










Services on Demand
Journal
Article
 text in
text in  English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista mexicana de ciencias pecuarias
On-line version ISSN 2448-6698Print version ISSN 2007-1124
Rev. mex. de cienc. pecuarias vol.10 n.4 Mérida Oct./Dec. 2019 Epub Apr 30, 2020
https://doi.org/10.22319/rmcp.v10i4.4908
Articles
Genetic diversity and structure in Criolla Negra goats in Queretaro, Mexico.
a Universidad Autónoma de Querétaro. Facultad de Ciencias Naturales. Doctorado en Ciencias Biológicas. Avenida de las Ciencias S/N Juriquilla, Delegación Santa Rosa Jáuregui, 76230 Querétaro, México.
b Universidad Autónoma de Querétaro. Facultad de Ciencias Naturales. Licenciatura en Medicina Veterinaria y Zootecnia. México.
c INIFAP, Centro Nacional de Investigación Disciplinaria en Fisiología y Mejoramiento Animal. México.
d Universidad de Córdoba. Campus de Excelencia Internacional Agroalimentario ceiA3. Departamento de Genética. España.
eProyecto de Biodiversidad Caprina Iberoamericana. España.
Since their introduction to Mexico goats have undergone a long process of adaptation and selection, resulting in highly rustic local animals. However, importation of improved breeds has led to the extinction of some regional breeds. For example, the Criolla Negra goat breed is known for its rusticity and high milk quality, but is in decline. A genetic characterization was done of a Criolla Negra population. Hair samples were collected in three goat herds located in different municipalities of the state of Querétaro, Mexico: Cadereyta de Montes (n= 7); El Marqués (n= 11); and San Juan del Río (n= 27). Thirty microsatellites were used to quantify the number of alleles per marker (NA), median number of alleles (MNA), number of effective alleles (NEA), observed heterozygosis (Ho), expected heterozygosis (He), polymorphic data content (PDC), the fixation index (FIS) and Hardy Weinberg equilibrium (HWE). The Criolla Negra population was compared to thirteen breeds forming part of the BioGoat project. Genetic diversity was found to be high in this population. A total of 243 alleles were identified with an MNA of 8.1 alleles per marker. The markers were informative (PDC= 0.06) for polymorphism. The He (0.71) and Ho (0.62) values indicate a slight imbalance in the population. Reynolds genetic distance results showed the Criolla Negra breed to be genetically furthest from the Anglonubia breed and nearest the Murciano-Granadina breed. The studied Criolla Negra goat population exhibits a breed structure well differentiated from the other breeds in the analysis.
Key words Genetic characterization; population genetics; Criolla Negra goats
Desde su llegada a México, las cabras han experimentado un largo proceso de adaptación y selección, resultando animales locales de elevada rusticidad. Sin embargo, la importación de razas mejoradas, ha inducido la extinción de algunas razas. Por ejemplo, la cabra criolla negra (CCN), reconocida por su rusticidad y la calidad de su leche. El objetivo del presente trabajo fue caracterizar genéticamente una población de CCN. Se colectaron muestras de pelo en tres rebaños caprinos ubicados en diferentes municipios del estado de Querétaro; Cadereyta de Montes (n= 7), El Marqués (n= 11) y San Juan del Río (n= 27). Se utilizaron 30 microsatélites, obteniendo; el número de alelos por marcador (NA), el número medio de alelos (NMA), número efectivo de alelos (NEA), la heterocigosis observada (Ho) y esperada (He), el contenido de información polimórfica (CIP), el índice de fijación (FIS) y el equilibrio de Hardy Weinberg (EHW). La población se comparó con 13 razas del proyecto BioGoat. Los resultados mostraron que existe una elevada diversidad genética en este ganado. Se obtuvieron 243 alelos con un NMA de 8.1 alelos por marcador. Los marcadores resultaron informativos (CIP= 0.06) respecto a su polimorfismo. La He (0.71) y Ho (0.62) indican que existe un ligero desequilibrio en la población. La distancia de Reynolds mostró que la CCN se encuentra más distanciada genéticamente de la población Anglonubia y más cercana a la Murciano-Granadina. Los resultados aquí presentados sugieren que la población CCN representa una estructura racial bien diferenciada de las poblaciones incluidas en el estudio.
Palabras clave Caracterización genética; Genética de poblaciones; Cabra criolla negra
Introduction
Goats have closely coexisted with human beings since their domestication began approximately 10,000 years ago1,2. One of the first domesticated livestock species, goats formed part of the Neolithic agricultural revolution, development of trade and human migrations3. All these events involved some basic evolutionary mechanisms, such as animal migration, selection, gene drift and even mutation. This helps in explaining goats’ high capacity for adaptation to different ecosystems and the more than 300 breeds currently in existence1,4.
First brought to the Americas in 1493, goats were initially propagated by the Spanish although native peoples also learned how to manage them and select for certain traits. New breeds, known as Creoles, consequently arose over time as selection aimed to better adapt them to local environmental conditions5.
