










Services on Demand
Journal
Article
 text in
text in  English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista mexicana de ciencias agrícolas
Print version ISSN 2007-0934
Rev. Mex. Cienc. Agríc vol.13 n.6 Texcoco Aug./Sep. 2022 Epub Feb 24, 2023
https://doi.org/10.29312/remexca.v13i6.2800
Articles
Quality of light from fluorescent lamps in cucumber growth and severity of Oidium sp.
1Doctorado en Ciencias Agropecuarias-Facultad de Medicina Veterinaria y Zootecnia-Universidad Autónoma de Sinaloa. Boulevard San Ángel s/n, fraccionamiento San Benito, predio Las Coloradas, Culiacán, Sinaloa, México. CP. 80246. (norma-zazueta2812@hotmail.com).
2Facultad de Agronomía-Universidad Autónoma de Sinaloa. Carretera Culiacán-Eldorado km 17.5, Culiacán, Sinaloa, México. AP. 25. CP. 80000. (moisesyj@uas.edu.mx; teresadejesus-v@yahoo.com.mx; clopezorona@uas.edu.mx).
3 Dirección de Gestión de Investigaciones Científicas-Universidad Central del Este. Ave. Francisco Alberto Caamaño Deñó, San Pedro de Macorís, República Dominicana. CP. 21000. (tdiaz10@hotmail.com).
The quality of light affects the development of plants, due to the specific effects on photosynthesis, photomorphogenesis and other physiological and biochemical processes. It also has an important role in plant-pathogen interactions and controls various metabolic activities of fungi that determine their pathogenicity and severity. Three experiments were carried out under completely randomized designs to know the influence of fluorescent lamps of cool, neutral and warm white light, on the morphology and growth of cucumber plants (Cucumis sativus L.) and the severity of powdery mildew (Oidium sp.). In the growth chambers used, the photosynthetic photon flux density (PPFD) averaged 305 μmol m-2 s-1, but the spectral parameters related to red light (R:WWL> NWL> CWL) and blue light (B:CWL> NWL> WWL) were contrasting. The highest absolute amount of light R (122.04 μmol m-2 s-1), relative amount of R:PPFD (40.09%) and proportional amount of R:B (2.67) and R:FR (3.25) of WWL promoted greater height, leaf area, fresh and dry weight of leaves, stem and root of plants, while the greater absolute amount of light B (84.19 μmol m-2 s-1), relative amount of B:PPFD (27.48%) and proportional amount of B:R (1.04) and B:FR (2.65) of CWL induced lower plant height and greater stem thickness and leaf greenness index. The spectral parameters of CWL also disturbed the development of Oidium sp., which was reflected in less severity of powdery mildew compared to NWL or WWL.
Keywords: Cucumis sativus L.; blue light; powdery mildew; red light.
La calidad de la luz afecta el desarrollo de plantas, por los efectos específicos sobre la fotosíntesis, fotomorfogénesis, procesos fisiológicos y bioquímicos. También tiene un papel importante en las interacciones planta-patógeno y controla varias actividades metabólicas de hongos que determinan su patogenicidad y severidad. Se realizaron tres experimentos bajo diseños completamente al azar para conocer la influencia de lámparas fluorescentes de luz blanca fría, neutra y cálida, sobre la morfología y crecimiento de plantas de pepino (Cucumis sativus L.) y la severidad de la cenicilla (Oidium sp.). En las cámaras de crecimiento utilizadas, la densidad de flujo de fotones fotosintéticos (DFFF) promedió 305 μmol m-2 s-1, pero los parámetros espectrales relacionados con la luz roja (R:LBC> LBN> LBF) y la luz azul (A:LBF> LBN> LBC) fueron contrastantes. La mayor cantidad absoluta de luz R (122.04 µmol m-2 s-1), cantidad relativa de R:DFFF (40.09%) y cantidad proporcional de R: A (2.67) y R:RL (3.25) de LBC promovieron mayor altura, área foliar, peso fresco y seco de hojas, tallo y raíz de las plantas, mientras que, la mayor cantidad absoluta de luz A (84.19 µmol m-2 s-1), cantidad relativa de A:DFFF (27.48%) y cantidad proporcional de A:R (1.04) y A:RL (2.65) de LBF indujeron menor altura de planta y mayor grosor de tallo e índice de verdor foliar. Los parámetros espectrales de LBF también perturbaron el desarrollo de Oidium sp., que se reflejó en menor severidad de la cenicilla en comparación con LBN o LBC.
Palabras claves: Cucumis sativus L.; cenicilla; luz azul; luz roja.
Introduction
Light quality, photon flux and photoperiod are aspects of light that affect plant development, because of the specific effects they have on different types of plant responses, such as photosynthesis, photomorphogenesis and other physiological and biochemical processes (Hogewoning et al., 2010; Nelson and Bugbee, 2015; Snowden et al., 2016; Yan et al., 2019). The compact fluorescent lamp represents a simple and inexpensive way to replace the incandescent lamp used in growth chambers (Runkle et al., 2012).
The spectrum of light emitted by a compact fluorescent lamp is different from that of an incandescent lamp. An incandescent lamp emits more far-red light (FR=700-800 nm) than red light (R= 600-700 nm) and has a low R:FR ratio of approximately 0.7 (Runkle et al., 2012; Gupta and Jatothu, 2013), which causes stem elongation, leaf expansion and other responses induced by phytochromes (Casal, 2013; Demotes-Mainard et al., 2016). In contrast, the compact fluorescent lamp emits more blue light (B= 400-500 nm) and red light, and little far-red light, so the R:FR ratio is between 3 and 8, depending on the wave bands used and the lamp model, which induce more compact plant growth (Runkle et al., 2012; Gupta and Jatothu, 2013).
