










Services on Demand
Journal
Article
 text in
text in  English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista mexicana de ciencias agrícolas
Print version ISSN 2007-0934
Rev. Mex. Cienc. Agríc vol.13 n.6 Texcoco Aug./Sep. 2022 Epub Feb 24, 2023
https://doi.org/10.29312/remexca.v13i6.3311
Articles
Effect of mineral N and N2 on nitrogen nutrition in soybean plant
1Campo Experimental Las Huastecas-INIFAP. Carretera Tampico-Mante km 55, Villa Cuauhtémoc, Tamaulipas, México. CP. 89610. (zuniga.lamberto@inifap.gob.mx).
2Tecnológico Nacional de México-Campus Instituto Tecnológico de Torreón. Antigua carretera Torreón-San Pedro km 7.5, Torreón, Coahuila, México. (reguillen@outlook.com).
3 División Académica de Ciencias Agropecuarias-Universidad Juárez Autónoma de Tabasco. Carretera Villahermosa-Teapa, Ra. km 25, La Huasteca, 2da. Sección. Centro, Tabasco, Mexico. (efrain.delacruz@ujat.mx).
4 Universidad Autónoma Agraria Antonio Narro-Unidad Laguna. Periférico y Carretera a Santa Fe s/n, Torreón, Coahuila, Mexico. (juan-manuelnava@hotmail.com).
The biological nitrogen fixation is a sustainable option to nitrogen fertilization. The objective of this study was to evaluate the effect of mineral nitrogen and N2 from the biological nitrogen fixation on nitrogen nutrition in soybean plants, six treatments resulting from combining three levels of mineral N: (0.0, 3.5 and 7 meq L-1 of NO3 -) and inoculation with the CP-2 strain of Bradhyrizobium japonicum (with and without inoculation) were evaluated, each treatment was established in triplicate, in three-liter plastic pots and agrolite as substrate, transplanting six soybean seedlings per pot. Dry matter (DM) in plant organs, number and dry weight of nodules, fixation of N2, soluble N (N-amino acid and N-ureido) and total N per organ were evaluated. The plants that received mineral N transported between 86 and 95% of the total soluble N as N amino acid, while in inoculated plants, the supply of mineral N negatively affected the number of nodules (29%) and the weight of nodules (64%), decreasing 55% the fixation of N2, in these plants between 50 and 70% of the soluble N transported was as N ureido. At the R6 stage, it was estimated that 70% of the total N accumulated in the DM came from the fixed N2, favoring the DM of leaves, stems, petioles and mainly the DM in pods. Therefore, in this study it is demonstrated that plants inoculated and supplied with mineral N obtain better effects for DM and total N.
Keywords: inoculation; N-amino acid; N-ureido.
La fijación biológica del nitrógeno, es una opción sustentable a la fertilización nitogenada. Con el objetivo de este estudio fue evaluar el efecto del nitrógeno mineral y del N2 proveniente de la fijación biológica del nitrógeno sobre la nutrición nitrogenada en plantas de soya, fueron evaluados seis tratamientos resultantes de combinar tres niveles de N mineral:(0.0, 3.5 y 7 meq L-1 de NO3 -) e inoculación con la cepa CP-2 de Bradhyrizobium japonicum (con y sin inoculación), cada tratamiento se estableció por triplicado, en macetas de plástico de tres litros de capacidad y agrolita como sustrato transplantándose seis plántulas de soya por maceta. Se evaluó la materia seca (MS) en órganos de la planta, número y peso seco de nódulos, fijación de N2, N soluble (N-aminoácido y N-ureido) y N total por órgano. Las plantas que recibieron N mineral transportaron entre 86 y 95% del N soluble total como N aminoacido, mientras que en plantas inoculadas el aporte de N mineral afectó negativamente la cantidad de nódulos (29%) y el peso de nódulos (64%), disminuyendo 55% la fijación de N2, en estas plantas entre 50 y 70% del N soluble transportado fue como N ureido. A la etapa de R6, se estimó que 70% del N total acumulado en la MS provino del N2 fijado, favoreciéndose la MS de hojas, tallos, peciolos y principalmente la MS en vainas. Por lo tanto, en este estudio demuestra que las plantas inoculadas y abastecidas con N mineral obtienen mejores efectos para la MS y el N total.
Palabras clave: inoculación; N-aminoácido; N-ureido
Introduction
In agriculture, many plants of interest are legumes due to their high protein content (~40%), having a great nutritional value, however, it requires high nitrogen content, known to be a limiting factor, at the same time, a natural alternative implemented by plants to meet this need is through the BNF (Meena et al., 2017). However, some authors favor the implementation of inorganic fertilizers, but their excessive use and a bad practice leads to the alteration of ecosystems (Soumare et al., 2020). On the other hand, nitrogen fertilizers and BNF carried out by some microorganisms are the sources that allow nitrogen (N) to be incorporated into the soil; both processes use the N contained in the atmosphere.
