











 text in
text in  English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkIntroducción
El tomate (Solanum lycopersicum L.) es la hortaliza de mayor preferencia a nivel mundial (Infoagro, 2017). Presenta altos contenidos de compuestos bioactivos como: folato, ascorbato, polifenoles, carotenoides, vitaminas y otros nutrientes esenciales, por lo cual es apetecible para la salud humana (Ahmed et al., 2017). FIRA (2019) menciona que el tomate a nivel mundial es cultivado aproximadamente en cinco millones de hectáreas (ha), con una producción de 170.08 millones de toneladas (t) de tomate fresco, donde China ocupa el primer lugar en el mundo con una producción de tomate de 46.2% del total, mientras que México ocupa el décimo lugar con una producción de 1 878 289 t anuales.
La salinidad afecta a 1 125 millones de ha en todo el mundo (Hossain, 2019). Esta condición genera problemas que la agricultura enfrenta a nivel mundial, particularmente por la presencia de NaCl, la cual limita el óptimo desarrollo de las plantas de interés comercial (Durukan y Demirbas 2018). México, posee una superficie de 54% de zonas áridas en todo el territorio nacional, ubicadas principalmente en las regiones norte y noreste del país (Briones et al., 2018), las cuales están afectadas con un alto grado de salinidad, donde los cationes de sodio (Na+) desplazan el calcio (Ca+) del complejo arcillo-húmico, incrementando así, el potencial osmótico de la solución del suelo (Cisneros et al., 2020).
En la actualidad se buscan diversas alternativas para mitigar los efectos negativos de la salinidad, como la obtención de variedades tolerantes, productos naturales como biofertilizantes y bioestimulantes que favorezcan el crecimiento y desarrollo de las plantas y beneficien al medio ambiente (Mazón et al., 2020).
El Si es un mineral que protege a la planta contra el estrés biótico y abiótico; además, el Si químicamente activo restaura el daño y optimiza la fertilidad del suelo, mantiene los nutrientes en forma disponible para la planta, mejora la capacidad de intercambio catiónico y pH mayor a 7 (Meunier et al., 2018). Por lo tanto, el objetivo fue evaluar el efecto del Si como atenuante de la salinidad en la germinación y crecimiento inicial en plántulas de tres variedades de tomate.
Materiales y métodos
La investigación se realizó en el laboratorio de germoplasma de la Universidad Autónoma de Baja California Sur (UABCS). El material biológico utilizado fue tres cultivares de tomate: cherry (Solanum lycopersicum var. Cerasiforme), bola (Solanum lycopersicum var. Floradade) y saladette (Solanum lycopersicum var. Río grande), la variedad bola y saladette son variedades comerciales, mientras que el cherry es un criollo reproducido en la Universidad Autónoma de Baja California Sur. Las pruebas de germinación se realizaron en cajas de Petri (150 x 15 mm) esterilizadas, cubiertas del fondo con una capa de papel absorbente. Cada caja fue humedecida con 5 ml de la solución resultante de la combinación de cloruro de sodio (NaCl) (0, 25 y 50 mM) y dióxido de silicio amorfo (SiO2) (0, 1 y 2 mM), la aplicación de la solución se realizó cada cuatro días, para el tratamiento control se utilizó agua destilada. Las pruebas de germinación se realizaron durante 14 días, en una cámara de germinación (Seedburo, Model 549, NY, USA) a una temperatura de 25 °C ±0.5 °C y 80% de humedad relativa, las cuales se estuvieron monitoreando diariamente.
Variables evaluadas
Porcentaje de germinación. Se registró diariamente utilizando la siguiente fórmula de Al-mudaris (1998). Donde:
Las semillas se consideraron germinadas cuando la radícula presentó una longitud mínima de 2 mm. La Tasa de germinación, se calculó usando la fórmula de Maguire (1962), donde n1, n2, n30 son el número de semillas germinadas en los tiempos t1, t2, t7 (en días). Mientras que la energía de germinación se calculó usando la siguiente fórmula de Maguire (1962):
Donde: N= número de semillas germinadas en la fecha de conteo; D= número de días. A su vez el tiempo medio de germinación, se realizó por la fórmula propuesta por Orchard (1977)
Donde: N= número de semillas germinadas en el día D.
