and fimA were present in the seven non-biofilm-forming isolates.
E. coli Virulence in G. mellonella
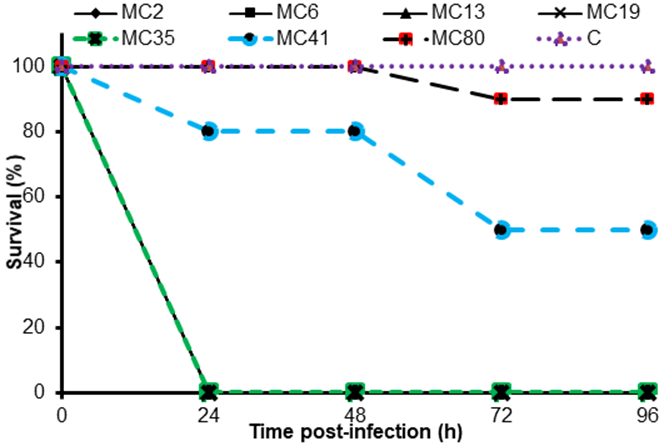
Differences in the pathogenesis of the BMAEC isolates were analyzed by infection trials using G. mellonella larvae. Seven representative isolates were tested: five strong biofilm-forming isolates and two non-forming isolates. The biofilm-forming isolates killed 100 % of the larvae 24 h after infection (Figure 5). The two non-biofilm-forming isolates caused from 0 to 20 % mortality at 24 h, and from 10 to 50 % at 96 h. The control resulted in no mortality during the 96-h experiment.
Discussion
The severity of E. coli-caused bovine mastitis can range from mild to severe. No specific virulence factors have yet been associated with these disease-causing bacteria11,30, and it has even been suggested that infection severity is determined by cattle characteristics rather than by the microorganisms31. One line of evidence suggests that mammary gland infections can be caused by any E. coli strain11, while another has shown that not all E. coli isolates are capable of producing mastitis in animal models5.
The E. coli strains known to cause bovine mastitis belong mainly to the A and B1 phylogenetic groups, and to a lesser extent to the D group7. This coincides with the present results in which 67.7 % of the analyzed isolates belong to phylogenetic group A, 17.6 % to B1 and 14.7 % to D. This overall pattern has been reported previously, with 50 % of the E. coli strains associated with both persistent and transient mastitis belonging to the A phylogenetic group, 28.6 % to B1 and 7.1 % to groups B2 and D, respectively14. In another study 44.88 % of mastitis-associated E. coli belonged to phylogenetic group A, 38.58 % to B1 and 16.53 % to D32. Similar results have also been reported elsewhere13,33. In addition, the E. coli isolated from bovine mastitis exhibit great diversity in terms of serotype, disease type (transient or persistent) and genotype, even when they belong to the same phylogenetic group3,34. The 34 E. coli isolates analyzed here also exhibited broad genetic diversity, since only two had identical band patterns and the remaining 32 had similarity ranging from 92 to 58 %. The present results also indicated no clear separation between the different phylogenetic groups, which shows great heterogeneity among the bovine mastitis-causing E. coli strains in the study area; this also coincides with previous reports3,30,34,35.
The ability to form biofilms in pathogenic bacteria provides them protection against the host immune system and antibiotic resistance, as well as affecting virulence26. The bacteria associated with bovine mastitis manifest a variable pattern of biofilm formation36, although recurrent infections have been associated with biofilm-forming strains16. In the present results 20.6 % of the isolates formed strong to moderate biofilms, while 35.3 % formed weak ones. Other studies have reported similar results with different BMAEC strains having variable biofilm forming capabilities. In one, 18.5 % of the strains had a strong capacity, while 40.7 % had a moderate to weak capacity36, and in another study 40 % had strong capacity, 12 % had a moderate capacity and 4 % had a weak one33.
Among the 34 analyzed E. coli, the csgA and fimA genes were detected in 52.9 %, only fimA in 11.8 %, and neither gene in the remaining 35.3 %. At least one of the genes was detected in 15 of the 26 biofilm-forming isolates and the seven non-biofilm forming isolates. Both genes have been associated with biofilm formation, as have a variety of other genes and environmental conditions37. Further research is clearly needed to generate a more detailed characterization of the factors affecting or promoting biofilm formation in the studied E. coli isolates. Biofilm formation can also vary in response to strain, culture medium, methodology and quantification method. Only one culture medium and one temperature were tested in the present study, raising the possibility that under different conditions the non-biofilm-forming bacteria that were csgA- and fimA-positive may be capable of forming biofilms38.
Bacterial motility is another important factor in the spread and production of persistent infections in cattle15. In the present results 70.6 % of the studied E. coli were mobile, suggesting that there were both persistent and transient mastitis bacteria among the studied strains.
The in vivo results indicated that the biofilm-forming isolates exhibited greater virulence against G. mellonella larvae than the non-biofilm-forming isolates. More study is required to establish the correlation between pathogenesis in vivo in G. mellonella with the severity of E. coli-caused mastitis, but the present results do suggest that this organism is an adequate model for analyzing the pathogenesis of mastitis-causing E. coli.