The soybean plant, one of the most important legumes, establishes symbiosis with bacteria of the genus Bradhyrizobium japonicum and satisfies its needs for N through two assimilation pathways, one pathway is the BNF that is carried out in the bacteroids of the nodules formed in the root, where between 50 and 60% of the total N required by the plant is fixed, and the second pathway is the absorption by the root of the N-mineral (NH4 + and NO3 -) present in the soil naturally or applied with chemical fertilizers (Ortez et al., 2017).
Worldwide, an estimated of ~2.5 × 1011 kg NH3 is fixed annually with the BNF (Meena et al., 2017). Currently, the potential for symbiotic N fixation by legumes is underexploited because it only involves 13% of fertilization on farmland (Anglade et al., 2015). Since the concentration of N in soil organic matter is limiting and rapidly reduces with a few cultivation cycles and that the efficiency of nitrogen fertilizers turns out to be less than 50%, from an economic and environmental perspective, BNF is currently the most sustainable way to incorporate N into the soybean plant (Saturno et al., 2017).
Traditionally, the soybean plant has been used in controlled laboratory and field studies to understand the processes of absorption and metabolism of N-mineral and N2 from BNF, as well as their distribution in plant organs (Ohyama et al., 2017; Senthilkumar et al., 2021). Studies with 15N as NO3 -, NH4 + or N2, showed that the two sources of N are reduced to ammonium (NH4 +) at the root (Yoneyama et al., 2020) and used in the synthesis of amino acids and proteins (Ertani et al., 2013); however, the source of N absorbed by the plant controls its distribution in the organs of the plant (Balta-Crisólogo et al., 2015).
In the soybean plant supplied with mineral N, the N is transported as N-amino acid and as N-ureido (allantoin and allantoic acid) the N from the BNF (Hartmann et al., 2012). Both forms of N supply foliage organs and reproductive structures (Kipp et al., 2020). N-amino acid supplies the vegetative apparatus and more specifically the stems (Yan et al., 2016), while N-ureido is used in developing organs and during the reproductive period are the pods, structures in which 52% of N from the BNF was determined (Gregg et al., 2015; Chen et al., 2018). However, the use of mineral N could increase yield in the soybean plant under very specific conditions, but this practice is rarely economically viable (Saturno et al., 2017).
Studies on BNF in soybeans have also focused on understanding the complex molecular interactions that control a) the selectivity existing between the plant and Bradhyrizobium japonicum; b) the formation of the nodules (Bala and Giller, 2006); c) the factors that affect the efficiency of fixation, Freixas et al. (2011); d) the factors that influence the senescence of the nodules (Fernández-Luqueño and Espinosa-Victoria, 2008); and e) the methods to quantify the N from the BNF (Ortez et al., 2017).
In soybean-producing countries, genetic improvement programs have taken advantage of advances in the BNF, identifying lines highly efficient in the BNF (Cerezini et al., 2020), increasing yield and decreasing the use of nitrogen fertilizers (Fontanetto et al., 2011). Studies conducted on genotypes commercially released in different decades (1980-2010) in the US and Argentina confirmed greater yield potential when receiving nitrogen fertilization, the genotypes released in 2010 had a higher yield between 15 and 34% than that recorded in materials released before 2010 in the US and Argentina, respectively.
Despite the aforementioned advances, the interaction between soybean genotypes and the nitrogen fertilizer response is a topic that requires further study, so the objective of the present study was to contribute to the knowledge of nitrogen nutrition and the dry matter production of the soybean plant.
Materials and methods
The research was carried out at the College of Postgraduates, Montecillo Campus (19° 28’ 4.26” north latitude, 98° 53’ 42.18” west longitude and an altitude of 2 250 m) in Texcoco, State of Mexico, under conditions of zenith-type greenhouse of metal structure covered with milky white plastic (gauge 720).
Seeds of soybean cv Santa Rosa were sown in polypropylene trays of 200 cavities, until the seedlings showed the first pair of true leaves. Subsequently, six seedlings were transplanted in a previously disinfected 5 L plastic pot (experimental unit), agrolite was used as a substrate in the pots. The treatments consisted of developing the soybean plant under six conditions, resulting from combining: a) three levels of mineral nitrogen 0, 3.5 and 7 meq L-1 of N, using reactive grade NH4 NO3 as a source of nitrogen; and b) two levels of inoculation (with and without inoculation); each treatment was applied in triplicate.