Por su parte el índice de germinación se calculó por la fórmula propuesta por Scott et al. (1962),
Donde: n= número de semillas germinadas en el día 1; t1= número de días después de la siembra; N= total de semillas sembradas. Mientras que la velocidad de germinación se determinó por la fórmula de Maguire (1962),
Donde: n= número de semillas germinadas en el día 1; t= tiempo de germinación desde la siembra hasta la germinación de la última semilla.
Las semillas germinadas se mantuvieron por 14 días y se seleccionaron al azar 10 plántulas por repetición. A cada plántula se le midió longitud de tallo y radícula con un pie de rey digital (AutoTec, Rhos, 2002 EC). Posteriormente se pesó la biomasa fresca y seca, utilizando una balanza analítica (Ohaus, modelo PA224C). Los tejidos vegetales se colocaron en bolsas de papel y se introdujeron en una estufa de secado (Ríos Rocha modelo EO-50) a una temperatura de 70 °C durante 72 h hasta su deshidratación completa.
Diseño experimental
El diseño experimental fue completamente al azar con arreglo factorial de 32, donde el factor A fueron las concentraciones salinas de NaCl (0, 25 y 50 mM) y el factor B las diluciones de Si (0, 1 y 2 mM). Cada tratamiento contó con cuatro repeticiones de 25 semillas cada una.
Análisis estadístico
A los datos obtenidos se les realizó el análisis de varianza. En las variables donde se mostró diferencia significativa se realizó una prueba de polinomios ortogonales. Las diferencias entre las medias de tratamientos fueron comparadas con la prueba de rango múltiple de Tukey (p≤ 0.05) de nivel de confianza. Los análisis estadísticos se realizaron con el programa Statistical Package for the Social Sciences (SPSS) versión 22.0 (IBM Corp, 2013).
Resultados y discusión
El análisis estadístico mostró que, el efecto del NaCl, el Si y la interacción entre estos factores fue estadísticamente diferente (p≤ 0.05) en cada variedad de tomate (Cuadro 1).
Cuadro 1 Valores de F y niveles de significancia (p) observados en los análisis de varianza para las variables de germinación y crecimiento.
| Variables | Cherry | Floradade | Río grande | ||||||||
| NaCl | Si | NaCl*Si | NaCl | Si | NaCl*Si | NaCl | Si | NaCl*Si | |||
| %GP | |||||||||||
| F | 3.737 | 0.712 | 1.017 | 1.004 | 0.049 | 1.25 | 2.497 | 0.113 | 1.061 | ||
| p | 0.037 | 0.5 | 0.416 | 0.38 | 0.952 | 0.314 | 0.101 | 0.893 | 0.395 | ||
| GE | |||||||||||
| F | 12.339 | 2.932 | 1.624 | 0.454 | 0.125 | 1.309 | 1.5 | 0.001 | 0.891 | ||
| p | <0.001 | 0.07 | 0.197 | 0.64 | 0.883 | 0.292 | 0.241 | 0.999 | 0.483 | ||
| GRI | |||||||||||
| F | 11.124 | 9.312 | 6.387 | 5.562 | 8.677 | 2.558 | 7.926 | 2.138 | 1.637 | ||
| p | <0.001 | <0.001 | <0.001 | 0.009 | <0.001 | 0.061 | 0.002 | 0.137 | 0.194 | ||
| MGT | |||||||||||
| F | 3.154 | 0.673 | 1.051 | 0.785 | 0.06 | 0.936 | 3.052 | 0.031 | 1.054 | ||
| p | 0.059 | 0.519 | 0.399 | 0.466 | 0.942 | 0.458 | 0.064 | 0.969 | 0.398 | ||
| IG | |||||||||||