Conclusions and implications
The bovine mastitis-associated E. coli analyzed here were genetically and physiologically diverse. The isolates strongly capable of forming biofilms were more pathogenic than the non-forming isolates. The observed genetic and phenotypic diversity indicates that there is no strain, genotype or specific virulence factor associated with mastitis. However, because biofilm-forming bacteria have been associated with recurrent and persistent mammary gland infections, better strategies for controlling infections need to be developed with the aim of reducing the economic losses associated with the consequent reductions in milk production and quality.
Acknowledgements
The research reported here was financed by the PRODEP (IDCA-11106) and UCEMICH (UCEMICH-2016-006 and UCEMICH-2017-004). The authors thank Dr. Iván Medina and Dr. José Antonio Aguilar for their assistance with manuscript review.
REFERENCES
1. Halasa T, Huijps K, Østerås O, Hogeveen H. Economic effects of bovine mastitis and mastitis management: A review. Vet Q 2007;29(1):18-31. [ Links ]
2. Wenz JR, Barrington GM, Garry FB, Ellis RP, Magnuson RJ. Escherichia coli isolates’ serotypes, genotypes, and virulence genes and clinical coliform mastitis severity. J Dairy Sci 2006;89(9):3408-3412. [ Links ]
3. Hagiwara S, Mori K, Okada H, Oikawa S, Nagahata H. Acute Escherichia coli mastitis in dairy cattle: diagnostic parameters associated with poor prognosis. J Vet Med Sci 2014;76(11):1431-1436. [ Links ]
4. Shpigel NY, Elazar S, Rosenshine I. Mammary pathogenic Escherichia coli. Curr Opin Microbiol 2008;11(1):60-65. [ Links ]
5. Blum SE, Heller ED, Sela S, Elad D, Edery N, Leitner G. Genomic and phenomic study of mammary pathogenic Escherichia coli. PLoS ONE 2015;10(9):e0136387. [ Links ]
6. Kempf F, Slugocki C, Blum SE, Leitner G, Germon P. Genomic comparative study of bovine mastitis Escherichia coli. PLoS ONE 2016;11(1):e0147954. [ Links ]
7. Goldstone RJ, Harris S, Smith DG. Genomic content typifying a prevalent clade of bovine mastitis-associated Escherichia coli. Sci Rep 2016;(6):30115. [ Links ]
8. Olson MA, Siebach TW, Griffitts JS, Wilson E, Erickson DL. Genome-wide identification of fitness factors in mastitis-associated Escherichia coli. Applied Environ Microbiol 2018;84(2):e02190-17. [ Links ]
9. Leimbach A, Poehlein A, Vollmers J, Görlich D, Daniel R, Dobrindt U. No evidence for a bovine mastitis Escherichia coli pathotype. BMC Genomics 2017:18(1):359. [ Links ]
10. Clermont O, Bonacorsi S, Bingen E. Rapid and simple determination of the Escherichia coli phylogenetic group. Appl Environ Microbiol 2000;66(10):4555-4558. [ Links ]
11. Suojala L, Pohjanvirta T, Simojoki H, Myllyniemi AL, Pitkälä A, Pelkonen S, Pyörälä S. Phylogeny, virulence factors and antimicrobial susceptibility of Escherichia coli isolated in clinical bovine mastitis. Vet Microbiol 2011;147(3-4):383-388. [ Links ]
12. Liu Y, Liu G, Liu W, Liu Y, Ali T, Chen W, Yin J, Han B. Phylogenetic group, virulence factors and antimicrobial resistance of Escherichia coli associated with bovine mastitis. Res Microbiol 2014;165(4):273-277. [ Links ]
13. Keane OM. Genetic diversity, the virulence gene profile and antimicrobial resistance of clinical mastitis-associated Escherichia coli. Res Microbiol 2016;167(8):678-684. [ Links ]
14. Dogan B, Rishniw M, Bruant G, Harel J, Schukken YH, Simpson KW. Phylogroup and lpfA influence epithelial invasion by mastitis associated Escherichia coli. Vet Microbiol 2012;159(1-2):163-170. [ Links ]
15. Lippolis JD, Brunelle BW, Reinhardt TA, Sacco RE, Nonnecke BJ, Dogan B, Simpson K, Schukken YH. Proteomic analysis reveals protein expression differences in Escherichia coli strains associated with persistent versus transient mastitis. J Proteomics 2014;(108):373-381. [ Links ]
16. Melchior MB, Vaarkamp H, Fink-Gremmels J. Biofilms: a role in recurrent mastitis infections?. Vet J 2006;171(3):398-407. [ Links ]
17. Atulya M, Mathew AJ, Rao JV, Rao CM. Influence of milk components in establishing biofilm mediated bacterial mastitis infections in cattle: a fractional factorial approach. Res Vet Sci 2014;96(1):25-27. [ Links ]
18. Ramarao N, Nielsen-Leroux C, Lereclus D. The insect Galleria mellonella as a powerful infection model to investigate bacterial pathogenesis. J Vis Exp 2012;(70):e4392. [ Links ]