The experimental design used was completely randomized. On the third day after transplantation, the plants of the corresponding treatments were inoculated, applying at the base of the stem of each plant 1 ml of culture that contained 109 cells of Bradyrizobium japonicum bacteria from the CP2 strain of the microbiology collection of the CP. Irrigations, from germination to inoculation, were provided daily with distilled water and after inoculation with nitrogen-free nutrient solution (Tirado and Alcántar, 1989).
The three nitrogen treatments applied to the soybean plants were prepared with the nutrient solution, starting their application 15 days after inoculation, this nutrient solution was renewed every week. (Table 1)
Table 1 Composition of the nutrient solution without nitrogen used during the development of the study.
| Salt | (meq L-1) |
| KH2PO4 | 2.4 |
| MgSO4 | 1.2 |
| CaCl2 | 1 |
| K2SO4 | 1 |
| (ppm) | |
| H3BO3 | 2 |
| MnSO4 4H2O | 1.8 |
| ZnSO4 | 0.2 |
| CuSO4 | 0.8 |
| NaMoO4 2H2O | 0.04 |
| Fe-EDTA | 16 |
During the development of the plant, four destructive samplings were carried out; vegetative stage (V7), flowering (R2), pod formation (R4) and maximum grain filling (R6), stages recorded at 28, 54, 79 and 103 days after sowing (DAS), respectively. The six plants per pot were taken in each evaluation, recording the wet (WW) and dry (DM) weight of leaves, stems (when present) and roots. In this last organ, in the first three phenological stages, the biological nitrogen fixation (BNF) was evaluated by the nitrogenase (N’asa) activity with the acetylene reduction method (ARA), the number and dry weight of nodules per plant.
At each sampling stage, 0.22 g of fresh matter from petioles, 0.5 g for stems and leaves and 1 g in roots were taken, each sample was wrapped in aluminum foil and preserved in liquid nitrogen (-70 °C) until their use in the determination of N-soluble; the samples used for the determination of total nitrogen were dried in a forced-air oven for 72 h at 70 C. Samples of fresh plant material were ground with a mortar in 10 ml of the potassium phosphate buffer solution 0.1 M, pH 7.4, filtered through gauze to separate the thick residues. From this plant extract, the N-amino acid and the N-ureido were determined (Tirado and Alcántar, 1989). In dry plant material, the total N was determined by the Microkjeldahl method. The data obtained from the response variables were analyzed by analysis of variance and mean separation test using the Tukey test (p≤ 0.05).
Results and discussion
Number and activity of nodules
The number, dry weight of nodules and ARA values in the soybean plant, from the V7 stage to the R4 stage, were negatively affected by the mineral N supplied in the nutrient solution (Table 2). The supply of 3.5 and 7 meq L-1 of N in the nutrient solution caused a decrease of 9 and 25 nodules in the root of the soybean plant, respectively. Similarly, the dry weight of nodules per plant was mostly affected when the plant received 7 meq of N L-1 (Table 2).
Table 2 Average values of the number, dry weight and activity of nodules in the fixation of N2 (ARA) recorded in three stages of evaluation due to the effect of the mineral N supplied in the nutrient solution.
| Treatment | Nodules plant-1 | ARA (nmol of C2H4 h-1 p-1) | |
| Number | Dry weight (mg) | ||
| 0 meq L-1 of N + Inoc | 85 (100)* | 117.5 (100) | 1.02 (100) |
| 3.5 meq L-1 of N + Inoc | 76 (89) | 80.5 (69) | 0.89 (87) |
| 7 meq L-1 of N + Inoc | 60 (71) | 42.1 (36) | 0.46 (45) |
*= value in parentheses is % relative to nitrogen-free treatment.
N’asa activity showed variation with plant development and was also negatively affected by the application of mineral N. Due to the effect of the supply of N in the nutrient solution, the mean value of the N’asa activity estimated in three stages of the development of the plant (V7, R2 and R4) decreased by 13 and 55% when they received 3.5 and 7 meq L-1 of N in the nutrient solution, respectively.
The causes that originate the decrease in the N’asa activity could be due, according to Stal et al. (2017), to the effect of regulating compounds such as oxygen or temperatures, while Xia et al. (2017) mentioned that the application of NO3 - reduced the sending of carbohydrates to the nodules and they attribute it to the formation of NO2 -, which is toxic to nitrogenase and interferes with oxygen regulation and therefore in the fixation of N2.
