











 text in
text in  English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkIntroducción
Las bacterias asociadas a la rizosfera, también llamadas promotoras del crecimiento vegetal (BPCV) cumplen funciones específicas en los procesos biogeoquímicos, interactúan con las plantas, establecen protección contra fitopatógenos, aumentan la disponibilidad de nutrientes y nivel de fertilidad del suelo (Martínez-Romero et al., 2018). Dentro de estas especies destaca el género Klebsiella, principalmente las especies de K. variicola y K. quasipneumoniae.
Cumplen mecanismos como la modulación del sistema radicular, promueven el fototropismo positivo, cumplen funciones específicas como la producción del ácido indol-3-acético, son fijadoras de nitrógeno atmosférico, solubilizadoras de fósforo (Wei et al., 2013; Vega Celedón et al., 2016), los cuales son esenciales para la formación de aminoácidos, compuestos nitrogenados para el metabolismo y parte estructural de la planta, vías de síntesis de fosfolípidos de membrana, elongación de la raíz, floración, parte del metabolismo para la obtención de energía, reparación y síntesis de ácidos grasos (Beltrán, 2014).
Otra de las funciones de las BPCV es estimular el desarrollo del sistema radicular de la planta y por ende aumentar la absorción de nutrientes, la principal vía de absorción se da por medio de los pelos radiculares presentes en las raíces primarias y secundarias, a través de los cuales ingresa una gran cantidad de compuestos y moléculas orgánicas para el metabolismo de la planta, se ha demostrado que existen microorganismos presentes en el suelo que han desarrollado la capacidad para internalizarse, translocar en el sistema vascular de la planta y persistir en el fruto como es el caso de E coli O157:H7 (Heaton et al., 2008; Ocaña et al., 2018).
La interacción y el metabolismo con otros microorganismos, permiten la asociación de comunidades microbianas que establecen simbiosis entre bacterias con hongos antagonistas que brindan protección a las plantas de fitopatógenos presentes del suelo. Entre los hongos antagonistas presentes, se encuentra Trichoderma comúnmente aislado del suelo, siendo ampliamente estudiado y comercializado como biopesticidas, biofertilizantes y enmiendas del suelo, que permiten el equilibrio del nicho ecológico. (Benítez et al., 2004; Do Nascimento et al., 2017; Hirpara et al., 2017). Se han utilizado varias estrategias para identificar los factores moleculares implicados en esta compleja interacción de simbiosis, incluida la genómica, la proteómica y más recientemente, la metabolómica, con el fin de mejorar nuestra comprensión (Vinale et al., 2008).
Se han evaluado biofertilizantes a base de soluciones microbianas en hortalizas, como Solanum lycopersicum conocido comúnmente como jitomate (SIAP, 2019). No obstante, es de vital importancia la vigilancia de la internalización, persistencia y translocación de estas bacterias en los tejidos internos de frutos con el fin de garantizar la sanidad del producto. Por ello, se analizó la translocación in vivo de K. variicola y K. quasipneumoniae desde el sistema radicular al fruto de S. lycopersicum DT-22 en invernadero.
Materiales y métodos
Aislamiento de cepas bacterianas de diversos ambientes
Se seleccionaron muestras de raíces de plátano, maíz y jitomate, y diversas muestras aguas (río, laguna, planta de tratamiento de aguas residuales y de riego) de los municipios de Mochitlán, Acapulco, Chilpancingo y Tixtla respectivamente. Se realizó un lavado con agua estéril de la raíz y se localizaron los nódulos. Las muestras de rizósfera se tomaron con hisopo de forma directa y se transportaron al laboratorio en solución salina estéril al 0.85% y en viales en tubo con agar MacConkey. De las muestras de aguase hicieron diluciones hasta 1x10-3, siguiendo la metodología de Puerta et al. (2010). Después de 24 h a temperatura ambiente, se seleccionaron colonias características del género Klebsiella, colonias lactosa positivas, de borde irregular y sumamente mucoides en forma de cúpula, lisas y convexas. Se les realizó las pruebas de oxidasa, y se evaluó el perfil bioquímico convencional: en Kligler, MIO, citrato, urea, lisina para separar al género.