| F | 7.903 | 1.144 | 0.917 | 1.004 | 0.049 | 1.25 | 2.497 | 0.113 | 1.061 | ||
| p | 0.002 | 0.333 | 0.468 | 0.38 | 0.952 | 0.314 | 0.101 | 0.893 | 0.395 | ||
| VG | |||||||||||
| F | 3.737 | 0.712 | 1.017 | 1.004 | 0.049 | 1.25 | 2.497 | 0.113 | 1.061 | ||
| p | 0.037 | 0.5 | 0.416 | 0.38 | 0.952 | 0.314 | 0.101 | 0.893 | 0.395 | ||
| LR | |||||||||||
| F | 8.301 | 2.098 | 6.109 | 4.621 | 5.434 | 1.571 | 13.076 | 10.335 | 0.667 | ||
| p | 0.002 | 0.142 | <0.001 | 0.019 | 0.01 | 0.211 | <0.001 | <0.001 | 0.62 | ||
| LT | |||||||||||
| F | 222.456 | 119.31 | 38.123 | 8.786 | 12.858 | 5.975 | 33.489 | 10.317 | 2.152 | ||
| p | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | 0.102 | ||
| BF | |||||||||||
| F | 15.374 | 1.508 | 0.786 | 18.816 | 4.517 | 3.268 | 78.338 | 12.499 | 4.93 | ||
| p | <0.001 | 0.239 | 0.544 | <0.001 | 0.02 | 0.026 | <0.001 | <0.001 | 0.004 | ||
| BS | |||||||||||
| F | 16.487 | 4.737 | 7.643 | 6.09 | 0.89 | 0.578 | 4.454 | 3.794 | 0.588 | ||
| p | <0.001 | 0.017 | <0.001 | 0.007 | 0.422 | 0.681 | 0.021 | 0.035 | 0.674 | ||
Porcentaje de germinación (%GP); energía de germinación (GE); tasa de germinación (GRI); tiempo medio de germinación (MGT); índice de germinación (IG); velocidad de germinación (VG); longitud de raíz (LR); longitud de tallo (LT); biomasa fresca (BF) y biomasa seca (BS).
Para la variedad cherry, la concentración de 50 mM NaCl incrementó la energía de germinación y el índice de germinación. Mientras que, la biomasa fresca se incrementó en las concentraciones de 25 y 50 mM NaCl. Sin embargo, el porcentaje de germinación, energía de germinación, índice de germinación y la velocidad de germinación se vieron afectados en la concentración de 25 mM de NaCl, con respecto al tratamiento control (Cuadro 2).
Cuadro 2 Efectos principales de las concentraciones de NaCl y del Si en las variables fisiológicas de germinación de tomate cherry (Solanum lycopersicum var. Cerasiforme).
| Dosis | Variables evaluadas | |||||
| % GP | GE | MGT | IG | VG | BF | |
| NaCl | ||||||
| 0 mM | 89.667 a | 11.716 ab | 0.219 | 0.152 a | 0.897 a | 1.41 b |
| 25 mM | 82.667 b | 10.723 b | 0.207 | 0.14 b | 0.827 b | 1.47 ab |
| 50 mM | 89.667 a | 13.002 a | 0.219 | 0.161 a | 0.897 a | 1.67 a |
| ±EE | 2.091 | 0.325 | 0.004 | 0.004 | 0.021 | 0.001 |
| Lineal | ns | 0.009 | ns | ns | ns | < 0.001 |
| Cuadrática | 0.011 | < 0.001 | ns | 0.001 | 0.009 | 0.012 |
| Si | ||||||
| 0 mM | 86.667 | 12.348 | 0.215 | 0.154 | 0.867 | 1.6 |
| 1 mM | 89.333 | 11.855 | 0.218 | 0.152 | 0.893 | 1.48 |
| 2 mM | 86 | 11.237 | 0.212 | 0.147 | 0.86 | 1.47 |
| ±EE | 2.091 | 0.325 | 0.004 | 0.004 | 0.021 | 0.001 |
| Lineal | ns | ns | ns | ns | ns | ns |
| Cuadrática | ns | ns | ns | ns | ns | ns |
Literales diferentes entre las medias indican diferencia significativa. Porcentaje de germinación (%GP); energía de germinación (GE); tiempo medio de germinación (MGT); índice de germinación (IG); velocidad de germinación (VG); biomasa fresca (BF); NS= no significativo; ±E= error estándar.