There are currently over 8.7 million goats in Mexico6. As in other developing countries goats represent a subsistence resource for people living in arid and semi-arid areas with scarce vegetation and poor rangeland7,8. Goats can be found throughout Mexico but are far more common in three main regions: the Mixtec mosaic; central Mexico (El Bajio); and northern Mexico (El Lagunero)9. The Criolla Negra (CN) breed is found primarily in the central region, particularly in the states of Querétaro and Guanajuato. Used mostly for dairy production, milk from this breed contains higher total solids content than other goat breeds in Mexico and provides excellent cheese yield10,11,12. Long considered a Granadina breed based on its morphological characteristics and origin, over 500 yr of independent evolution have genetically differentiated it from this breed. No studies have been done on the genetic status of the CN breed. This is important because improved goat breeds are increasingly being imported into Mexico and are used in indiscriminate crosses, threatening the CN breed’s genetic health13.
The evaluation of genetic diversity within and between breeds helps to understand a population’s genetic structure, and to establish strategies for conservation, genetic improvement and sustainable use of genetic resources14. Microsatellite molecular markers are useful in genetic characterization studies within and between populations. They provide genetic codominance, abundance, random distribution across the genome, high reproducibility, neutrality with respect to selection and high levels of polymorphism14,15. Numerous genetic diversity studies have been done recently on several cattle species using microsatellite markers and they have become the genetic markers of choice for molecular applications such as genetic diversity16,17, population structure18,19, phylogeny20, paternity evaluation21, etc. The present study objective was to evaluate genetic diversity and population structure of Criolla Negra goats using microsatellite markers.
Material and methods
Biological samples
Hair samples were collected from 45 individual goats distributed in three herds in three municipalities of the state of Querétaro, Mexico: Cadereyta de Montes (n= 7); El Marqués (n= 11); and San Juan del Río (n= 27). Samples were collected following the non-probabilistic opportunity method. Inclusion criteria were animals must not be related, they must be older than one year of age, have a black coat and erect or semi-lopped ears. Because genealogical data is unavailable for these populations kinship data provided by the producers was utilized.
The analysis included data for 25 microsatellites from 455 individuals from 13 goat populations: Retinta; Verata; Blanca Serrana; Celtibérica; Malagueña; Murciano-Granadina; Florida; Payoya; Serrana; Formentera; Saanen; Alpina; and Anglonubia. All populations form part of the Biodiversidad Caprina Iberoamericana (BioGoat) project22.
Molecular analysis
Extraction of DNA was done from the hair samples using a chelating resin (Chelex® 100, Bio-Rad Laboratories, Inc. USA)23. Thirty microsatellites recommended by the mixed ISAG/FAO committee for analysis of genetic diversity in domestic animals were used14. Of these, 25 were found to be held in common among the thirteen BioGoat populations. Marker amplification was done by polymerase chain reaction (PCR) using florescent primers24. The amplicons produced with the PCR were separated by capillary electrophoresis (ABI PRISM 3130 Genetic Analyzer, Applied Biosystems) following manufacturer instructions. Allele size was quantified with an internal size standard (GeneScan-400HD ROX, Applied Biosystems), and genotypes were identified with the GENOTYPER 2.5.1. software. Reference samples were included in each sample to confirm the results.
Statistical analysis
The total number of alleles per marker (NA) was determined by direct counts, while the median number of alleles (MNA) was calculated as the sum of all NA data divided by the number of makers used (n= 30). Observed heterozygosis (Ho) was calculated by dividing the number of individual heterozygotes in each marker by the number of individuals positive for each marker. Expected heterozygosis (He) was estimated with Nei’s formula25. Polymorphic data content, an indicator of marker quality26, was estimated using the MICROSATELLITE TOOLKIT complement for Microsoft Excel 201027. The number of effective alleles (NEA), which is the number of alleles able to pass to the following generation28, was generated with the POPGENE v. 1.32. software. The exact test of Hardy-Weinberg equilibrium (HWE), which considers heterozygote deficit, was calculated with the GENEPOP v.4.2 software29, using the thirty markers for the CN and the Markov chain method (5,000 dememorizations; 100 lots; 10,000 interactions per lot).
A 95%25,30 confidence interval was used when calculating endogamy coefficients for individuals versus subpopulations (FIS), individuals versus total population (FIT), and subpopulations versus total populations (FST), as well as the genetic differentiation coefficient (GST). All were generated with the GENETIX v. 4.05 software31.
A matrix for Reynolds genetic distance32, the minimum normalized Nei distance with a heterozygosis value in the founding population, was calculated with the POPULATIONS v.1.2.28 software. Split Graphs were then generated with the “NeighborNet” algorithm in the SPLITSTREE4 program33.
Genetic structure origin of the populations included in the study was analyzed with cluster (K) techniques, which represent the number of populations. These use a Bayesian algorithm employing a model based on the Montecarlo Markov Chains (MCMC) method, which estimates the a posteriori distribution of each mix coefficient for each individual. This was done with the STRUCTURE v.2.3.4 software34. The MCMC burn-in was 50,000 iterations and 200,000 repetitions, and results were viewed with the DISTRUCT program35. Optimum K was estimated by fixing values of K2 to K15 and running the analysis with fifteen repetitions for each K value, following the method of Evanno36 and using the STRUCTURE HARVESTER program37.