Soluble nitrogen (N amino acid and N ureido)
As the cycle progressed, the growth of vegetative and reproductive organs caused in the soybean plant an increase in the demand for N of the foliage, with significant increases (p≤ 0.05) in the concentration of soluble N transported from the V7 stage to the R4 stage. The exception of this trend was recorded in inoculated plants that did not receive mineral N and in non-inoculated plants developed with 7.5 meq L-1 of mineral N, both plants showed at R2 the maximum value in the concentration of soluble N transported and it decreased at the R4 stage (Table 3).
Particularly at the R2 stage, the plants that received mineral N in the nutrient solution (3.5 and 7 meq L-1 of N) had the highest concentration of soluble N transported, whose values decreased 20 and 7% when receiving inoculation; a decrease that could be caused by the competition between the development of nodules at the root and the growth of the foliage organs (Table 3). At the R4 stage, once the nodules in these same plants were established and developed, the activity of the nodules and the mineral N absorbed by the root from the nutrient solution caused an increase of 7 and 43% in the values in the concentration of soluble N transported in relation to the concentration of soluble N transported in non-inoculated plants supplied with 3.5 and 7 meq L-1 of mineral N in the nutrient solution, respectively (Table 3).
Table 3 Concentration of soluble N (μg of N gmf-1) transported in the stem at different stages of development of the soybean plant.
| Treatment | Vegetative (V7) | Flowering (R2) | Reproductive (R4) |
| (μg of N g mf-1) | |||
| 0 meq of N | 77.5 c | 104.5 f | 128.8 e |
| 3.5 meq of N | 168.1 a | 295.6 d | 318.4 c |
| 7 meq of N | 111.3 b | 384.7 a | 284.3 d |
| 0 meq L-1 of N + Inoc | 83.7 c | 329.3 c | 296.5 d |
| 3.5 meq L-1 of N + Inoc | 110.6 b | 235.2 e | 341.5 b |
| 7 meq L-1 of N + Inoc | 120.2 b | 359 b | 406.9 a |
| HSD | 17.2 | 9.6 | 15.8 |
Values with different letters in the same column are different (Tukey p≤ 0.05).
The total soluble N transported was formed by N-amino acid and N-ureido, their proportion in each evaluation depended on the stage of development and on the source of assimilated N (mineral N or N2). In the initial stages of plant development (V7), N-amino acid predominated as the soluble N form regardless of the N form the plant received, Figure 1A. When the plants received exclusively mineral N, in the stages of R2 and R4, they presented between 86 and 95% of the soluble N in the form of N-amino acid, respectively (Figure 1B and 1C), values similar to those reported by Santachiara et al. (2017); Junior et al. (2020), who mentioned that, in soybean plants supplied with mineral N, between 77% and 90% of soluble N is transported as N amino acid.
The same Figures 1B and 2C show that when the soybean plant, in addition to the mineral N, also received inoculation, it had a decrease in the value of N amino acid transported with respect to the plants supplied only with mineral N. At the R2 stage the values of N amino acid fluctuated between 66 and 82%, Figure 1 B; while at the R4 stage, the mean value of the transported N amino acid decreased up to 35%, the decrease in the N amino acid was caused by the activity of N2 fixation of the nodules that stimulated the production of N ureido, as mentioned by Lopes (2015); McCoy et al. (2018), results that agree with those reported by Tamagno et al. (2018), who mentioned that, in plants developed under these conditions, 60% to 81% of soluble N is transported for rhizodeposition pathways; that is, decomposition and deterioration of root nodules and cells (Fustec et al., 2010; Collier et al., 2012).
In the inoculated plants, the N-ureido transported showed the maximum values in the R4 stage, an inverse situation occurred in the values of ARA determined during this stage, the above can be attributed to the fact that both methods must be calibrated, as performed by Grageda et al. (2003), using non-destructive techniques, they reported that after R5 the soybean plant fixed 78% of N.
It is important to note that between the stages of R2 and R4, the plants supplied with 3.5 meq of N presented 79% of the soluble N transported as N ureido, observing that this form of N was reduced to 50% when the plant received 7 meq of N in the nutrient solution. Decreases in the amount of N-ureido due to the effect of external application of N had been reported by Ono et al. (2021), these researchers mentioned that, in nodulated plants, the N-ureido is relatively low, because it is converted to urea, actively occurring in the underground parts, especially in the roots, in their study, nitrate represented about 50% of N applying 5 mM of NaNO3.