Cuartero et al. (2006) informaron que el género lycopersicum posee un carácter heredable de tolerancia a la salinidad. Vítámvá et al. (2007) mencionaron que existen un grupo de proteínas inducibles por el ácido abscísico (ABA), las cuales están asociadas con la tolerancia a la salinidad y el déficit hídrico, lo cual podría explicarse a lo sucedido en el presente trabajo.
Diversos estudios muestran, que se han encontrado incrementos en el porcentaje, la energía y el índice de germinación en concentraciones de 50 mM NaCl en cultivos como: espinaca Spinacia oleracea (Turhan et al., 2011), tomate solanum lycopersicum (Doğan et al., 2008) y berenjena Solanum melongena (Akıncı et al., 2004).
Por otra parte, Eitel (2021) evaluó el efecto del estrés salino en la producción y calidad de semillas de tomate, en distintas concentraciones, donde se observó que el índice de germinación disminuyó significativamente. Por otro lado, Ruiz et al. (2014) evaluaron la germinación de ocho genotipos de tomate (Solanum lycopersicum L.), los cuales fueron expuestos a diferentes niveles de NaCl. Donde observaron que la biomasa fresca y seca de la parte aérea se incrementaba en presencia de NaCl. De la misma manera, Can et al. (2017) evaluaron la germinación y crecimiento inicial de frijol (Phaseolus vulgaris) en respuesta a la salinidad inducida con NaCl, sulfato de sodio (Na2SO4) y bicarbonato de sodio (NaHCO3), donde observaron que la biomasa fresca fue disminuida con el tipo de sal NaHCO3, mientras que, NaCl la incrementó.
A pesar de que no se indicó el impacto por parte de Si como factor principal, sí se mostró efecto significativo en la interacción con NaCl. La presencia de Si en la tasa de germinación señala un efecto cuadrático en la presencia de NaCl (Figura 1A). Resultados similares fueron encontrados por Haghighi et al. (2012), quienes evaluaron el factor interacción de Si + NaCl en la germinación de semillas de tomate, donde observaron que, el Si estimuló las variables germinativas, en especial la tasa de germinación, la cual incrementó alrededor de 20% con respecto al tratamiento control. De la misma manera, Sun et al. (2021) observaron el efecto del Si en la germinación de maíz (Zea mays) la cual aumentó significativamente el porcentaje de germinación (GP), tasa de germinación (GR), índice de germinación (GI) e índice de vigor (VI) en comparación para el tratamiento control.
La presencia de Si en condiciones salinas (50 mM) generó un incremento en la longitud de la raíz (Figura 1B). Resultados similares fueron reportados por Asgari et al. (2018) quienes encontraron un mayor incremento en la longitud del tallo, biomasa fresca y saca de los brotes y raíces en plantas de avena tratadas (Avena sativa) tratadas con Si.
Sin embargo, Gong et al. (2006) evaluaron la interacción de Si + NaCl en plántulas de arroz (Oryza sativa). Donde observaron que el Si no mitigó los efectos adversos de NaCl. En el desarrollo del tallo se observó un efecto positivo del Si (Figura 1C), particularmente las plántulas en condiciones salinas (50 mM) incrementaron la longitud del tallo con presencia de Si. Mientras que, en la producción de biomasa seca (Figura 1D) se observó un impacto positivo del Si en situación no salina, mostrando un efecto cuadrático. Haghighi y Pessarakli (2013) evaluaron el factor interacción de Si + NaCl en tomate cherry. Donde observaron que, el Si mitigó los efectos adversos de NaCl e incrementó la longitud del tallo, biomasa fresca y seca, tasa fotosintética y contenido de agua.