Diferenciación molecular de K. variicola, K. pneumoniae y K. quasipneumoniae mediante PCR multiplex
Las cepas identificadas bioquimicante del género Klebsiella, realizó la extracción de ADN por choque térmico, solubilizando tres colonias en 50 µl de agua estéril y sometiendo a una temperatura de 96 °C por 10 min y rápidamente en hielo durante 5 min y posteriormente se centrifuga y se toman 10 µl del sobrenadante y se resuspendiócon 90 µl de agua destilada estéril y se mantienen a -20 °C. La mezcla de reacción para la PCR-multiplex se llevó a cabo a un volumen final de 25 µl, con una concentración 1X de buffer, 2 mM de dNTP’s, 25 mM de MgCl, final de 1X de buffer, 2 mM de dNTP’s, 25 mM de MgCl, 1µl de los oligonucleótido de K. variicola 2006 a una concentración de 10 pmol DNA, 2µl de K. quasipneumoniae 10441 a una concentración de 5 pmol y 2µl de los oligonucleótidos de K. pneumoniae 13883 a una concentración final de 5 pmol, 0.5µl de Taq ADN polimerasa y 3 μl de la solución de ADN al 1%.
La mezcla de reacción estuvo sujeta a un paso de desnaturalización inicial de 2 min, a 92 °C, seguida por 30 ciclos de 30 s, a 92 °C, 30 s, a 60 °C, 40 s, a 72 °C y una extensión final de 3 min, a 72 °C. Los productos de PCR fueron sometidos a electroforesis en un gel de agarosa al 1.5% a 120 V por 1 h. Se utilizaron como controles un fragmento de 275 pb de K. variicola 2006, uno de 372 pb K. quasipneumoniae 10 441 y de 650 pb de K. pneumoniae 13 883 (Martínez et al., 2018).
Detección de los mecanismos de promoción de crecimiento vegetal: fijación de nitrógeno
Las cepas se inocularon con palillo de madera estéril en medio Rennie modificado libre de N2 (Rennie, 1986) y se incubaron a 30 °C por 24-48 h. El crecimiento es indicador de la fijación de nitrógeno. Como control positivo se utilizó a K. variicola PB y negativo E. coli DH5-α.
Solubilización de fosfato
La capacidad de solubilizar fosforo inorgánico se evaluó en el medio National Botanical Research Institute’s Phosphate (NBRIP) con fosfato tricálcico siguiendo la metodología de Nautiyal, (1999), se midió el diámetro del halo y de la colonia (mm) para el cálculo del índice de solubilización (IS), que relaciona la suma de los diámetros de halo y colonia, y el diámetro de la colonia. Como control positivo se empleó K. variicola PB y como control negativo E. coli DH5-α.
Cuantificación del ácido indol acético
La producción de ácido indol acético (AIA), se evaluó en caldo soya tripticaseína suplementado con triptófano siguiendo la metodología e interpretación Lara et al. (2011). Para cuantificar la producción de AIA, en sobrenadantes de las cepas evaluadas, se realizó una curva patrón con diferentes concentraciones de AIA comercial (4 a 140 µg ml-1) y el reactivo de Salkowski relación 2:1. Se midió la absorbancia a 542 nm en un lector Stat Fax-2100, empleando como control positivo K. variicola PB y como control negativo E. coli DH5-α.
Producción de sideróforos
Se evaluó la producción de sideróforos creciendo las cepas en caldo F, adicionado con glicerol 10 ml L-1, a partir de un cultivo puro de 18-24 h, se incubo durante 24-72 h a 37 °C y se examinó el crecimiento bajo una luz ultravioleta ligera de 260 nm en un mini transiluminador BioRad. Se interpretó según Villa et al. (2017) empleando como control positivo K. variicola PB y control negativo caldo F sin inocular.