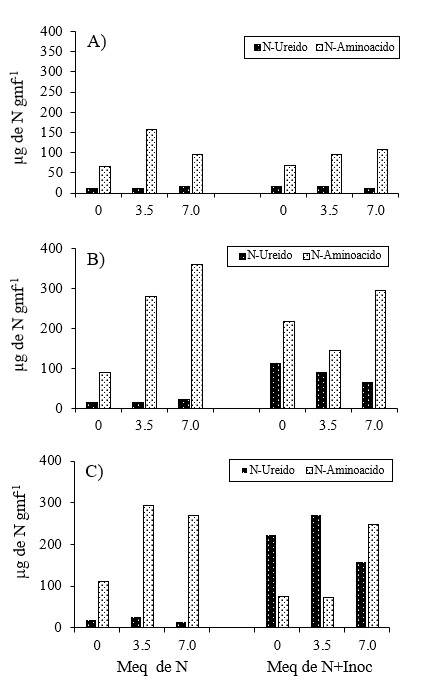
Accumulation and distribution of N
The mineral N absorbed by the root or the N2 fixed in the nodules, once assimilated and transported as soluble N, accumulated as total N in the DM of the organs of greatest demand at each stage of plant development. At the R2 stage, the plants that received 7 meq of N in the nutrient solution had the greatest accumulation of total N in their DM, this form of N favored the total N accumulated in the DM of the stems and petioles, whose differences due to the form of N received in the same organ were significant (p≤ 0.05).
The lowest value of total N occurred in the DM of plants that did not receive mineral N and inoculation, and represented 10% with respect to plants that accumulated the highest amount of total N. At this R2 stage, regardless of the source of N that the plant received, the total N recorded on average represented 72, 18 and 10% of the total N of the DM of leaves, stems and petioles, respectively.
The largest increase in total N in the DM of all plants was recorded between the stage of R2 and R6. When these were inoculated and received mineral N accumulated the largest amount of total N and on average it was 2.3 times more than the total N value recorded in the DM of plants that only received the mineral N, these differences were significant (p≤ 0.05). Particularly, in the soybean plant that received 3.5 meq of N L-1, it had an accumulation of 158 mg of N plant-1 in the DM, amount of total N that, considering a density of 437 plant ha-1, allows estimating a total extraction of 72.2 kg of N ha-1; this extraction of total N, due to the effect of the mineral N, represented 30% of the total N determined in the DM of plants supplied with the same dose of mineral N and which were also inoculated (Table 4). That is, in this plant, 160.5 kg of N ha-1 determined in the DM was obtained due to the effect of the activity of the nodules, deducing 70% of the N accumulated in the DM at the R6 stage came from the fixation of N2; the above shows that both pathways of assimilation of N act in the plant in a complementary way, as previously mentioned by Lu et al. (2021).
Table 4 Amount of N accumulated (mg plant-1) in the dry matter of the organs of the soybean plant during the reproductive stage (R6).
| Treatment | Total | Leaf | Stem | Petiole | Pod |
| (mg of N plant-1) | |||||
| 0 meq of N | 41.9 e | 7.1 e | 2.9 d | 1 d | 31 f |
| 3.5 meq of N | 158 c | 36.7 c | 4.6 c | 2.2 d | 114.5 d |
| 7 meq of N | 274.1 b | 93.6 b | 12.9 b | 6.2 c | 161.4 c |
| 0 meq of N + Inoc | 98.4 d | 23.4 d | 3.6 cd | 2.2 d | 69.2 e |
| 3.5 meq of N + Inoc | 509.2 a | 94.6 b | 21 a | 13.9 a | 379.7 a |
| 7 meq of N + Inoc | 487.1 a | 128.9 a | 22.2 a | 10.3 b | 325.7 b |
| HSD | 26 | 4 | 1.7 | 0.9 | 7.1 |
Values with different letters in the same column are different (Tukey p≤ 0.05).
During this reproductive stage, the high demand for N of the pods caused that, in the plants supplied with mineral N exclusively and in the inoculated ones that did not receive mineral N, part of the total N determined at flowering in the organs of the foliage was mobilized towards the pods, in the inoculated plants that did not receive mineral N, 41.6% of the N determined in the pods came from the N stored in the foliage at R2, while in plants supplied exclusively with the highest dose of N, this percentage was 9.9% and 4.4% in those that received the intermediate dose.
Accumulation and distribution of dry matter (DM) per plant
The lowest amount of DM was recorded in the plant that did not receive mineral N and neither inoculation, at the R2 stage, in this plant the accumulated DM represented 46% of the DM produced in the plant that received only inoculation. At this stage, the plant that received the highest dose of N and inoculation was surpassed 0.5 g of DM plant-1 by the registered DM that received the highest dose of N (Table 5), a situation that can be associated with the competition that generated the formation of nodules in the root.