Light-emitting diodes (LEDs) emerged as a novel and efficient light source to promote plant growth in space-limited research chambers (Cope and Bugbee, 2013). With LEDs, monochromatic red or blue light can be provided, but none of these manage to satisfy the requirement of normal plant growth (Wang et al., 2016; Yang et al., 2017). However, when adequate R:B ratios are supplied, supplemented or not with other wavelengths: UV-A (350-400 nm), green (500-600 nm) or far red, greater compactness in tomato seedling is obtained (Javanmardi and Emami, 2013; Hernández et al., 2016), greater biomass, chlorophyll and photosynthetic capacity of seedlings of cucumber (Hogewoning et al., 2010; Song et al., 2017) and tomato (Xiaoying et al., 2012), optimal production of seedlings of lettuce (Yan et al., 2019) and potato (Chen et al., 2020).
However, several studies have reported higher growth under fluorescent lamps compared to LEDs with the same photonic flux (Lin et al., 2013; Chen et al., 2014; Wang et al., 2014). Fluorescent lamps have more diffused light compared to direct light from LEDs. Diffused light penetrates the plant canopy better than direct light and increases photosynthesis and dry biomass (Li and Yang, 2015). Fluorescent lamps also increase infrared radiation (Nelson and Bugbee, 2015) and have some FR radiation (Snowden et al., 2016), which increases leaf and petiole expansion and therefore radiation capture.
A variety of compact fluorescent lamps are available, which differ in luminous flux (lm); luminous efficacy (lm W-1), lifespan (h), color rendering index (CRI) and color appearance, most commonly described as color temperature and expressed in Kelvin units (K). Of which there is greater availability in three main groups: 2 700 to 3 000 K, which produces warm white light similar to that of incandescent lamps, 3 500 to 4 100 K of neutral or natural white light and 5 000 to 6 500 K of cool white light, which provides light with a blue hue (Saavedra et al., 2016).
Light is an important environmental factor also for fungi, which has an important role in plant-pathogen interactions and controls various metabolic activities of pathogenic fungi (Rahman et al., 2003; Wang et al., 2010). More than 100 species of fungi, which represent all phyla, have been found to respond to the effect of light (Tisch and Schmoll, 2010). The metabolic activities it regulates include circadian rhythms, asexual conidiation, pigmentation, secondary metabolism, and sexual development (Purschwitz et al., 2006; Suzuki et al., 2018).
Although the interaction between light and fungi has been studied and reviewed by several researchers (Idnurm and Heitman, 2005; Purschwitz et al., 2006; Chen et al., 2009), reports on the effect of light quality on the pathogenicity or virulence of fungi are limited. The objective of the research was to determine the effect of the light spectrum emitted by compact fluorescent lamps of cool, neutral and warm white light, on the growth of cucumber plants (Cucumis sativus L.) and the severity of powdery mildew of cucurbits (Oidium sp.).
Materials and methods
The research was conducted inside growth chambers of 44 x 70 x 80 cm (246 400 cm3) with woven mesh of 16 x 16 crystalline monofilaments of high-density polyethylene per cm2, on all their sides and sides lined with high reflectance Mylar paper (Figure 1) at the Faculty of Agronomy of the Autonomous University of Sinaloa. Seeds of ‘Poinsett 76’ cucumber were sown in polystyrene trays of 128 cavities and when the seedlings had two true leaves, they were individually transplanted into polystyrene cups of 0.5 L. In both types of containers, peat (Pro-Mix® FLX, Premier Horticulture, USA) was used as a substrate and they were irrigated to saturation with a fertilizer solution (pH= 6.5 to 7 and EC= 1.12 to 1.35 dS m-1) composed of 0.5 to 1.2 g L-1 of monopotassium phosphate and 0.3 to 0.7 g L-1 of potassium nitrate.

Figure 1 Cucumber plants inside a growth chamber with lighting of fluorescent lamps of cool (CWL, left), neutral (NWL, center) and warm white light (WWL, right).
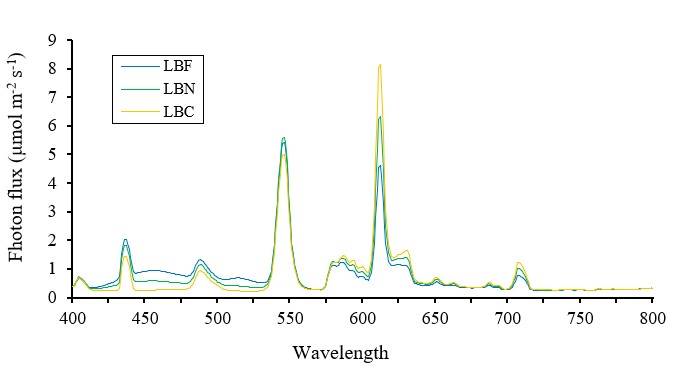
Fluorescent lamps (FLE23HLX, GE, USA) of cool (CWL= 6 500 K), neutral (NWL= 4 000 K) and warm white light (WWL= 2700 K) were used. The measurement of the photon flux, in the range of 350 to 1 050 nm, was performed with a spectroradiometer (Field SpecPro®Vnir, ASD, USA), obtaining: a) absolute amount of the photosynthetic photon flux density (PPFD= 400 to 700 nm), blue light (B= 400 to 500 nm), red light (R= 600 to 700 nm) and far-red light (FR= 700 to 800 nm); b) relative amount (per cent of total PPFD) of blue light (B:PPFD) and red light (R:PPFD); and c) proportional amount of blue-to-red light (B:R), blue-to-far-red light (B:FR), red-to-blue light (R:B), and red-to-far-red light (R:FR) (Table 1). The light spectrum of the three fluorescent lamps is shown in (Figure 2).