Material vegetal y diseño de los tratamientos
Las semillas de S. lycopersicum variedad Saladette cultivar DT-22, se germinaron en semilleros de 54 cavidades en diciembre de 2018, a una edad de 39 días posgerminación, se trasplantaron en invernadero sin ningún tipo de abono. Para su estudio, se realizó un diseño de bloques completos al azar con siete plantas cada uno de acuerdo con la NOM-032-SAG/FITO-2014, esto por cada tratamiento y distribuidos como se muestra en la (Figura 1) con un total de 252 plantas en un invernadero de 500 m2 área, bajo condiciones de temperatura, humedad, y riego controladas (la temperatura se medía con un termómetro digital dentro del invernadero, cuando está llegaba a 40 °C se tomaban medidas correctivas como abrir las alas del invernadero, lo mismo para la humedad y el riego estaba medido por dos tensiómetros que mide el esfuerzo que hacen las raíces para extraer la humedad del suelo, uno estaba a 60 cm y otro a 30 cm del suelo y estos encendían el riego por goteo cuando el de 30 cm rebasaba los 5 kPa (kilopascales) y se apagaba el riego cuando el de 60 cm llegaba a 0 kPa, todo esto era para no generar estrés hídrico en las plantas. Además, se agregó una cepa de Tricoderma sp., X2, proporcionada por el Dr. Sergio Ramírez del INIFAP, Zacatepec, Morelos, con la finalidad de evaluar en conjunto con las cepas de Klebsiellas. Cabe destacar que el T1 es el grupo testigo o control, el T6 un biofertilizante comercial como control positivo.
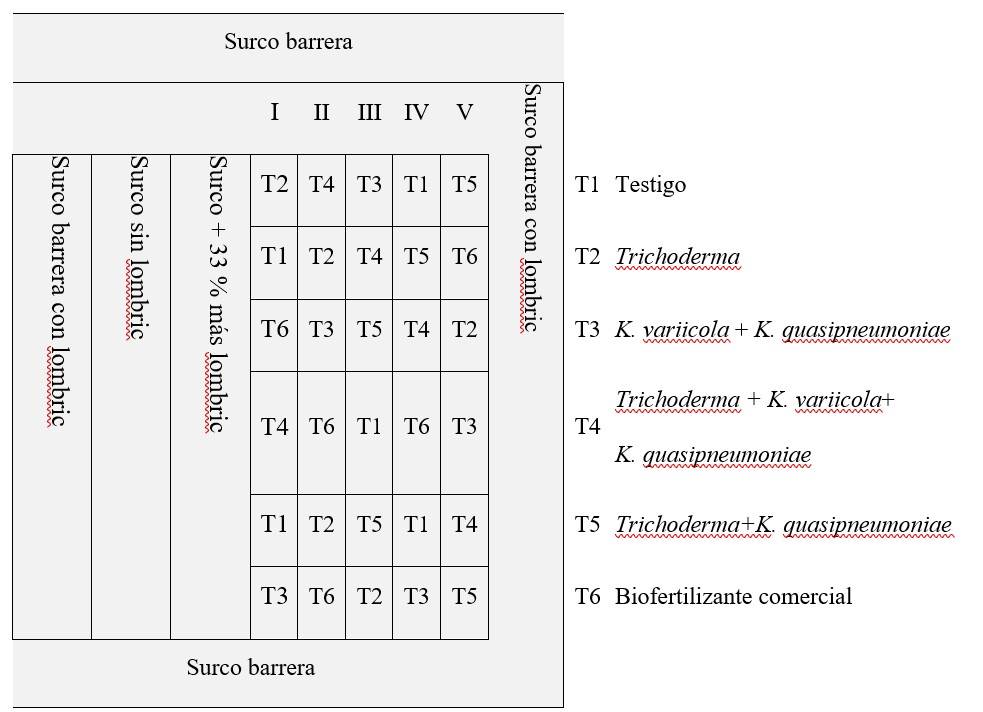
Figura 1 Diseño de bloques completos al azar de las plantas expuestas a los tratamientos microbianos.
Las pruebas de efectividad biológica y translocación a fruto se realizaron durante el ciclo agrícola comprendido de enero-junio de 2019. Las condiciones ambientales fueron monitoreadas durante todo el ciclo hasta finalizar la etapa de producción.
Preparación de los tratamientos microbianos
La selección de las cepas PB02 y HPA43 utilizadas en este trabajo, fue de acuerdo con su previa caracterización como bacterias fijadoras de nitrógeno, solubilizadoras de fosfatos, productoras de ácido indol-3-acético, identificadas molecularmente por secuenciación del gen rpoB como K. variicola y K. quasipneumoniae respectivamente y evaluadas como promotoras de crecimiento radicular in vitro en Solanum lycopersicum (Gutiérrez et al., 2017). Así, también a los resultados obtenidos al ser evaluadas in vivo en otros cultivos de interés comercial por este equipo de trabajo. Se realizó un pre-inóculo de las cepas de manera individual en caldo nutritivo, ajustando una densidad óptica de 1.5x108 microorganismos y el inóculo en solución salina a un volumen final de 1.5 L por cada tratamiento, a la misma concentración de microorganismos.