Results
Genetic variation in Criolla Negra population
A total of 243 alleles were identified with the thirty markers used in the analyses. Median number of alleles (MNA) was 8.1 per locus in this population (Table 1). The highest NA (13) was observed for markers MM12 and SRCRSP23, followed by BM6526 and HSC with twelve alleles. The lowest NA (2) was observed in MAF209. The HSC marker had the highest NEA (9.14) while MAF209 had the lowest (1.25). This may have occurred due to the proportion of polymorphic markers, the number of alleles per marker and their frequencies, and sample size. Average population He was 0.71, but varied from 0.20 in MAF209 to 0.90 in HSC. Average Ho was 0.62, and ranged from 0.18 in MAF209 to 0.93 in HSC.
Average PDC in the CN population was 0.66. The least informative marker (PDC<0.25) was MAF209 (PDC= 0.18), followed by ETH225 (PDC= 0.26) and SPS115 (PDC= 0. 44). The remaining 27 markers were the most informative (CIP >0.5). Fourteen of the thirty tested microsatellites exhibited significant deviation for the HWE (P≤0.05).
Table 1: Analyzed microsatellites, number of alleles detected (NA), number of effective alleles (NEA), expected heterozygosis (He), observed heterozygosis (Ho), polymorphic data content (PDC) and Hardy-Weinberg equilibrium (HWE) deviations
| Microsatellites | NA | NEA | He | Ho | PDC | HWE (P) |
|---|---|---|---|---|---|---|
| BM1329 | 8 | 3.44 | 0.72 | 0.67 | 0.68 | 0.20 |
| BM1818 | 8 | 4.63 | 0.79 | 0.71 | 0.76 | 0.12 |
| BM6506 | 9 | 3.26 | 0.70 | 0.55 | 0.66 | 0.02 |
| BM6526 | 12 | 4.93 | 0.81 | 0.86 | 0.78 | 0.90 |
| BM8125 | 6 | 3.64 | 0.73 | 0.62 | 0.68 | 0.02 |
| CRSM60 | 8 | 4.32 | 0.78 | 0.71 | 0.73 | 0.03 |
| CSRD247 | 6 | 3.25 | 0.70 | 0.77 | 0.66 | 0.97 |
| CSSM66 | 11 | 7.06 | 0.87 | 0.34 | 0.84 | 0.00 |
| ETH010 | 4 | 2.69 | 0.63 | 0.56 | 0.56 | 0.19 |
| ETH225 | 4 | 1.41 | 0.29 | 0.25 | 0.26 | 0.22 |
| HAUT27 | 8 | 4.11 | 0.77 | 0.71 | 0.72 | 0.28 |
| HSC | 12 | 9.14 | 0.90 | 0.93 | 0.88 | 0.31 |
| ILSTS011 | 6 | 2.87 | 0.66 | 0.59 | 0.59 | 0.33 |
| INRA063 | 5 | 2.46 | 0.60 | 0.64 | 0.51 | 0.72 |
| MAF065 | 11 | 5.78 | 0.84 | 0.87 | 0.81 | 0.24 |
| MAF209 | 2 | 1.25 | 0.20 | 0.18 | 0.18 | 0.43 |
| McM527 | 8 | 5.33 | 0.82 | 0.82 | 0.79 | 0.58 |
| MM12 | 13 | 7.00 | 0.87 | 0.75 | 0.84 | 0.03 |
| OarFCB011 | 10 | 5.73 | 0.83 | 0.75 | 0.80 | 0.12 |
| OarFCB048 | 10 | 7.14 | 0.87 | 0.78 | 0.84 | 0.01 |
| OarFCB304 | 10 | 3.72 | 0.74 | 0.61 | 0.69 | 0.00 |
| SPS115 | 3 | 2.02 | 0.51 | 0.27 | 0.44 | 0.00 |
| SRCRSP08 | 10 | 3.76 | 0.74 | 0.70 | 0.69 | 0.23 |
| TGLA122 | 8 | 2.20 | 0.55 | 0.49 | 0.52 | 0.02 |
| SRCRSP05 | 7 | 3.09 | 0.68 | 0.59 | 0.63 | 0.05 |
| SRCRSP23 | 13 | 8.49 | 0.89 | 0.68 | 0.87 | 0.00 |
| SRCRSP24 | 10 | 3.59 | 0.73 | 0.56 | 0.69 | 0.00 |
| ILSTS019 | 6 | 2.27 | 0.57 | 0.51 | 0.53 | 0.04 |
| INRA005 | 5 | 2.23 | 0.56 | 0.45 | 0.51 | 0.03 |
| INRA006 | 10 | 5.29 | 0.82 | 0.77 | 0.79 | 0.18 |
| Average | 8.1 | 4.20 | 0.71 | 0.62 | 0.66 |
P>0.05= Not significant
Interpopulation genetic differentiation
The endogamy coefficients (FIS, FIT and FST) and GST were estimated for each of the 25 microsatellites shared between the CN and the thirteen breeds included in BioGoat22 (Table 2). Average FIS was 0.067. Negative values for the markers BM8125 (-0.002) and MAF209 (-0.006) indicate these heterozygotes were present in excess18,19. Of the 25 markers eleven had a FIS greater than 0.05. Values for GST generally followed a similar trend to those for FST. The marker with the highest GST value was BM6526 (0.114) while the lowest was MAF209 (0.037).