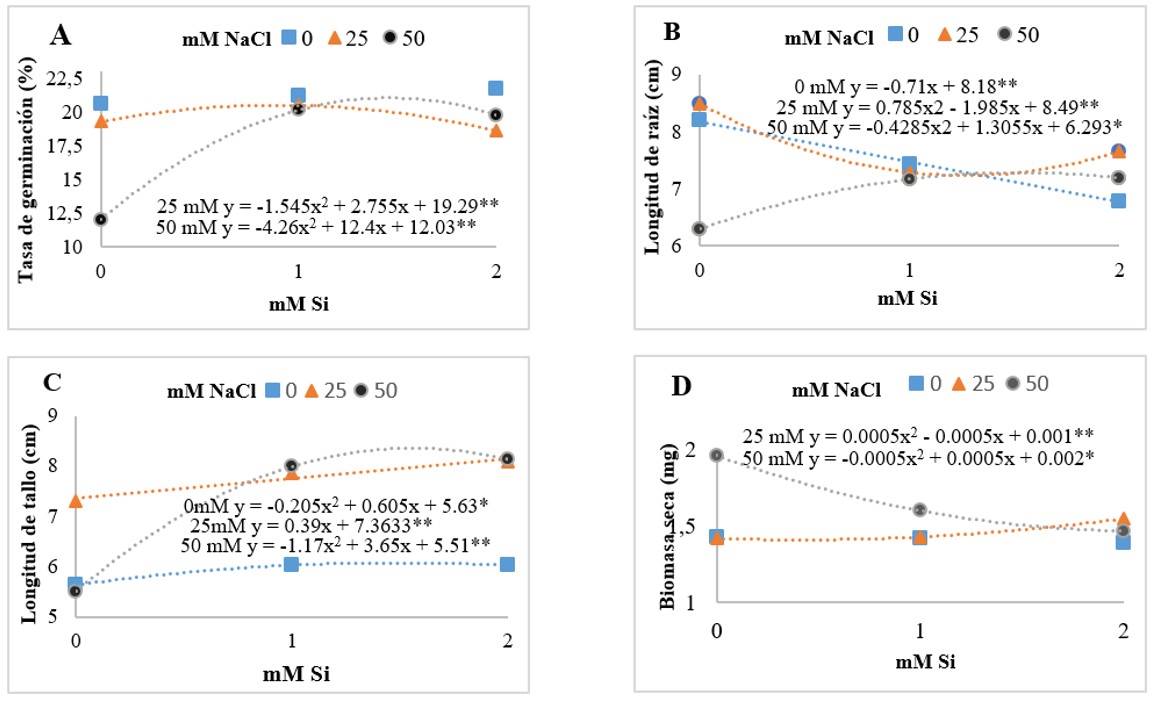
Figura 1. Efecto de la interacción de cloruro de sodio (NaCl) y silicio (Si) en la tasa de germinación (A); longitud de raíz (B); longitud de tallo (C); y biomasa seca (D) de plántulas de tomate cherry después de los 14 días del establecimiento del experimento. Los asteriscos indican los niveles de significancia entre los tratamientos 0.01** - 0.05*.
Wang et al. (2015) observaron que el Si promovió el incremento de la longitud del tallo, raíz y la biomasa fresca de las plantas de pepino (Cucumis sativus) sometidas a estrés por NaCl. Por otra parte, Carballo et al. (2019) evaluaron el factor interacción de Si + NaCl en la germinación de moringa (Moringa oleífera). Dichos resultados, mostraron un efecto positivo del Si al incrementar la biomasa, índice de vigor, fenoles totales, flavonoides totales y la capacidad antioxidante.
En la variedad Floradade se observó que la concentración de 50 mM de NaCl redujo la tasa de germinación en 14%. Contrario a lo anterior, la concentración de 25 mM NaCl incrementó en 19% la longitud de la raíz, mientras que concentración de 50 mM NaCl incrementó la biomasa seca en 19%, ambas respecto al control (Cuadro 3).