Table 1 Quality parameters of light emitted by fluorescent lamps of cool (CWL), neutral (NWL) and warm white light (WWL).
| Parameter/type of lamp | CWL | NWL | WWL |
| PPFD (400-700 nm)x | 306.4 | 305.25 | 304.37 |
| B (400-500 nm)x | 84.19 | 64.87 | 45.74 |
| R (600-700 nm)x | 81.34 | 100.44 | 122.04 |
| FR (700-800 nm)x | 31.79 | 33.93 | 37.51 |
| B:PPFD [(400-500/400-700 nm)*100]y | 27.48 | 21.25 | 15.03 |
| R:PPFD [(600-700/400-700 nm)*100]y | 26.55 | 32.9 | 40.1 |
| B:R (400-500/600-700 nm)z | 1.04 | 0.65 | 0.37 |
| B:FR (400-500/700-800 nm)z | 2.65 | 1.91 | 1.22 |
| R:B (600-700/400-500 nm)z | 0.97 | 1.55 | 2.67 |
| R:FR (600-700/700-800 nm)z | 2.56 | 2.96 | 3.25 |
PPFD= photosynthetic photon flux density; B= blue light; R= red light; FR= far-red light. Absolutex (μmol m-2 s-1), relativey (%) and proportionalz (dimensionless) quantities.
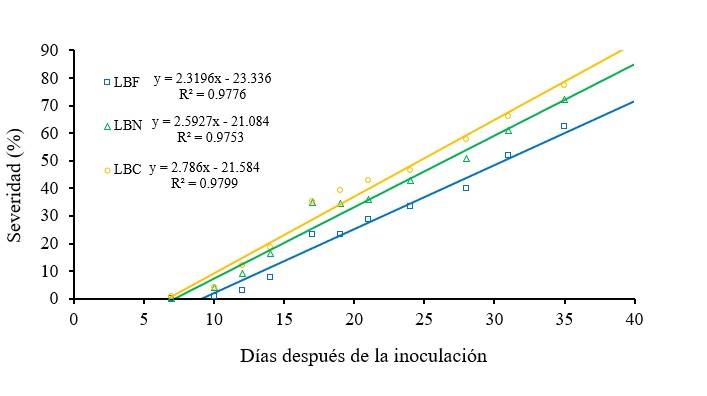
Figure 2 Spectral distribution of light emitted by fluorescent lamps of cool (LBF), neutral (LBN) and warm white light (LBC).
The luminous environment had means of 305.3 μmol m-2 s-1 of PPFD and 13.2 mol m-2 d-1 of integrated daily light, suitable for the production of vegetable seedlings (Fan et al., 2013). The photoperiod was 12/12 h of light/darkness. The temperature and relative humidity, recorded with thermohygrometers (CM-DT171, Twilight, Mexico), had means ± standard error of 24.5 ±0.15 °C and 62.5 ± 0.6%, respectively. The CO2 level was 420 ±20 μmol mol-1, obtained with CO2 meter (CO2-100, Amprobe, Germany).
For the study of plant growth, a completely randomized experimental design was used, with three treatments: compact fluorescent lamps of cool (CWL), neutral (NWL) and warm white light (WWL) and four repetitions (four plants per repetition). Cucumber plants were evaluated for 35 days after emergence (dae), a period during which they were exposed to lamp light. The experiment was repeated three times, in each of which the following response variables were evaluated: plant height, measured with tape measure, stem diameter, obtained with digital caliper (6MP, Truper, Mexico), foliar greenness, by means of a chlorophyll meter (SPAD 502 Plus, Minolta, Japan), leaf area, obtained with a non-destructive method proposed by Blanco and Follegati (2003), fresh and dry biomass of leaves, stem and root per plant, by analytical balance (SA120, Scientech, USA), after drying in an oven (FE293AD, Felisa, Mexico) at 70 °C, up to constant dry weight.
For the study of the severity of powdery mildew, a completely randomized experimental design was used, with three treatments: compact fluorescent lamps of cool (CWL), neutral (NWL) and warm white light (WWL) and 12 repetitions (one plant per repetition). The primary inoculum of Oidium sp. was obtained from naturally infected Cucurbita pepo L. plants.
The conidia suspension was prepared by brushing the source leaves with distilled water and maintained at a concentration of approximately 5.1 x 104 conidia ml-1 by hemocytometer (79003, Cole-Parmer, USA). Inoculation was carried out by foliar spraying of 30 ml of conidial suspension for each tray of 128 seedlings, the day they unfurled the second true leaf. The experiment was repeated three times, and each spanned a period of 35 days after inoculation (dai), during which the percentage of leaf area with symptoms of the disease on each leaf of the plants was determined, as well as the average of the entire plant.
The data obtained from the three experiments were averaged and subjected to the analysis of variance and comparison of means with the Tukey test (p≤ 0.05), using the statistical package Statistica version 7.0 (StatSoft, 2004).
Results and discussion
The quality of light emitted by fluorescent lamps (Table 1) caused significant differences (p≤ 0.05) in the height, stem diameter, leaf area and greenness of cucumber plants (Table 2). The luminous environment created by WWL lamps showed the highest values of absolute amount of red light (122.04 μmol m-2 s-1), R:PPFD ratio (40.1%) and R:B ratio (2.67), which caused increases in plant height from 3.3 to 30.2% and 15.4 to 82.3%, compared to the effect caused by NWL and CWL lamps, which emitted 17.5 and 45.1% less red light than WWL, respectively. The result of increased plant height due to red light is consistent with that described in Paeonia suffruticosa (Ding et al., 2010), Solanum lycopersicum (Xiaoying et al., 2012), Morus alba (Hu et al., 2016), Camptotheca acuminata (Yu et al., 2017) and Solanum tuberosum (Chen et al., 2020).