Table 2 Genetic differentiation coefficient and endogamy coefficients for each microsatellite compared between the Criolla Negra breed and breeds in BioGoat (Retinta, Verata, Blanca Serrana, Celtibérica, Malagueña, Murciano-Granadina, Florida, Payoya, Serrana, Formentera, Saanen, Alpina and Anglonubia)
| Microsatellite | GST | FIS | FIT | FST |
|---|---|---|---|---|
| BM1329 | 0.081 | 0.024 | 0.078 | 0.056 |
| BM1818 | 0.077 | 0.024 | 0.085 | 0.062 |
| BM6506 | 0.074 | 0.044 | 0.094 | 0.053 |
| BM6526 | 0.114 | 0.045 | 0.111 | 0.069 |
| BM8125 | 0.076 | -0.002 | 0.065 | 0.067 |
| CRSM60 | 0.041 | 0.064 | 0.091 | 0.029 |
| CSRD247 | 0.082 | 0.026 | 0.095 | 0.071 |
| CSSM66 | 0.083 | 0.271 | 0.316 | 0.062 |
| ETH010 | 0.058 | 0.032 | 0.076 | 0.047 |
| ETH225 | 0.057 | 0.015 | 0.058 | 0.044 |
| HSC | 0.068 | 0.095 | 0.141 | 0.050 |
| ILSTS011 | 0.058 | 0.068 | 0.114 | 0.049 |
| INRA063 | 0.051 | 0.164 | 0.195 | 0.038 |
| MAF065 | 0.073 | 0.025 | 0.084 | 0.060 |
| MAF209 | 0.037 | -0.006 | 0.018 | 0.024 |
| McM527 | 0.081 | 0.087 | 0.151 | 0.070 |
| MM12 | 0.060 | 0.059 | 0.098 | 0.042 |
| OarFCB011 | 0.084 | 0.065 | 0.134 | 0.073 |
| OarFCB048 | 0.060 | 0.058 | 0.103 | 0.049 |
| SPS115 | 0.097 | 0.187 | 0.257 | 0.086 |
| SRCRSP08 | 0.106 | 0.049 | 0.143 | 0.099 |
| TGLA122 | 0.091 | 0.070 | 0.146 | 0.082 |
| Promedio | 0.073 | 0.067 | 0.121 | 0.058 |
GST= genetic differentiation coefficient; FIS= endogamy coefficient of individuals vs. subpopulations; FIT= endogamy coefficient of individuals vs. total population; and FST= endogamy coefficient of subpopulations vs. total population.
Interpopulation genetic distance and its graphic representation
Genetic distance analysis between the fourteen compared breeds showed the shortest distance to be between the CN and Murciano-Granadina breeds (MG) (0.133), and the longest to be between CN and Anglonubia (ANG) (0.420) (Table 3). A neighbor-net dendrogram was built to assist in interpreting values in the genetic distance matrix (Figure 1).
Table 3 Reynolds genetic distance matrix between the fourteen studied goat breeds
| RET | VERA | BLANCA | CELTIB | MALAG | MG | FLO | PAY | SER | FOR | SAAN | ALP | ANG | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| VERA | 0.025 | ||||||||||||
| BLANCA | 0.025 | 0.032 | |||||||||||
| CELTIB | 0.025 | 0.036 | 0.027 | ||||||||||
| MALAG | 0.021 | 0.025 | 0.023 | 0.021 | |||||||||
| MG | 0.044 | 0.045 | 0.031 | 0.034 | 0.041 | ||||||||
| FLO | 0.023 | 0.023 | 0.028 | 0.026 | 0.015 | 0.041 | |||||||
| PAY | 0.034 | 0.044 | 0.047 | 0.036 | 0.046 | 0.061 | 0.038 | ||||||
| SER | 0.034 | 0.038 | 0.035 | 0.028 | 0.024 | 0.048 | 0.027 | 0.045 | |||||
| FOR | 0.071 | 0.094 | 0.081 | 0.078 | 0.083 | 0.099 | 0.083 | 0.081 | 0.107 | ||||
| SAAN | 0.071 | 0.061 | 0.070 | 0.069 | 0.070 | 0.069 | 0.061 | 0.077 | 0.078 | 0.119 | |||
| ALP | 0.063 | 0.056 | 0.052 | 0.062 | 0.055 | 0.076 | 0.049 | 0.064 | 0.073 | 0.119 | 0.071 | ||
| ANG | 0.124 | 0.125 | 0.130 | 0.151 | 0.132 | 0.146 | 0.126 | 0.178 | 0.149 | 0.221 | 0.161 | 0.142 | |
| CN | 0.039 | 0.052 | 0.045 | 0.044 | 0.046 | 0.038 | 0.048 | 0.067 | 0.053 | 0.102 | 0.083 | 0.073 | 0.130 |
RET= Retinta, VERA= Verata, BLANCA= Blanca Serrana, CELTIB= Celtibérica, MALAG= Malagueña, MG= Murciano Granadina, FLO= Florida, PAY= Payoya, SER= Serrana, FOR= Formentera, SAAN= Saanen, ALP= Alpina, ANG= Anglonubia, CN= Criolla negra.