19. Nathan S. New to Galleria mellonella: modeling an ExPEC infection. Virulence 2014;5(3):371-374. [ Links ]
20. Alghoribi MF, Gibreel TM, Dodgson AR, Beatson SA Upton M. Galleria mellonella infection model demonstrates high lethality of ST69 and ST127 uropathogenic E. coli. PLoS ONE 2014;9(7):e101547. [ Links ]
21. Morgan JK, Ortiz JA, Riordan JT. The role for TolA in enterohemorrhagic Escherichia coli pathogenesis and virulence gene transcription. Microb Pathog 2014;(77):42-52. [ Links ]
22. Ciesielczuk H, Betts J, Phee L, Doumith M, Hope R, Woodford N, Wareham DW. Comparative virulence of urinary and bloodstream isolates of extra-intestinal pathogenic Escherichia coli in a Galleria mellonella model. Virulence 2015;6(2):145-151. [ Links ]
23. Jiménez-Mejía R, Gudiño-Sosa LF, Aguilar-López JA, Loeza-Lara PD. Caracterización molecular de Escherichia coli resistente a antibióticos aislada de mastitis bovina en Michoacán, México. Rev Mex Cienc Pecu 2017;8(4):387-396. [ Links ]
24. Madico G, Akopyants NS, Berg DE. Arbitrarily primed PCR DNA fingerprinting of Escherichia coli O157: H7 strains by using templates from boiled cultures. J Clin Microbiol 1995;33(6):1534-1536. [ Links ]
25. Stepanović S, Vuković D, Hola V, Bonaventura GD, Djukić S, Ćirković I, Ruzicka F. Quantification of biofilm in microtiter plates: overview of testing conditions and practical recommendations for assessment of biofilm production by staphylococci. APMIS 2007;115(8):891-899. [ Links ]
26. Silva VO, Soares LO, Júnior AS, Mantovani HC, ChangYF, Moreira MAS. Biofilm formation on biotic and abiotic surfaces in the presence of antimicrobials by Escherichia coli isolates from cases of bovine mastitis. Appl Environ Microbiol 2014;80(19):6136-6145. [ Links ]
27. Wilson K. Preparation of genomic DNA from bacteria. Curr Protoc Mol Biol 2001;56(1):241-245. [ Links ]
28. Mohapatra BR, Broersma K, Mazumder A. Comparison of five rep-PCR genomic fingerprinting methods for differentiation of fecal Escherichia coli from humans, poultry and wild birds. FEMS Microbiol Lett 2007;277(1):98-106. [ Links ]
29. Heras J, Domínguez C, Mata E, Pascual V, Lozano C, Torres C, Zarazaga M. GelJ-a tool for analyzing DNA fingerprint gel images. BMC Bioinformatics 2015;(16):270. [ Links ]
30. Blum SE, Leitner G. Genotyping and virulence factors assessment of bovine mastitis Escherichia coli. Vet Microbiol 2013;163(3-4):305-312. [ Links ]
31. Burvenich C, Van Merris V, Mehrzad J, Diez-Fraile A, Duchateau L. Severity of E. coli mastitis is mainly determined by cow factors. Vet Res 2003;34(5):521-564. [ Links ]
32. Ghanbarpour R, Oswald E. Phylogenetic distribution of virulence genes in Escherichia coli isolated from bovine mastitis in Iran. Res Vet Sci 2010;88(1):6-10. [ Links ]
33. Milanov D, Prunić B, Velhner M, Todorović D, Polaček V. Investigation of biofilm formation and phylogenetic typing of Escherichia coli strains isolated from milk of cows with mastitis. Acta Veterinaria 2015;65(2):202-216. [ Links ]
34. Dogan B, Klaessig S, Rishniw M, Almeida RA, Oliver SP, Simpson K, Schukken YH. Adherent and invasive Escherichia coli are associated with persistent bovine mastitis. Vet Microbiol 2006; 116(4):270-282. [ Links ]
35. Moser A, Stephan R, Corti S, Lehner A. Resistance profiles and genetic diversity of Escherichia coli strains isolated from acute bovine mastitis. Schweiz Arch Tierheilkd 2013;155(6):351-357. [ Links ]
36. Fernandes JBC, Zanardo LG, Galvão NN, Carvalho IA, Nero LA, Moreira MAS. Escherichia coli from clinical mastitis: serotypes and virulence factors. J Vet Diagn Invest 2011;23(6):1146-1152. [ Links ]
37. Flemming HC, Wingender J, Szewzyk U, Steinberg P, Rice SA, Kjelleberg S. Biofilms: an emergent form of bacterial life. Nat Rev Microbiol 2016;14(9):563-575. [ Links ]
38. Naves P, Del Prado G, Huelves L, Gracia M, Ruiz V, Blanco J, Rodríguez-Cerrato V, Ponte MC, Soriano F. Measurement of biofilm formation by clinical isolates of Escherichia coli is method‐dependent. J Appl Microbiol 2008;105(2): 585-590. [ Links ]
Received: July 24, 2018; Accepted: January 16, 2019
 Este es un artículo publicado en acceso abierto bajo una licencia
Creative Commons
Este es un artículo publicado en acceso abierto bajo una licencia
Creative Commons