Phytochromes, red and far-red light receptors, regulate stem elongation by both cell division and extension (Neff et al., 2000). The activity of phytochromes during cell elongation is controlled by the biosynthesis of gibberellins AG1 and AG4, mainly, and auxin IAA (Damayanthi-Ranwala and Decoteau, 1998; Kurepin et al., 2007; Fukuda et al., 2016; Li et al., 2017).
The CWL lamps emitted the highest absolute amount of blue light (84.19 μmol m-2 s-1), 29.8 and 90.3% higher compared to the respective NWL and WWL lamps, which contained the elongation of the stem, as it was negatively correlated with plant height. These lamps also produced the highest B:PPFD ratio (27.48%) and the highest B:R ratio (1.04), in addition to the lowest R: PPFD ratio (26.55%) and the lowest R:B ratio (0.97). This result is related to the ability of blue light to inhibit stem elongation (Ding et al., 2010; Xiaoying et al., 2012; Hu et al., 2016; Yu et al., 2017; Chen et al., 2020), in response to synergistic interactions between phytochromes and cryptochromes, red and blue light receptors; respectively, in the promotion or inhibition of stem elongation (Heo et al., 2002).
The spectral characteristics of the light emitted by the CWL lamps, referred to above, also increased the stem diameter from 0.1 to 9.1% and 5.1 to 13.2% compared to that of the plants that grew with light from NWL and WWL lamps, respectively, which emitted light with a B:PPFD ratio (15.03 and 21.25%) and a B:R ratio (0.37 and 0.65) lower than those of CWL. Li et al. (2017); Chen et al. (2020) point out that blue light stimulates a greater expression of proteins associated with plant microtubules and tubulin, which promote secondary cell wall formation and atherefore thickening of the stem.
The quality of light emitted by WWL lamps, with relevance in red light, promoted increases of 2.2 to 14.2% and 18.6 to 25.5% in the leaf area per seedling, compared to the effect caused by NWL and CWL lamps, respectively.
Consistent result with those obtained by Cope and Bugbee (2013) in Raphanus sativus and Glycine max, and by Hernández and Kubota (2016) in Cucumis sativus, where the leaf area grew as blue light decreased and red light increased. Both blue and red light stimulate the flux of protons in cells, apoplastic acidification, cell wall elasticity and the accumulation of solutes for the maintenance of the turgor of growing leaves, through separate mechanisms. Blue light induces a direct interaction between the proton pump and a blue light photoreceptor, while red light indirectly influences the proton pump, modulating calcium and potassium channels (Staal et al., 1994; Van Volkenburgh, 1999).
While the spectral parameters in the light emitted by CWL (Table 1) with relevance in blue light, increased the foliar greenness of the plants from 2.1 to 8% and 10.1 to 18% compared to the effect caused by the NWL and WWL lamps, respectively. This is consistent with studies conducted on seedlings of Solanum lycopersicum (Hernández and Kubota, 2016) and Cucumis sativius (Hogewoning et al., 2010; Hernández et al., 2016), which showed a higher concentration of foliar chlorophyll as the flux of blue photons increased, due to the additive effect of cryptochromes and phytochromes, compared to lower biosynthesis of chlorophyll in plants grown under monochromatic blue or red light.
Table 2 Plant height (PH), stem diameter (SD), leaf area per plant (LAP) and leaf greenness (LG) of cucumber plants cv Poinsett 76 grown in a growth chamber with lamps of cool (CWL), neutral (NWL) and warm white light (WWL).
| Parameter | Treatment | Days after the emergence | |||||||||
| 7 | 14 | 21 | 28 | 35 | |||||||
| PH (cm) | CWL | 3.98 | c | 4.5 | c | 11.7 | b | 17.8 | c | 43.3 | b |
| NWL | 5.4 | b | 5.59 | b | 12.5 | b | 26.2 | b | 48.4 | ab | |
| WWL | 6.92 | a | 7.28 | a | 14.7 | a | 32.4 | a | 50 | a | |
| HMSD | 0.97 | 0.96 | 1.5 | 5.1 | 5.48 | ||||||
| SD (mm) | CWL | 2.3 | a | 3.3 | a | 4.4 | a | 5.83 | a | 5.98 | a |
| NWL | 2.1 | b | 3.2 | ab | 4.4 | a | 5.34 | b | 5.51 | b | |
| WWL | 2.1 | b | 2.9 | b | 4.2 | b | 5.21 | b | 5.42 | b | |
| HMSD | 0.13 | 0.32 | 0.16 | 0.2 | 0.31 | ||||||
| LAP (cm2) | CWL | 30 | b | 163.6 | b | 529.5 | b | 936.1 | b | 1319.3 | b |
| NWL | 48.4 | a | 200.9 | a | 579.6 | b | 1065.3 | a | 1525.1 | a | |
| WWL | 37 | b | 205.3 | a | 661.7 | a | 1114.8 | a | 1564.4 | a | |
| HMSD | 8.52 | 27.4 | 59.28 | 103.6 | 119.4 | ||||||
| LG (SPAD u) | CWL | 30.1 | a | 41.6 | a | 43.5 | a | 41.5 | a | 36.4 | a |
| NWL | 27.9 | ab | 39.7 | b | 42 | a | 38.4 | b | 35.7 | a | |
| WWL | 25.5 | b | 37.8 | c | 37.8 | b | 36.1 | c | 33 | b | |
| HMSD | 2.94 | 1.84 | 1.63 | 1.99 | 1.82 | ||||||
Means with the same letter are statistically equal (Tukey, p≤ 0.05). HMSD= honest minimum significant difference.