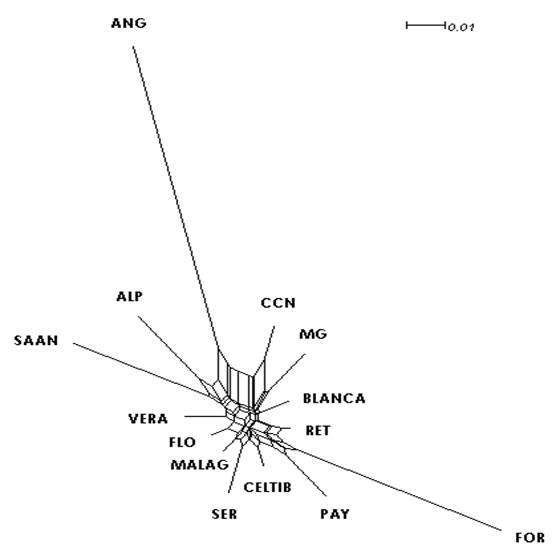
RET= Retinta, VERA= Verata, BLANCA= Blanca Serrana, CELTIB= Celtibérica, MALAG= Malagueña, MG= Murciano-Granadina, FLO= Florida, PAY= Payoya, SER= Serrana, FOR= Formentera, SAAN= Saanen, ALP= Alpina, ANG= Anglonubia, CCN= Criolla Negra.
Figure 1 Neighbor-net dendrogram built using Reynolds genetic distance data for the fourteen studied goat breeds.
Genetic structure analysis
Optimum K for the genetic structure of the studied populations was 9. When shown graphically (Figure 2), each individual is represented by a vertical line divided into color segments indicating to what extent the individual belongs to each group (K).
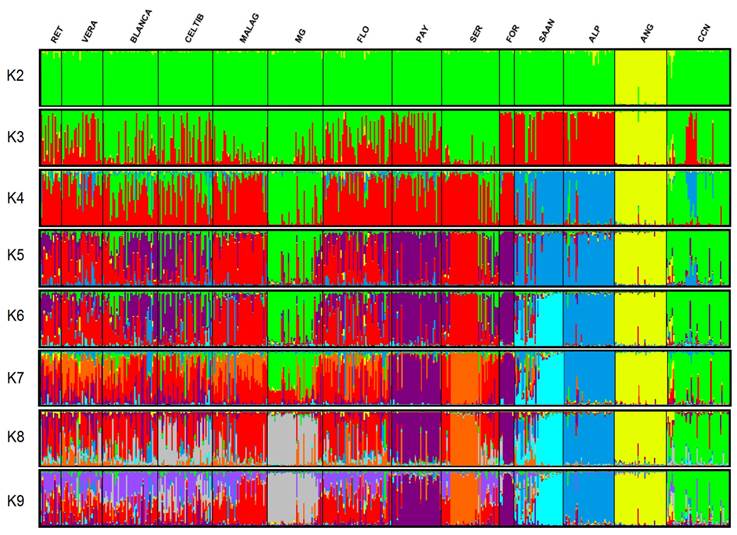
RET= Retinta, VERA= Verata, BLANCA= Blanca Serrana, CELTIB= Celtibérica, MALAG= Malagueña, MG= Murciano-Granadina, FLO= Florida, PAY= Payoya, SER= Serrana, FOR= Formentera, SAAN= Saanen, ALP= Alpina, ANG= Anglonubia, CCN= Cabra Criolla Negra.
Figure 2 Graphic representation of genetic structure in the fourteen studied goat breeds, assuming an ancestral populations number ranging from 2 to 9
Discussion
Genetic variation in Criolla Negra population
When running analyses with the microsatellites used in the present study it is recommended to have at least four alleles, and that the NEA be greater than two to be included in diversity studies and reduce standard error when estimating genetic distances38. Only two markers (MAF209 and SPS115) had 2 and 3 alleles in the present results. Values for NEA were lower than two in the markers ETH225 (1.41) and MAF209 (1.25). This indicates how appropriate the markers were for genetic diversity evaluation of NA and NEA. The MNA value for the CN population (8.1) provided information on genetic diversity in this population. When MNA values are higher diversity is greater and vice versa. For the studied CN population MNA was high compared to other characterization studies done with breeds such as the Criollo Cubano39; Saudi goats19; cashmere goats in China40,41, and some Iranian goat breeds42. However, the MNA values for CN were similar to those reported for dairy goats in South Africa43. Average PDC value for the markers used here was 0.66, which is similar to values reported for the Retinta Extremeña goat breed44.