Table 5 Dry matter production in the soybean plant in two phenological stages.
| Treatment | Flowering (R2) | Maximum grain filling (R6) | Increase (R2 to R6) |
| (g plant-1) | |||
| 0 meq L-1 of N | 0.6 d | 1.4 f | 0.8 |
| 3.5 meq L-1 of N | 2 b | 6.2 d | 4.2 |
| 7 meq L-1 of N | 2.8 a | 8.1 c | 5.3 |
| 0 meq L-1 of N + Inoc | 1.5 c | 3.3 e | 1.8 |
| 3.5 meq L-1 of N + Inoc | 2 b | 11.8 b | 9.8 |
| 7 meq L-1 of N + Inoc | 2.3 b | 13.3 a | 11 |
| HSD | 0.4 | 0.8 | |
Values with different letters in the same column are different (Tukey p≤ 0.05).
The highest production of DM in the plant occurred between the stage of R2 and R6, regardless of the dose and form of N the plant received for nutrition. Plants supplied with N-mineral and consistently inoculated produced more DM, the mean value of DM recorded at R6 in these plants exceeded 1.7 times the DM of plants that only received N-mineral (3 and 7 meq L-1) and 9.3 times the DM of plants developed without mineral N and without inoculation (Table 5).
Regardless of the nitrogen nutrition that the plant received during its development, of the total value of the DM produced at the R6 stage, on average 59% was recorded in pods, 26% in leaves and 15% in stems + petioles (Figure 1). The inoculated plants that received mineral N produced more amount of DM in their organs, in pods the value was 2.2 times significantly higher (p≤ 0.05) than the value of DM recorded in the pods of plants that only received mineral N; in stems plus petioles and leaves, this value was 1.4 and 1.3, respectively. These results are consistent with those recorded by Tirado et al. (1990), demonstrating that the two pathways of assimilation of N in the soybean plant are necessary to obtain high yields of DM.
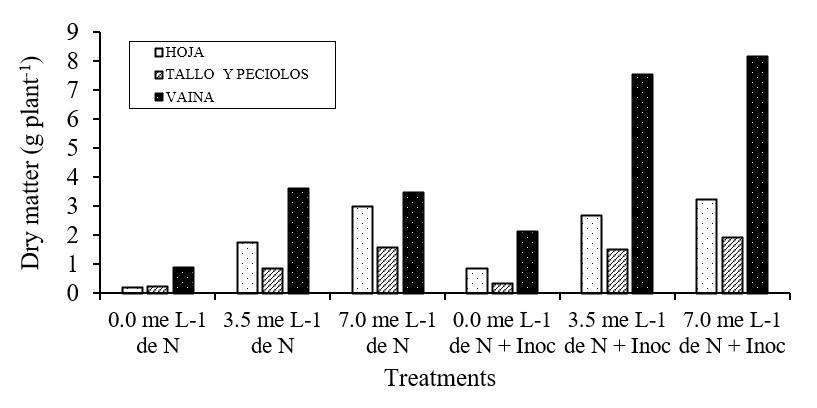
Figure 2 Dry matter (g plant-1) accumulated in the organs of the soybean plant of the Santa Rosa variety, during the grain filling stage (R6).
These results indicate that, in nodulated soybean plants, the accumulation of DM in the pods is favored by the activity of the nodules and that the foliage is an important source of N, being able to conclude that the fixed N (N-ureido) is destined primarily to the supply of grains, as mentioned by (Anglade et al., 2015; Zhou et al., 2016; Ono et al., 2021).
Conclusions
The application of mineral N affected the number of nodules per plant, the dry weight of nodules and the nitrogenase activity. The greatest accumulation of dry matter and total nitrogen was in plants inoculated and supplied with mineral N. In nodulated plants, between 39 and 79% of the soluble N transported in the stem after flowering was recorded in the form of N-ureido. Of the total N determined in the pods, between 50 and 70% was assimilated via N2 fixation.