En este sentido Camejo y Torres (2000) observaron un incremento de la biomasa seca en plantas de tomate en condiciones salinas. Las plantas al estar expuestas en un medio salino activan su mecanismo de defensa y elevan los niveles de enzimas, prolinas, aminoácidos y antioxidantes. La presencia de Si, independientemente de la concentración, incrementó la tasa de germinación y la longitud radicular (Cuadro 3).
Cuadro 3 Efectos principales de NaCl y del Si en la germinación de tomate Floradade (Solanum lycopersicum var. Floradade).
| Dosis | Variables evaluadas | |||||||
| % GP | GE | GRI | MGT | IG | VG | LR | BS | |
| NaCl | ||||||||
| 0 mM | 72.333 | 13.404 | 6.5 a | 0.132 | 0.09 | 0.283 | 4.034 b | 1.44 b |
| 25 mM | 72.667 | 13.658 | 6.211 ab | 0.133 | 0.091 | 0.284 | 4.814 a | 1.5 b |
| 50 mM | 68 | 12.925 | 5.585 b | 0.128 | 0.085 | 0.266 | 3.948 b | 1.71 a |
| ±EE | 2.599 | 0.553 | 0.198 | 0.003 | 0.003 | 0.01 | 0.222 | 0 |
| Lineal | ns | ns | 0.003 | ns | ns | ns | ns | 0.003 |
| Cuadrática | ns | ns | ns | ns | ns | ns | 0.005 | ns |
| Si | ||||||||
| 0 mM | 70.333 | 13.523 | 5.458 b | 0.13 | 0.088 | 0.275 | 3.675 b | 1.57 |
| 1 mM | 71.333 | 13.333 | 6.235 a | 0.131 | 0.089 | 0.279 | 4.479 a | 1.58 |
| 2 mM | 71.333 | 13.132 | 6.603 a | 0.132 | 0.089 | 0.279 | 4.642 a | 1.49 |
| ±EE | 2.599 | 0.553 | 0.198 | 0.003 | 0.003 | 0.01 | 0.222 | 0 |
| Lineal | ns | ns | <0.001 | ns | ns | ns | 0.005 | ns |
| Cuadrática | ns | ns | ns | ns | ns | ns | ns | ns |
Literales diferentes entre las medias indican diferencia significativa. Porcentaje de germinación (%GP); energía de germinación (GE); tasa de germinación (GRI); tiempo medio de germinación (MGT); índice de germinación (IG); velocidad de germinación (VG); longitud de raíz (LR); biomasa seca (BS); NS= no significativo; ±EE= error estándar.
La interacción NaCl-Si, mostró que la presencia de Si, incrementó la longitud de raíz y biomasa fresca (Figura 2). Khan et al. (2020) evaluaron la termo tolerancia inducida por Si en plantas de tomate con 1 y 2 mM. Los resultados mostraron que el Si mejoró significativamente el crecimiento de las plantas, biomasa fresca y longitud de la raíz, en comparación para el tratamiento control. De la misma manera, Mushinskiy et al. (2018) encontraron que la aplicación de Si, incrementó la longitud de la raíz de plántulas de tomate en 27.8% en comparación para el tratamiento control.
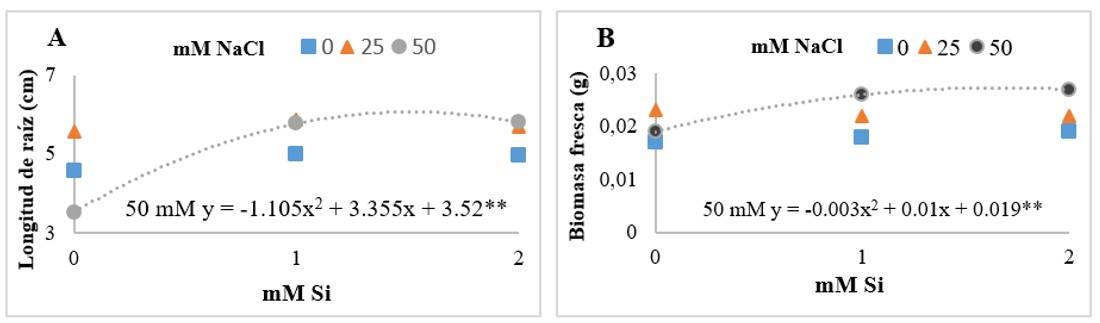
Figura 2. Efecto de la interacción de cloruro de sodio (NaCl) y silicio (Si) en la longitud de raíz (A); y la biomasa fresca (B) de plántulas de tomate Floradade después de los 14 días del establecimiento del experimento. Los asteriscos (*) indican los niveles de significancia entre los tratamientos 0.01** - 0.05*.