Heterozygosity values can also help to understand genetic diversity since they depend on allele number and relative frequency45. The average heterozygosis values in the present results (He = 0.71, Ho = 0.62) are very similar to those reported for the Blanca Andaluza46 and Retinta Extremeña goat breeds44, though the latter has a higher MNA than the CN. These He and Ho values indicate the presence of notable genetic variability in the CN considering that the population is shrinking in the studied area, which would be expected to promote marked consanguinity. The HWE test identified fourteen markers with significant deviations (P≤0.05), indicating there to be a heterozygous deficit. The fact that some of these markers behave homozygously may be due to actions such as management conditions (e.g. sire loan), low genetic flow in each flock, and markers that could be linked to productive traits due to production-focused selection of milk and weight gain traits regardless of kinship relations47.
Interpopulation genetic differentiation
Estimated values for FIS and FIT vary from 1 to -1, with positive values indicating heterozygote deficiency and negative values an excess. The present results for both indices (FIS= 0.067 and FIT= 0.121) indicate that some of the markers were homozygous. Although the values were near zero, they indicate possible mating between genetic relatives, which is consistent with values reported for native goats in China48, India47, Spain and Portugal24. The FST indicated that 94.2% of the genetic variability in the studied breeds was due to differences between individuals within the breed and 5.8% due to genetic differences between breeds. This genetic differentiation coefficient (GST = 0.073) confirmed this result in that it showed 92.7% of variability of total genetic diversity to be intrabreed and 7.3% to be interbreed. The discrepancy between these indicators exists because FST reflects the properties of interpopulation allelic frequency distribution while GST is defined in terms of intrapopulation frequencies49. Both the FST and GST values suggest that the level of genetic variation in the studied breeds has remained relatively constant. Percentages like those in the present study have been reported for other goat populations17,42,50.
Interpopulation genetic distance and its graphic representation
The neighbor-net dendrogram showed that the Spanish goat breeds included in the study (Retinta, Verata, Blanca Serrana, Celtibérica, Malagueña, Murciano-Granadina, Florida, Payoya, Serrana and Formentera) remain grouped. Reported in previous studies51, this effect is caused by the close genetic and geographical relationships between these breeds. The Saanen and Alpina breeds formed another group towards one end of the dendrogram. Of particular note is the Anglonubia breed’s large genetic distance from the other studied breeds. These kinds of relationships have also been observed in a comparison between Brazilian goat breeds52. This effect can be attributed to a greater genetic distance between a breed when compared to others and not necessarily to origin or kinship relations. Another important factor in any goat population is that individuals within it have also been selected based on morphological characteristics. Estimates generated from the Reynolds genetic distance data showed the shortest distance (0.038) to be between the CN and MG breeds, suggesting a possible genetic relationship between them.
Genetic structure analysis
Genetic structure analysis was used to evaluate the degree of kinship between the different studied populations, using optimum K (K= 9) to identify interbreed differences. No crossings were found between CN and the other studied breeds. The ANG population separated from the others beginning at K2 and remained so thereafter. Apparently, this population is more genetically distant from the other populations, a phenomenon reported for the population structure of other Creole goats in the Americas53. The ANG population also preserved its genetic structure, having a low level of mixing of individuals from the other thirteen studied populations. Genetic structure in the Spanish breeds included in this study was more intermixed, similar to the results reported in a study on goat biodiversity24. Intermixing is often due to geographical proximity between populations, which facilitates migration of individuals between populations. The genetic nearness of the CN breed to the MG breed confirm the supposed origin of the studied CN population. However, analysis using optimal K (K9) showed the CN population to have a totally different structure than the MG breed and the other studied breeds. This supports the distances shown in the neighbor-net dendrogram and suggests that the CN breed maintains a unique genetic structure that is differentiated from the populations that may have contributed to its origin.
Conclusions and implications
These are the first published data on genetic diversity and structure in a Criolla Negra goat population. The studied population has a certain degree of genetic diversity based on its level of polymorphism. The genetic distances between the Criolla Negra population and the other races included in the study indicate that this population is clearly differentiated from them and should thus be considered a distinct Mexican goat breed. The relatively short genetic distance between the Murciano-Granadina and Criolla Negra breeds suggests that both have a common ancestor, most probably the Granadina breed. The Criolla Negra goat has a defined breed structure and is differentiated from its possible precursor breeds. The Criolla Negra breed is the first breed of goat in central Mexico to be genetically described.