Literatura citada
Anglade, J.; Billen, G. and Garnier, J. 2015. Relationships for estimating N2 fixation in legumes: Incidence for N balance of legume-based cropping systems in europe. Ecosphere. 6(3):1-24. https://doi.org/10.1890/ES14-00353.1. [ Links ]
Bala, A. and Giller, K. E.. 2006. Relationships between rhizobial diversity and host legume nodulation and nitrogen fixation in tropical ecosystems. Nutr. Cycling Agroecosyst. 76(2):319-330. [ Links ]
Balta, C. R. A.; Rodríguez, D. Á. M.; Guerrero, A. R.; Cachique, D.; Edín, A. P.; Arévalo, L. L. y Oscar, L. O. L. I. 2015. Absorción y concentración de nitrógeno, fósforo y potasio en sacha inchi (Plukenetia volubilis L.) en suelos ácidos, San Martín, Perú. Folia Amazónica. 24(2):23-30 [ Links ]
Cerezini, P.; Kuwano, B. H.; Grunvald, A. K.; Hungria, M. and Nogueira, M. A. 2020. Soybean tolerance to drought depends on the associated Bradyrhizobium strain. Brazilian Journal of Microbiology. 51(4):1977-1986. [ Links ]
Chen, C.; Wang, J.; Liu, D.; Yang, C.; Liu, Y.; Ruoff, R. S. and Lei, W. 2018. Functionalized boron nitride membranes with ultrafast solvent transport performance for molecular separation. Nature communications. 9(1):1-8. [ Links ]
Collier, R. and Tegeder, M. 2012. Soybean ureide transporters play a critical role in nodule development, function and nitrogen export. The Plant Journal. 72(3):355-367. [ Links ]
Ertani, A.; Pizzeghello, D.; Altissimo, A. and Nardi, S. 2013. Use of meat hydrolyzate derived from tanning residues as plant biostimulant for hydroponically grown maize. Journal of Plant Nutrition and Soil Science. 176(2):287-295. [ Links ]
Fernández, L. L. y Espinosa, V. D. 2008. Bioquímica, Fisiología, Morfología y senescencia nodular; Una revisión crítica. Terra Lationamericana. 134(26):133-144 [ Links ]
Fontanetto, H.; Keller, O.; Sillón, M.; Albrecht, J.; Giailevra, D.; Negro, C. y Belotti, L. 2011. Manejo de la fertilización de la soja en regiones templadas. Informacion Tecnica Cultivos De Verano. Campaña 201.1INTA-Estación Experimental Agropecuaria Rafaela. Publicación Miscelánea núm. 121. [ Links ]
Freixas , C. J. A.; Torres de la N. W.; Escobar, M. I. M. R. y Napoles, G. M. 2011. Niveles de Ureidos en plantas de soya con diferentes iniculantes y sometidas a Dedicit Hidrico. Cultivos tropicales. 3(2):35-43. [ Links ]
Fustec, J.; Lesuffleur, F.; Mahieu, S. and Cliquet, J. B. 2010. Nitrogen rhizodeposition of legumes. A review. Agronomy for sustainable development. 30(1):57-66. [ Links ]
Grageda, C. O. A.; Mora, M.; Castellanos, R. J. Z.; Follet, R. F. and Peña, C. J. J. 2003. Fertilizer nitrogen recovery under different tillage treatments and cropping sequences in a vertisol in central México. IAEA-TECDOC. Viena. 1354(1):39-55. [ Links ]
Gregg, G. L.; Orlowski, J. M. and Lee, C. D. 2015. Input‐based stress management fails to increase soybean yield in Kentucky. Crop, Forage & Turfgrass Management. 1(1):1-7. [ Links ]
Hartmann, T. 2012. VI. Metabolism of Organic N-Compounds. Progress in Botany/Fortschritteder Botanik: Morphology physiology Genetics Taxonomy Geobotany /Morphologie Physiologie Genetik Systematik Geobotanik. 44:154-164. [ Links ]
Junior, C. P.; Favarin, J. L.; Lago, B. C.; Almeida, R. E. M.; Oliveira, S. M.; Trivelin, P. C. O. and Gilabel, A. P. 2020. Nitrogen Fertilizer Recovery and Partitioning Related to Soybean Yield. Journal of Soil Science and Plant Nutrition. 20(4):2566-2578. [ Links ]
Kipp, M. A.; Stüeken, E. E.; Gehringer, M. M.; Sterelny, K.; Scott, J. K.; Forster, P. I. and Buick, R. 2020. Exploring cycad foliage as an archive of the isotopic composition of atmospheric nitrogen. Geobiology.18(2):152-166. [ Links ]
Lopes, J. D. S. 2015. Crescimento e compostos nitrogenados de plantas jovens noduladas de Inga macrophylla benth. fertilizadas com diferentes formas de nitrogênio. In IV Congresso de Iniciação Científica do INPA-CONIC. Amazonia. Brasil. 410-415 pp. [ Links ]
Lu, M. Z.; Carter, A. M. and Tegeder, M. 2021. Altering ureide transport in nodulated soybean results in whole-plant adjustments of metabolism, assimilate partitioning, and sink strength. Journal of Plant Physiology. 269(1):153613. https://doi.org/10.1016/j.jplph.2021.153613. [ Links ]