The production of biomass of leaves, stem and roots by cucumber plants also showed significant differences (p≤ 0.05) due to the quality of light emitted by fluorescent lamps (Table 3).
Table 3 white light (Fresh weight (FW) and dry weight (DW) of leaves, stem and root of cucumber plants cv Poinsett 76 grown in a growth chamber with lamps of cool (CWL), neutral (NWL) and warm WWL).
| Parameter | Treatment | Days after the emergence | |||||||||
| 7 | 14 | 21 | 28 | 35 | |||||||
| FW/Leaves (g) | CWL | 0.17 | b | 3.93 | a | 11.52 | b | 16.21 | b | 20.18 | b |
| NWL | 0.2 | ab | 5.22 | a | 12.24 | ab | 16.25 | b | 20.97 | b | |
| WWL | 0.23 | a | 5.43 | a | 13.29 | a | 17.99 | a | 22.69 | a | |
| HMSD | 0.03 | 1.57 | 1.42 | 1.02 | 1.4 | ||||||
| DW/leaves (g) | CWL | 0.02 | b | 0.37 | b | 1.06 | a | 1.58 | b | 2.09 | b |
| NWL | 0.02 | b | 0.47 | a | 1.11 | a | 1.68 | ab | 2.21 | ab | |
| WWL | 0.03 | a | 0.47 | a | 1.14 | a | 1.8 | a | 2.49 | a | |
| HMSD | 0.006 | 0.096 | 0.109 | 0.167 | 0.291 | ||||||
| FW/Stem (g) | CWL | 0.41 | b | 1.58 | b | 5.18 | a | 7.12 | a | 8.06 | b |
| NWL | 0.43 | ab | 2.36 | a | 5.18 | a | 7.1 | a | 9.02 | a | |
| WWL | 0.49 | a | 2.8 | a | 5.31 | a | 7.42 | a | 9.53 | a | |
| HMSD | 0.068 | 0.598 | 1.086 | 0.963 | 0.878 | ||||||
| DW/Stem (g) | CWL | 0.03 | b | 0.04 | a | 0.18 | a | 0.34 | b | 0.5 | b |
| NWL | 0.03 | b | 0.05 | a | 0.17 | a | 0.37 | ab | 0.56 | a | |
| WWL | 0.04 | a | 0.05 | a | 0.22 | a | 0.39 | a | 0.58 | a | |
| HMSD | 0.01 | 0.019 | 0.073 | 0.046 | 0.056 | ||||||
| FW/Root (g) | CWL | 0.42 | b | 0.99 | b | 2.05 | b | 5.03 | b | 8.01 | b |
| NWL | 0.55 | a | 1.21 | a | 2.07 | b | 5.33 | b | 8.59 | ab | |
| WWL | 0.5 | a | 1.2 | a | 2.62 | a | 5.81 | a | 9 | a | |
| HMSD | 0.058 | 0.12 | 0.15 | 0.429 | 0.712 | ||||||
| DW/Root (g) | CWL | 0.018 | a | 0.065 | b | 0.085 | b | 0.213 | b | 0.341 | b |
| NWL | 0.019 | a | 0.07 | ab | 0.105 | a | 0.263 | a | 0.42 | a | |
| WWL | 0.017 | a | 0.075 | a | 0.1 | a | 0.256 | a | 0.413 | a | |
| HMSD | 0.004 | 0.006 | 0.006 | 0.023 | 0.04 | ||||||
Means with the same letter are statistically equal (Tukey, p≤ 0.05). HMSD= honest minimum significant difference.
The red light parameters (R= 122.04 μmol m-2 s-1; R:PPFD= 40.9%; R:B= 2.67) of WWL lamps caused plants to increase the fresh leaf weight, from 3.9 to 15.4% compared to the effect caused by NWL lamps (R= 100.44 μmol m-2 s-1; R:PPFD= 32.9%; R:B= 1.55) and from 8.6 to 38% in relation to CWL (R= 81.34 μmol m-2 s-1; R:PPFD= 26.55%; R:B= 0.97). With WWL, the dry weight of leaves also increased from 0 to 12.6% and from 8.9 to 27.4%, with respect to that obtained from plants grown with NWL and CWL, respectively.
The morphological response influenced those of plant growth, since the fresh and dry biomass of leaves (growth parameters) coincided closely with the leaf area per plant (morphological parameter), as observed by Hogewoning et al. (2010); Hernández and Kubota (2016). These authors indicated that the seedlings of Cucumis sativus, grown under combinations of red and blue light, require 15 to 50% blue light to have adequate growth and development, since such proportions of light were associated with increases in biomass per unit of leaf area, foliar nitrogen and chlorophyll content, photosynthetic assimilation of CO2 and stomatal conductance, which were lower with monochromatic red light, where plants showed a dysfunctional photosynthesis system.
The accumulation of biomass in the stem of the plants did not present significant differences (p≤ 0.05) during the first 28 days after the emergence (dae). However, at 35 dae, the stem biomass was more related to its length (plant height) than to its diameter, since the red-light parameters of the WWL lamps caused increases of 5.7 and 18.2% in the fresh weight of the stem and 0 and 12.2% in the dry weight of the stem, compared to those obtained with NWL and CWL lamps, respectively, where the stems elongated less and thickened more.