Inoculación de los tratamientos microbianos en la raíz de S. lycopersicum cultivar DT-22
Posterior al trasplante, se agregaron 50 ml del inóculo con jeringa estéril a nivel de raíz, esto fue a cada una de las plantas por bloque, se realizaron en total 3 inoculaciones por periodos de cada 30 días.
Rendimiento de producción de frutos de S. lycopersicum expuesto a los tratamientos microbianos
Se cuantificó y clasificó el número de frutos producidos por clase en base al peso (g): los de primera clase, pesaron de 100 g o más, segunda clase, de 50 a 99 g y tercera clase con peso menor a 49 g, también se obtuvo el peso absoluto (kg) de la producción por bloque respecto a cada tratamiento; cabe mencionar, que dicha clasificación se realiza en base a la experiencia del productor; Ing. Rubén Ocampo Espín quien ha producido y colocado su producto por más de 15 años en el mercado nacional. Se analizaron como variables de respuesta el peso absoluto de producción, peso por clase (I, II y III). El análisis estadístico se realizó en el programa estadístico SAS versión 9.4. Una prueba de Anova bifactorial, comparación de medias por la prueba de Tukey.
Inocuidad microbiológica de los frutos de S. lycopersicum cultivar DT-22 expuestos a los diversos tratamientos microbianos
Se realizó la recolección de tres frutos al azar por tratamiento en tres eventos distintos, con un total de 54 frutos por invernadero. La recolección y preparación de las muestras se realizó ejecutando las especificaciones de la NOM-109-SSA1-1994. Una vez que los frutos fueron transportados al laboratorio, se procedió a realizar la desinfección de la superficie de los frutos con yodo a una concentración de 0.5% en agua estéril durante 5 minutos, posterior a esto se retiró la cáscara del fruto.
Para el procesamiento de las muestras se tomaron 10 g del tejido interno del fruto y se diluyó en 90 ml de H2O peptonada con NaCl estéril al 0.85%, el procesamiento se realizó en base a las NOM-113-SSA1-1994 y NOM-210-SSA1-2014, de la suspensión resultante se tomaron 100 µl y se inocularon en placas Petri (superficie cultivable 100 mm) en agar Mac Conkey, utilizando la técnica de dispersión con perlas de vidrio en condiciones de esterilidad para el recuento de UFC g como se indica en NOM-110-SSA1-1994. Para la interpretación de resultado se tomaron los valores de referencia establecidos en la NOM-113-SSA1-1994, la cual indica que el límite máximo permisible (LMP) para mesofílicos anaerobios es de 150 000 UFC g-1 y para coliformes fecales de 100 000 UFC g-1 en muestras de alimentos, en conjunto con la ISO 21528-2:2017. Los aislamientos fueron identificados bioquímicamente por el sistema Api20E® Biomerix siguiendo las instrucciones del fabricante.
Determinación de la colonización de raíz de las plantas de S. lycopersicum DT-22
Una vez terminada la etapa de producción se procedió a sacrificar las plantas de S. lycopersicum para obtener la raíz, fueron transportadas al laboratorio para su análisis microbiológico, se realizó la desinfección de la superficie radicular con una solución Tween al 20% durante 10 min (modificado de Bueno et al., 2007). Se tomó 1 cm3 del tejido interno de meristemo apical de la raíz primaria. Se maceró con una varilla de vidrio estéril y se diluyó en 9 ml de agua peptonada con NaCl al 0.85% estéril y se vortexearon a 3 500 rpm durante 1 min. Posteriormente se hicieron diluciones hasta 10-8, en un tiempo no mayor a 20 min, se plaquearon 100 µl de cada dilución en placas de agar Mac Conkey por duplicado, incubando a 25-30 °C por 18 h. Posterior al tiempo de incubación se realizó el recuento de UFC g. Los aislamientos fueron identificados por morfología colonial característica y bioquímicamente.