McCoy, J. M.; Kaur, G.; Golden, B. R.; Orlowski, J. M.; Cook, D. R.; Bond, J. A. and Cox, M. S. 2018. Nitrogen fertilization of soybean affects root growth and nodulation on two soil types in Mississippi. Communications in Soil Science and Plant Analysis. 49(2):181-187. [ Links ]
Meena, V. S.; Mishra, P. K.; Bisht, J. K. and Pattanayak, A. 2017. Agriculturally important microbes for sustainable agriculture. Applications in crop production and protection. Springer. 2:81-128. https://doi.org/10.1007/978-981-10-5343-6Ohyama. [ Links ]
Ono, Y.; Fukasawa, M.; Sueyoshi, K.; Ohtake, N.; Sato, T.; Tanabata, S. and Ohyama, T. 2021. Application of Nitrate, Ammonium, or Urea Changes the Concentrations of Ureides, Urea, Amino Acids and Other Metabolites in Xylem Sap and in the Organs of Soybean Plants (Glycine max (L.) Merr.). International journal of molecular sciences. 22(9):4573. [ Links ]
Ortez, O.; Salvagiotti, F; Adee, E.; Enrico, J. and Ciampitti, I. A. 2017. “Soybean: Genetic Gain × Fertilizer Nitrogen Interaction”, Kansas Agricultural Experiment Station Research Reports. 3(6):6-21. https://doi.org/10.4148/2378-5977.7438. [ Links ]
Saturno, D. F.; Cerezini, P.; Moreira, S. P.; Oliveira, A. B.;Oliveira, M. C. N.; Hungria, M. and Nogueira, M. A. 2017. Mineral nitrogen impairs the biological nitrogen fixation in soybean of determinate and indeterminate growth types. Journal of Plant Nutrition. 40(12):1690-1701. https://doi.org/10.1080/01904167.2017.1310890. [ Links ]
Santachiara, G.; Borrás, L.; Salvagiotti, F.; Gerde, J. A. and Rotundo, J. L. 2017. Relative importance of biological nitrogen fixation and mineral uptake in high yielding soybean cultivars. Plant and Soil, 418(1):191-203. [ Links ]
Senthilkumar, M.; Amaresan, N. and Sankaranarayanan, A. 2021. Colorimetric analysis of ureide-N, amino-N, and nitrate-nitrogen Plant-Microbe Interactions.. Humana, New York. 49-52 pp. [ Links ]
Soumare, A.; Diedhiou, A. G.; Thuita, M. and Hafidi, M. 2020. Exploiting Biological Nitrogen Fixation. A Route Towards a Sustainable Agriculture. Plants. 9(8):1-22. doi: 10.3390/plants9081011. [ Links ]
Stal, L. J. 2017. The effect of oxygen concentration and temperature on nitrogenase activity in the heterocystous cyanobacterium Fischerella sp. Scientific Reports. 7(1):1-10. [ Links ]
Tamagno, S. X.; Sadras, V. O.; Haegele, J. W.; Armstrong, P. R. and Ciampitti, I. A. 2018. Interplay between nitrogen fertilizer and biological nitrogen fixation in soybean: implications on seed yield and biomass allocation. Scientific Reports . 8(1):1-11. [ Links ]
Tirado, T. J. L. y Alcántar, G. G. 1989. Dinámica del nitrógeno en plantas de soya. TERRA. 7(2):133-142. [ Links ]
Tirado, T. J. L.; Conejero, G. G. y Alcántar, G. G. 1990. Efecto del nitrógeno mineral y el déficit hídrico sobre las dos vías de asimilación del nitrógeno en plantas de soya. TERRA . 8(2):182-191. [ Links ]
Xia, X. Ma, C.; Dong, S.; Xu, Y. and Gong, Z. 2017. Effects of nitrogen concentrations on nodulation and nitrogenase activity in dual root systems of soybean plants. Soil Science and Plant Nutrition. 63(5):470-482. [ Links ]
Yan, Z.; Li, P.; Chen, Y.; Han, W. and Fang, J. 2016. Nutrient allocation strategies of woody plants: an approach from the scaling of nitrogen and phosphorus between twig stems and leaves. Scientific reports. 6(1):1-9. [ Links ]
Yoneyama, T.; Fujiwara, H. and Wilson, J. M. 2020. Variations in fractionation of carbon and nitrogen isotopes in higher plants: N metabolism and partitioning in phloem and xylem. Stable isotopes. Environmental Plant Biology. New York. 99-109 pp. [ Links ]
Zhou, W. J.; Ji, H. L.; Zhu, J. X.; Zhang, Y. P.; Sha, L. Q.; Liu, Y. T.; Zhang, X.; Zhao, W.; Dong, Y. X.; Bai, X. L.; Lin, Y. X.; Zhang, J. H. and Zheng, X. H. 2016. The effects of nitrogen fertilization on N2O emissions from a rubber plantation. Scientific Reports . 6(1):1-12. https://doi.org/10.1038/srep28230. [ Links ]
Received: May 01, 2022; Accepted: August 01, 2022
 Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