In this sense Ayala-Tafoya et al. (2015) observed increases in the dry weight of leaves and stem of Cucumis sativus plants grown under red mesh, due to the confluence of more PPFD and red light, compared to responses to blue mesh. Similarly, after 21 dae, with WWL lamps, the fresh root weight increased from 4.7 to 26.7% and from 12.4 to 28.1%, compared to NWL and CWL lamps, respectively. While, with WWL and NWL lamps, the dry root weight exceeded from 17.6 to 21% that obtained with CWL lamps.
Cucumber plants grown with CWL lamps showed a severity of powdery mildew (Oidium sp.) of 0.8% at 10 days after inoculation (dai) and 3% at 12 dai, which was lower from 4.9 and 5.3 times to 3 and 4 times compared to plants grown with NWL and WWL lamps, respectively. In the rest of the study, CWL lamps induced severity values of 7.9% at 14 dai and 62.5% at 35 dai, lower from 109.2 and 136.2% to 15.7 and 23.6%, compared to NWL and WWL lamps, respectively. However, the severity values of powdery mildew obtained with the three types of fluorescent lamps, throughout the study period, showed linear increases with similar coefficients of determination (Figure 3).
In the same sense, other studies showed that blue light reduced the severity of Botrytis cinerea in Solanum lycopersicum (Xu et al., 2017) and Podosphaera xanthii in Cucumis melo (Jing et al., 2018), by increasing the expression of defense-related genes in plants, which induced accumulation of: proline, H2O2, phenolic compounds, flavonoids, tannins and lignin, in addition to promoting a compact morphology and increased thickness of the cell wall in plant tissue.
Conclusions
Compact fluorescent lamps of warm white light promoted greater height, leaf area, fresh and dry weight of leaves, stem and root of plants. While compact fluorescent lamps of cool white light induced lower plant height and greater stem thickness and leaf greenness index. The spectral parameters of compact fluorescent lamps of cool white light also induced lower severity of the powdery mildew in cucumber plants, compared to lamps of neutral or warm white light.
Literatura citada
Ayala, T. F.; Yáñez, J. M. G.; Partida, R. L.; Ruiz, E. F. H.; Campos, G. H.; Vásquez, M. O.; Velázquez, A. T. J. y Díaz, V. T. 2015. Producción de pepino en ambientes diferenciados por mallas de sombreo fotoselectivo. ITEA. 1(111):3-17. https://doi.org/10.12706 /itea.2015.001. [ Links ]
Casal, J. J. 2013. Photoreceptor signaling networks in plant responses to shade. Annu. Rev. Plant Biol. 1(64):403-427. https://doi.org/10.1146/annurev-arplant-050312-120221. [ Links ]
Chen, C. H.; Ringelberg, C. S.; Gross, R. H.; Dunlap, J. C. and Loros, J. J. 2009. Genome-wide analysis of light-inducible responses reveals hierarchical light signalling in Neurospora. EMBO J. 8(28):1029-1042. https://dx.doi.org/10.1038%2Femboj.2009.54. [ Links ]
Chen, L.; Zhang, K.; Gong, X.; Wang, H.; Gao, Y.; Wang, X.; Zeng, Z. and Hu, Y. 2020. Effects of different LEDs light spectrum on the growth, leaf anatomy, and chloroplast ultrastructure of potato plantlets in vitro and minituber production after transplanting in the greenhouse. J. Integr. Agric. 1(19):108-119. https://doi.org/10.1016/S2095-3119(19)62633-X. [ Links ]
Chen, X. L.; Guo, W. Z.; Xue, X. Z.; Wang, L. C. and Qiao, X. J. 2014. Growth and quality responses of ‘Green Oak Leaf’ lettuce as affected by monochromic or mixed radiation provided by fluorescent lamp (FL) and light-emitting diode (LED). Sci. Hortic. 1(172):168-175. https://doi.org/10.1016/j.scienta.2014.04.009. [ Links ]
Cope, K. R. and Bugbee, B. 2013. Spectral effects of three types of white light-emitting diodes on plant growth and development: absolute versus relative amounts of blue light. HortSci. 4(48):504-509. https://doi.org/10.21273/HORTSCI.48.4.504. [ Links ]
Damayanthi, R. N. K. and Decoteau, D. R. 1998. Involvement of gibberellins in phytochrome-regulated stem and petiole elongation in watermelon plants. HortSci. 3(33):493-494. [ Links ]
Demotes, M. S.; Péron, T.; Corot, A.; Bertheloot, J.; Le Gourrierec, J.; Pelleschi, T. S.; Crespel, L.; Morel, P.; Huché, T. L.; Boumaza, R.; Vian, A.; Guérin, V.; Leduc, N. and Sakr, S. 2016. Plant responses to red and far-red lights, applications in horticulture. Environ. Exp. Bot. 1(121):4-21. https://doi.org/10.1016/j.envexpbot.2015.05.010. [ Links ]
Ding, Y.; He, S.; Silva, J. A. T.; Li, G. and Tanaka, M. 2010. Effects of a new light source (cold cathode fluorescent lamps) on the growth of tree peony plantlets in vitro. Sci. Hortic. 125(2):167-169. https://doi.org/10.1016/j.scienta.2010.03.019. [ Links ]