Literatura citada
1. Naderi S, Rezaeli HR, Taberlet P, Zundel S, Rafat S, Naghash H, et al. Large-Scale Mitochondrial DNA Analysis of the domestic goat reveals six haplogroups with high diversity. PLoS One 2007;2:1-12. [ Links ]
2. Zeder MA, Hesse B. The initial domestication of goats (Capra hircus) in the Zagros Mountains 10,000 years ago. Science 2000;287:2254-2257. [ Links ]
3. Nomura K, Yonezawa T, Mano S, Kawakami S, Shedlock AM, Hasegawa M, et al. Domestication process of the goat revealed by an analysis of the nearly complete mitochondrial protein-encoding genes. PLoS One 2013;8:1-15. [ Links ]
4. Luikart G, Gielly L, Excoffier L, Vigne J, Bouvet J, Taberlet P. Multiple maternal origins and weak phylogeographic structure in domestic goats. Proc Natl Acad Sci USA 2001;98:5927-5932. [ Links ]
5. Rodero A, Delgado JV, Rodero E. Primitive andalusian livestock and their implications in the discovery of America. Arch Zootec 1992;41:383-400. [ Links ]
6. FAO. Statistics Database. FAOSTAT. 2017 (Revisado 2017/12/15). Disponible en: Disponible en: http://www.fao.org/faostat/es/#data/QA . [ Links ]
7. Peacock C, Sherman DM. Sustainable goat production-Some global perspectives. Small Ruminant Res 2010;89:70-80. [ Links ]
8. FAO. La situación de los recursos zoogenéticos mundiales para la alimentación y la agricultura. 2010. Disponible en: http://www.fao.org/docrep/012/a1250s/ a1250s00.htm [ Links ]
9. SAGARPA. Informe sobre la Situación de los Recursos Genéticos Pecuarios en México. 2002. Disponible en: http://www.sagarpa.gob.mx/ganaderia/Publicaciones/ Paginas/InfoRGPecuariosM.aspx [ Links ]
10. Montaldo HH, Meza-Herrera CA. Genetic goat resources in Mexico: Bio- economical efficiency of local and specialized genotypes. Wool Technol Sheep Breed 1999;47:184-198. [ Links ]
11. Montaldo H, Jutiezb A, Berruecosc JM, Sinchezd F. Performance of local goats and their backcrosses with several breeds in Mexico. Small Ruminant Res 1995;16:97-105. [ Links ]
12. Montaldo H, Tapia G, Juárez A. Algunos factores genéticos y ambientales que influyen sobre la producción de leche y el intervalo entre partos en cabras. Tec Pecu Mex 1981;41:32-44. [ Links ]
13. Montaldo H, Torres-Hernández G, Valencia-Posadas M. Goat breeding research in Mexico. Small Ruminant Res 2010;89:155-163. [ Links ]
14. FAO. Molecular genetic characterization of animal genetic resources. FAO Anim Prod Heal Guid. Rome; 2011. [ Links ]
15. Baumung R, Simianer H, Hoffmann I. Genetic diversity studies in farm animals - A survey. J Anim Breed Genet 2004;121:361-373. [ Links ]
16. Jamil T, Antonella A, Philippe B, Roswitha B, Albano B, Marco B, et al. Genetic diversity of Albanian goat breeds based on microsatellite markers. Arch Zootec 2011;60:607-615. [ Links ]
17. Li JY, Chen H, Lan XY, Kong XJ, Min LJ. Genetic diversity of five Chinese goat breeds assessed by microsatellite markers. Czech J Anim Sci 2008;53:315-319. [ Links ]
18. Thuy LT, Binh D Van, Binh NT, Minh LQ, Thuy TTT, Ton ND, et al. Evaluation of genetic diversity and structure of Vietnamese goat populations using multi locus microsatellite markers. Small Ruminant Res 2017;148:43-50. [ Links ]
19. Al-Atiyat RM, Alobre MM, Aljumaah RS, Alshaikh MA. Microsatellite based genetic diversity and population structure of three Saudi goat breeds. Small Ruminant Res 2015;130:90-94. [ Links ]
20. Rout PK, Joshi MB, Mandal A, Laloe D, Singh L, Thangaraj K. Microsatellite-based phylogeny of Indian domestic goats. BMC Genet 2008;9:11. [ Links ]
21. Mello A, Facioni S, Silva C, Sávio P, Teixeira M, Mendeiros T. Paternity in Brazilian goats through the use of DNA microsatellites. Rev Bras Zootec 2010;39:1011-1014. [ Links ]
22. BioGoat Consortium. Proyecto Biodiversidad Caprina Iberoamericana. Disponible en: https://biogoat.jimdo.com/ [ Links ]
23. Walsh PS, Metzger DA, Higuchi R. Chelex 100 as a medium for simple extraction of DNA for PCR-based typing from forensic material. Biotechniques 2013;54:134-139. [ Links ]
24. Martínez AM, Gama LT, Delgado JV, Cañón J, Amills M, Bruno de Sousa C, et al. The Southwestern fringe of Europe as an important reservoir of caprine biodiversity. Genet Sel Evol 2015;47:86. [ Links ]
25. Nei M. Analysis of gene diversity in subdivided populations. Proc Nat Acad Sci 1973;70:3321-3323. [ Links ]
26. Botstein D, White RL, Skolnick M, Davis RW. Construction of a genetic linkage map in man using restriction fragment length polymorphisms. Am J Hum Genet 1980;32:314-331. [ Links ]
27. Park SDE. Trypanotolerance in West African cattle and the population genetic effects of selection [Ph.D. Thesis]. Dublin, Ireland: University of Dublin; 2001. [ Links ]
28. Kimura M, Crow JF. The number of alleles that can be maintained in a finite population. Genetics 1964;49:725-738. [ Links ]