En el presente estudio se observó que la concentración de 50 mM de NaCl redujo de manera significativa la tasa de germinación en 31% en comparación para el tratamiento control. Contrario a lo anterior, se observó que las dosis de 25 y 50 mM estimularon la longitud de la raíz en 36.02 y 27.57% en comparación para el tratamiento control. Además, la longitud del tallo también se vio incrementada en 40 y 26% en dichas concentraciones de NaCl (Cuadro 4).
Cuadro 4 Efectos principales de NaCl y del Si en la germinación de tomate saladette Río Grande (Solanum lycopersicum var. Río Grande).
| Dosis | Variables evaluadas | ||||||||
| % GP | GE | GRI | MGT | IG | VG | LR | LT | BS | |
| NaCl | |||||||||
| 0 mM | 71 | 13.253 | 6.169 a | 0.132 | 0.089 | 0.277 | 5.744 b | 5.351 c | 1.85 b |
| 25 mM | 67 | 12.879 | 5.13 ab | 0.128 | 0.084 | 0.262 | 7.813 a | 7.494 a | 2 ab |
| 50 mM | 59.333 | 11.579 | 4.253 b | 0.116 | 0.074 | 0.232 | 7.328 a | 6.766 b | 2.13 a |
| ±EE | 3.752 | 0.718 | 0.341 | 0.005 | 0.005 | 0.015 | 0.299 | 0.188 | 0 |
| Lineal | ns | ns | < 0.001 | ns | ns | ns | < 0.001 | < 0.001 | 0.006 |
| Cuadrática | ns | ns | ns | ns | ns | ns | 0.002 | < 0.001 | ns |
| Si | |||||||||
| 0 mM | 64.333 | 12.558 | 4.61 a | 0.124 | 0.08 | 0.251 | 5.942 b | 5.873 b | 2.11 a |
| 1 mM | 66.333 | 12.553 | 5.442 a | 0.125 | 0.083 | 0.259 | 7.091 a | 6.68 a | 1.86 b |
| 2 mM | 66.667 | 12.601 | 5.501 a | 0.126 | 0.083 | 0.26 | 7.853 a | 7.058 a | 2.02 ab |
| ±EE | 3.752 | 0.718 | 0.341 | 0.005 | 0.005 | 0.015 | 0.299 | 0.188 | 0 |
| Lineal | ns | ns | ns | ns | ns | ns | < 0.001 | < 0.001 | ns |
| Cuadrática | ns | ns | ns | ns | ns | ns | ns | ns | 0.016 |
Literales diferentes entre las medias indican diferencia significativa. Porcentaje de germinación (%GP); energía de germinación (GE); tasa de germinación (GRI); tiempo medio de germinación (MGT); índice de germinación (IG); velocidad de germinación (VG); longitud de raíz (LR); longitud de tallo (LT); biomasa seca (BS); ns= no significativo; ±EE= error estándar.
Nuestros resultados se corroboran con los obtenidos por Loudari et al. (2020) quienes observaron el efecto por NaCl en tomate, el cual incremento la longitud y diámetro de la raíz. Por otra parte, Batista et al. (2017) evaluaron el efecto del NaCl en la germinación de tres variedades de albahaca (Ocimum basilicum L.). Donde observaron una disminución en el porcentaje y tasa de germinación. De la misma manera, González et al. (2020) investigaron el efecto del NaCl en la germinación de semillas de tomate cv ‘Río Grande. Observaron que la tasa de germinación disminuía de manera significativa en comparación para el tratamiento control.