Fan, X. X.; Xu, Z. G.; Liu, X. Y.; Tang, C. M.; Wang, L. W. and Han, X. I. 2013. Effects of light intensity on the growth and leaf development of young tomato plants grown under a combination of red and blue light. Sci. Hortic. 1(153):50-55. https://doi.org/10.1016/j. scienta.2013.01.017. [ Links ]
Fukuda, N.; Ajima, C.; Yukawa, T. and Olsen, J. E. 2016. Antagonistic action of blue and red light on shoot elongation in petunia depends on gibberellin, but the effects on flowering are not generally linked to gibberellin. Environ. Exp. Bot. 1(121):102-111. https://doi.org/10.1016/ j.envexpbot.2015.06.014. [ Links ]
Gupta, S. D. and Jatothu, B. 2013. Fundamentals and applications of light-emitting diodes (LEDs) in in vitro plant growth and morphogenesis. Plant Biotechnol. Rep. 3(7):211-220. http://dx.doi.org/10.1007/s11816-013-0277-0. [ Links ]
Heo, J.; Lee, C.; Chakrabarty, D. and Paek, K. 2002. Growth responses of marigold and salvia bedding plants as affected by monochromic or mixture radiation provided by a Light-Emitting Diode (LED). Plant Growth Regul. 3(38):225-230. https://doi.org/10.1023/A: 1021523832488. [ Links ]
Hernández, R.; Eguchi, T.; Deveci, M. and Kubota, C. 2016. Tomato seedling physiological responses under different percentages of blue and red photon flux ratios using LEDs and cool white fluorescent lamps. Sci. Hortic. 1(213):270-280. http://doi.org/10.1016/j.scienta. 2016.11.005. [ Links ]
Hernández, R. and Kubota, C. 2016. Physiological responses of cucumber seedlings under different blue and red photon flux ratios using LEDs. Environ. Exp. Bot. 1(121):66-74. http://dx.doi.org/10.1016/j.envexpbot.2015.04.001. [ Links ]
Hogewoning, S. W.; Trouwborst, G.; Maljaars, H.; Poorter, H.; Van Ieperen, W. and Harbinson, J. 2010. Blue light dose-responses of leaf photosynthesis, morphology, and chemical composition of Cucumis sativus grown under different combinations of red and blue light. J. Exp. Bot. 11(61):3107-3117. http://doi.org/10.1093/jxb/erq132. [ Links ]
Hu, J.; Dai, X. and Sun, G. 2016. Morphological and physiological responses of Morus alba seedlings under different light qualities. Not. Bot. Horti Agrobot. Cluj-Napoca. 2(44):382-392. https://doi.org/10.15835/nbha44210486. [ Links ]
Idnurm, A. and Heitman, J. 2005. Light controls growth and development via a conserved pathway in the fungal kingdom. PLoS Biol. 4(3):615-626. https://dx.doi.org/10.1371%2 Fjournal.pbio.0030095. [ Links ]
Javanmardi, J. and Emami, S. 2013. Response of tomato and pepper transplants to light spectra provided by light emitting diodes. Inter. J. Veg. Sci. 2(19):138-149. http://doi.org/10.1080/ 19315260.2012.684851. [ Links ]
Jing, X.; Wang, H.; Gong, B.; Liu, S.; Wei, M.; Ai, X.; Li, Y. and Shi, Q. 2018. Secondary and sucrose metabolism regulated by different light quality combinations involved in melon tolerance to powdery mildew. Plant Physiol. Biochem. 1(124):77-87. https://doi.org/10. 1016/j.plaphy.2017.12.039. [ Links ]
Kurepin, L. V.; Emery, R. J. N.; Pharis, R. P. and Reid, D. M. 2007. Uncoupling light quality from light irradiance effects in Helianthus annuus shoots: putative roles for plant hormones in leaf and internode growth. J. Exp. Bot. 58(8):2145-2157. https://doi.org/10.1093/jxb/ erm068. [ Links ]
Li, C.; Xu, Z. G.; Dong, R. Q.; Chang, S.; Wang, L. Z.; Khalil, U. R. M. and Tao, J. M. 2017. An RNA-seq analysis of grape plantlets grown in vitro reveals different responses to blue, green, red LED light, and white fluorescent light. Front. Plant Sci. 1(8):1-16. https://doi.org /10.3389/fpls.2017.00078. [ Links ]
Li, T. and Yang, Q. 2015. Advantages of diffuse light for horticultural production and perspectives for further research. Front. Plant Sci. 1(6):1-5. https://doi.org/10.3389/fpls.2015.00704. [ Links ]
Lin, K. H.; Huang, M. Y.; Huang, W. D.; Hsu, M. H.; Yang, Z. W. and Yang, C. M. 2013. The effects of red, blue, and white light-emitting diodes on the growth, development, and edible quality of hydroponically grown lettuce (Lactuca sativa L. var. capitata). Sci. Hortic. 1(150):86-91. https://doi.org/10.1016/j.scienta.2012.10.002. [ Links ]
Neff, M. C.; Fankhauser, J. and Chory, J. 2000. Light: an indicator of time and place. Genes Develop. 3(14):257-271. [ Links ]
Nelson, J. A. and Bugbee, B. 2015. Analysis of environmental effects on leaf temperature under sunlight, high pressure sodium and light emitting diodes. PLoS ONE. 10(10):1-13. https://doi.org/10.1371/journal.pone.0138930. [ Links ]
Purschwitz, J.; Muller, S.; Kastner, C. and Fischer, R. 2006. Seeing the rainbow: light sensing in fungi. Curr. Opin. Microbiol. 6(9):566-571. https://doi.org/10.1016/j.mib.2006.10.011. [ Links ]