29. Raymond M, Rousset F. GENEPOP (Version 1.2): Population genetics software for exact tests and Ecumenicism. J Hered 1995;86:248-249. [ Links ]
30. Nei M. F-statistics and analysis of gene diversity in subdivided populations. Ann Hum Genet 1977;41:225-233. [ Links ]
31. Belkhir K, Borsa P, Chikhi L, Raufaste N, Bonhomme F. GENETIX 4.05, logiciel sous Windows TM pour la génétique des populations. Laboratoire Génome, Populations, Interactions, CNRS UMR 5000, Université de Montpellier II, Montpellier (France). 1996. [ Links ]
32. Reynolds J, Weir BS, Cockerham CC. Estimation of the coancestry coefficient: basis for a short-term genetic distance. Genetics 1983;105:767-779. [ Links ]
33. Huson DH, Bryant D. Application of phylogenetic networks in evolutionary studies. Mol Biol Evol 2006;23:254-267. [ Links ]
34. Pritchard JK, Stephens M, Donnelly P. Inference of population structure using multilocus genotype data. Genetics 2000;155:945-959. [ Links ]
35. Rosenberg NA. DISTRUCT : a program for the graphical display of population structure. Mol Ecol 2004;4:137-138. [ Links ]
36. Evanno G, Regnaut S, Goudet J. Detecting the number of clusters of individuals using the software STRUCTURE : a simulation study. Mol Ecol 2005;14:2611-2620. [ Links ]
37. Earl DA, Bridgett M. STRUCTURE HARVESTER : a website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv Genet Resour 2012;4:359-361. [ Links ]
38. Barker JSF. A global protocol for determining genetic distances among domestic livestock breeds. 5th World Cong Genet Appl Livest Prod. Canadá; 1994:501-508. [ Links ]
39. Chacón E, Martínez A, La OM, Velázquez FJ, Pérez E, Vicente-Delgado J. Caracterización genética de la cabra Criolla Cubana mediante marcadores microsatélites. Rev Cub Cienc Agr 2010;44:221-226. [ Links ]
40. Di R, Farhad Vahidi SM, Ma YH, He XH, Zhao QJ, Han JL, et al. Microsatellite analysis revealed genetic diversity and population structure among Chinese cashmere goats. Anim Genet 2011;42:428-431. [ Links ]
41. Du X, Cao J, Han X, Hao H, Yu M, Zhang G, et al. Genetic diversity and population structure among eight Chinese indigenous goat breeds in the Yellow River valley. Small Ruminant Res 2017;148:87-92. [ Links ]
42. Vahidi SMF, Tarang AR, Naqvi AN, Falahati Anbaran M, Boettcher P, Joost S, et al. Investigation of the genetic diversity of domestic Capra hircus breeds reared within an early goat domestication area in Iran. Genet Sel Evol 2014;46:1-12. [ Links ]
43. Bosman L, van Marle-Köster E, Visser C. Genetic diversity of South African dairy goats for genetic management and improvement. Small Ruminant Res 2015;123:224-231. [ Links ]
44. Parejo JC, Padilla JA, Calero R, Martínez-Trancón M, Sansinforiano E, Rabasco A, et al. Population genetic structure and conservation management of Retinta Extremeña goats. Small Ruminant Res 2015;124:9-16. [ Links ]
45. Hale ML, Burg TM, Steeves TE. Sampling for microsatellite-based population genetic studies: 25 to 30 individuals per population is enough to accurately estimate allele frequencies. PLoS One 2012;7:1-10. [ Links ]
46. Martínez AM, Carrera MP, Acosta JM, Rodríguez-Gallardo PP, Cabello A, Camacho E, et al. Genetic characterization of the Blanca Andaluza goat based on microsatellite markers. South African J Anim Sci 2004;34:17-19. [ Links ]
47. Dixit SP, Verma NK, Aggarwal RAK, Vyas MK, Rana J, Sharma A. Genetic diversity and relationship among Indian goat breeds based on microsatellite markers. Small Ruminant Res 2012;105:38-45. [ Links ]
48. Cao J, Li X, Du X, Zhao S. Microsatellite based genetic diversity and population structure of nine indigenous Chinese domestic goats. Small Ruminant Res 2017;148:80-86. [ Links ]
49. Holsinger KE, Weir BS. Genetics in geographically structured populations: defining, estimating and interpreting Fst. Nat Rev Genet 2009;10:639-650. [ Links ]
50. Martínez AM, Acosta J, Vega-Pla JL, Delgado JV. Analysis of the genetic structure of the canary goat populations using microsatellites. Livest Sci 2006;102:140-145. [ Links ]
51. Cañón J, García D, García-Atance MA, Obexer-Ruff G, Lenstra JA, Ajmone-Marsan P, et al. Geographical partitioning of goat diversity in Europe and the Middle East. Anim Genet 2006;37:327-334. [ Links ]
52. Oliveira JD De, Luiza M, Paiva S De, Mírian T, Machado M, Miretti MM, et al. Structure and genetic relationships between Brazilian naturalized and exotic purebred goat domestic goat (Capra circus) breeds based on microsatellites. Genet Mol Biol 2007;30:356-363. [ Links ]
53. Ginja C, Gama LT, Mart A, Sevane N, Lanari MR. Genetic diversity and patterns of population structure in Creole goats from the Americas. Anim Genet 2017;1-15. [ Links ]
Received: May 19, 2018; Accepted: October 22, 2018
 Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