Las concentraciones de 1 y 2 mM de Si impactaron de manera significativa al incrementar la longitud de raíz y de tallo, ambas concentraciones fueron superiores al tratamiento control (Cuadro 4). Se ha demostrado que el Si a pesar de no ser un elemento esencial para las plantas, favorece en el crecimiento y desarrollo de algunas especies. En este sentido Chourasiya et al. (2021) mencionan que el efecto del Si en la germinación y crecimiento inicial de trigo (Triticum), presenta un efecto positivo en las variables evaluadas.
El Si incremento de manera significativa la biomasa fresca (Figura 3). Particularmente en la concentración 50 mM NaCl, la presencia de Si generó un efecto lineal positivo. Ahmad et al. (2019) evaluaron el factor interacción de Si + NaCl en plántulas de frijol, las cuales mejoró los efectos adversos del NaCl sobre los parámetros de crecimiento, la biomasa fresca y seca, síntesis de pigmentos y contenido relativo de agua.
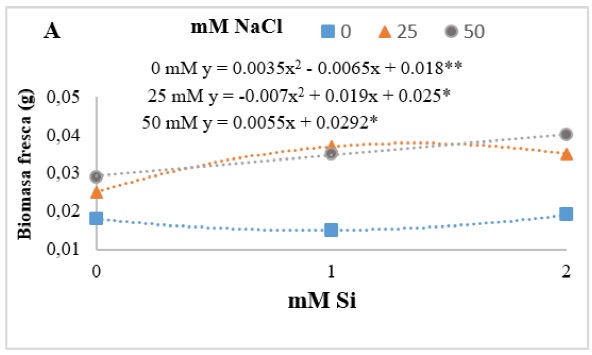
Figura 3. Efecto de la interacción de cloruro de sodio (NaCl) y silicio (Si) en la biomasa fresca (A) de plántulas de tomate Río Grande después de los 14 días del establecimiento del experimento. Los asteriscos (*) indican los niveles de significancia entre los tratamientos 0.01** - 0.05*.
Además, Torabi et al. (2012) informaron que el Si impacto de manera positiva en el crecimiento de las plántulas de borraja (Borago officinalis), incrementando así la biomasa fresca y seca de las mismas. Por lo consecuente, el Si una vez dentro de la planta se localiza en diferentes estructuras, se difunde a través de los tejidos y forma una capa continua entre la cutícula y la epidermis compuesta por dos subcapas, una como gel de Si y otra en complejo silico-celulosa, la cual aísla y defiende a la planta de los factores adversos del medio ambiente.
En este sentido Emamverdian et al. (2018), menciona que el Si tiene la capacidad de activar los genes Lsi1, Lsi2 y Lsi6 en las plantas, el gen Lsi2 se considera un transportador de aniones y tiene la capacidad de expresarse en la endodermis de la raíz; mientras que, Lsi1 y Lsi6 son genes que pertenecen a la familia de las acuaporinas y su función es transportar el silicio a los tejidos de los brotes y raíces. Además, al expresarse los genes se activan múltiples enzimas tales como: catalasas, peroxidasas, superóxido dismutasas, ascorbato peroxidasas y glutatión reductasas las cuales contribuyen con la planta a aliviar el estrés (biótico o abiótico).
Conclusiones
Los cultivares de tomate mostraron efectos diferentes respecto a la salinidad. El cultivar cherry mostró disminuciones en sus variables germinativas al incrementarse la salinidad, lo que indica que los cultivares Floradade (bola) y Río Grande (saladette) presentaron mayor tolerancia a la salinidad. El silicio (Si) tuvo un efecto protector en la interacción (NaCl + Si), mostrando efectos positivos al incrementar las variables evaluadas. Por lo anterior, el Si pudiera considerarse como un agente atenuador de la salinidad.