Rahman, M. Z.; Honda, Y. and Arase, S. 2003. Red-light induced resistance in broad bean (Vicia faba L.) to leaf spot disease caused by Alternaria tenuissima. J. Phytopathol. 2(151):86-91. https://doi.org/10.1046/j.1439-0434.2003.00685.x. [ Links ]
Runkle, E. S.; Padhye, S. R.; Oh, W. and Getter, K. 2012. Replacing incandescent lamps with compact fluorescent lamps may delay flowering. Sci. Hortic. 1(143):56-61. https://doi.org/ 10.1016/j.scienta.2012.05.028. [ Links ]
Saavedra, E.; Rey, F. J. y Luyo, J. 2016. Sistemas de iluminación, situación actual y perspectivas. TECNIA. 2(26):44-62. http://dx.doi.org/10.21754/tecnia.v26i2.57. [ Links ]
Snowden, M. C.; Cope, K. R. and Bugbee, B. 2016. Sensitivity of seven diverse species to blue and green light: interactions with photon flux. PLoS One. 10(11):e0163121. http://dx.doi.org/10.1371/journal.pone.0163121. [ Links ]
Song, J.; Meng, Q. W.; Du, W. F. and He, D. 2017. Effects of light quality on growth and development of cucumber seedlings in controlled environment. Inter. J. Agric. Biol. Eng. 3(10):312-318. http://dx.doi.org/10.3965/j.ijabe.20171003.2299. [ Links ]
Staal, M.; Elzenga, J. T. M.; Van Elk, A. G.; Prins, H. B. A. and Van Volkenburgh, E. 1994. Red and blue light-stimulated proton efflux by epidermal leaf-cells of the Argenteum mutant of Pisum sativum. J. Exp. Bot. 9(45):1213-1218. http://dx.doi.org/10.1093/jxb/45.9.1213. [ Links ]
StatSoft. 2004. Statistica (data analysis software system), version 7. www.statsoft.com. [ Links ]
Suzuki, T.; Nishimura, S.; Yagi, K.; Nakamura, R.; Takikawa, Y.; Matsuda, Y.; Kakutani, K. and Nonomura, T. 2018. Effects of light quality on conidiophore formation of the melon powdery mildew pathogen Podosphaera xanthii. Phytoparasitica. 1(46):31-43. https://doi.org/10.1007/s12600-017-0631-9. [ Links ]
Tisch, D. and Schmoll, M. 2010. Light regulation of metabolic pathways in fungi. Appl. Microbiol Biotechnol. 5(85):1259-1277. https://dx.doi.org/10.1007%2Fs00253-009-2320-1. [ Links ]
Van Volkenburgh, E. 1999. Leaf expansion -an integrating plant behaviour. Plant Cell Environ. 12(22):1463-1473. http://dx.doi.org/10.1046/j.1365-3040.1999.00514.x. [ Links ]
Wang, H.; Jiang, Y. P.; Yu, H. J.; Xia, X. J.; Shi, K.; Zhou, Y. H. and Yu, J. Q. 2010. Light quality affects incidence of powdery mildew, expression of defense-related genes and associated metabolism in cucumber plants. Eur. J. Plant Pathol. 1(127):125-135. https://doi.org/ 10.1007/s10658-009-9577-1. [ Links ]
Wang, J.; Lu, W.; Tong, Y. and Yang, Q. 2016. Leaf morphology, photosynthetic performance, chlorophyll fluorescence, stomatal development of lettuce (Lactuca sativa L.) exposed to different ratios of red light to blue light. Front. Plant Sci. 1(7):1-10. https://doi.org/10.3389 /fpls.2016.00250. [ Links ]
Wang, X. Y.; Xu, X. M. and Cui, J. 2014. The importance of blue light for leaf area expansion, development of photosynthetic apparatus, and chloroplast ultrastructure of Cucumis sativus grown under weak light. Photosynthetica. 2(53):213-222. https://doi.org/10.1007/s11099-015-0083-8. [ Links ]
Xiaoying, L.; Shirong, G.; Taotao, C.; Zhigang, X. and Tezuka, T. 2012. Regulation of the growth and photosynthesis of cherry tomato seedlings by different light irradiations of light emitting diodes (LED). Afr. J. Biotechnol. 22(11):6169-6177. https://doi.org/10.5897/ AJB11.1191. [ Links ]
Xu, H.; Fu, Y.; Li, T. and Wang, R. 2017. Effects of different LED light wavelengths on the resistance of tomato against Botrytis cinerea and the corresponding physiological mechanisms. J. Integr. Agric. 16(1):106-114. https://doi.org/10.1016/S2095-3119(16) 61435-1. [ Links ]
Yan, Z.; He, D.; Niu, G. and Zhai, H. 2019. Evaluation of growth and quality of hydroponic lettuce at harvest as affected by the light intensity, photoperiod and light quality at seedling stage. Sci. Hortic. 1(248):138-144. https://doi.org/10.1016/j.scienta.2019.01.002. [ Links ]
Yang, Z.; He, W.; Mou, S.; Wang, X.; Chen, D.; Hu, X.; Chen, L. and Bai, J. 2017. Plant growth and development of pepper seedlings under different photoperiods and photon flux ratios of red and blue LEDs. Trans. Chin. Soc. Agri. Eng. 33(17):173-180. https://doi.org/10. 11975/j.issn.1002-6819.2017.17.023. [ Links ]
Yu, W.; Liu, Y., Song, L.; Jacobs, D. F.; Du, X.; Ying, Y.; Shao, Q. and Wu, J. 2017. Effect of differential light quality on morphology, photosynthesis, and antioxidant enzyme activity in Camptotheca acuminata seedlings. J. Plant Growth Regul. 1(36):148-160. https://doi.org/10.1007/s00344-016-9625-y [ Links ]
Received: January 01, 2022; Accepted: April 01, 2022
 Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