Evaluación de la translocación por huella genómica de cada aislamiento de Klebsiella
Al total de cepas identificadas como Klebsiella, se les extrajo ADN genómico por choque térmico y se identificaron en base a patrones genéticos por PCR de secuencias consenso intergénicas repetitivas de enterobacterias (ERIC) para evidenciar su origen clonal. Los oligonucleótidos utilizados fueron: ERIC-1 (5’-ATGTAAGCTCCTGGGGATTCAC-3’) y ERIC-2 (5’- AAGTAAGTGACTGGGGTGAGCG-3’), bajo las siguientes condiciones: desnaturalización inicial a 95 °C por 3 min, seguido por 30 ciclos de (93 °C por 45 s, 50 °C por 1 min, 65 °C por 8 min) y una extensión final a 65 °C por 16 min (Versalovic et al., 1991). Los productos del ERIC-PCR se analizaron en geles de agarosa al 1% a 75 V por 2 h y teñidos con bromuro de etidio (3 µl 100 ml-1 de agarosa) y visualizados bajo luz UV (365/302 nm). El patrón genómico de las cepas aisladas del fruto y raíz fueron comparados con las cepas tipo de K. variicola PB02 y K. quasipneumoniae HPA43. Finalmente, la interpretación de los patrones obtenidos se realizó de acuerdo con los criterios de Tenover (Tenover et al., 1995).
Resultados y discusión
Se recolectaron un total de 13 cepas aisladas de la rizosfera de diversos cultivos agrícolas (plátano, maíz, y jitomate) y agua. Las especies de K. variicola, K. pneumoniae y K. quasipneumoniae se diferenciaron molecularmente. Obteniendo 37.5% (3/13) confirmadas como K. pneumoniae las cepas RS10, GN7 y KA2, 6.5% (1/13) K. variicola PB02 y 12.5% (2/13) identificadas como K. quasipneumoniae HPA4-3 y PB06. K. quasipneumoniae y K. variicola han tenido un gran impacto, sin embargo, un mal enfoque, la falta recursos, el desconocimiento de la diversidad entre las especies, la falta de actualización de las bases de datos de los equipos utilizados para la identificación bacteriana en los centros hospitalarios, dificultan su diferenciación y diagnóstico.
Por ello, hay que emplear métodos moleculares para confirmar la identificación y diferenciación de las especies dentro de la rutina microbiológica, dado que estudios de Garza- Ramos et al. (2015) menciona que no es posible diferenciar a K. quasipneumoniae y K. variicola de K. pneumoniae por pruebas bioquímicas y fenotípicas, si no por métodos moleculares (Figura 2).

Figura 1 Diferenciación de las cepas de Klesiella sp. Carril 1: marcador de peso molecular 1kb DNA; carril 2= control positivo; DNA K. variicola, K. quasipneumoniae, K. pneumoniae; carril 3= DNA K. variicola 2006 (control); carril 4= DNA K. quasipneumoniae 10442 (control); carril 5= DNA K. pneumoniae 13883 (control); carril 6= control negativo (agua destilada); carril 7= DNA cepa KC1; carril 8= DNA cepa KC2; carril 9= DNA cepa KC3; carril 10= DNA cepa RS10; carril 11= DNA cepa PB02 (K. variicola); carril 12= DNA cepa HPA4-3 (K. quasipneumoniae); carril 13= DNA cepa PB06 (K. variicola).
Bernabé (2016) reporta a K. quasipneumoniae de la rizósfera de Opuntia ficus identificada por metagenómica dirigida al gen del ARNr16S. Siendo este el primer reporte de K. quasipneumoniae en área ambiental, la cepa HPA4-3 aislada de agua del río azul y la cepa PB06 localizada en la rizosfera de maíz. Cabe mencionar que 43.75% (7/13) de los aislados no amplificaron para ninguna especie, pero si para el género.
Fijación biológica del nitrógeno
Las cepas identificadas como K. variicola PB02 y K. quasipneumoniae HPA4-3 son capaces de fijar nitrógeno atmosférico en 100%, ya que presentan una actividad del complejo nitrogenasa a las 24 h, evaluadas in vitro. Esto último se confirma con el estudio de Fouts et al. (2008) que reporta que K. pneumoniae presenta en su genoma el regulón (KPK-1696-KPK-1715) de la fijación de nitrógeno, indicando que en las cepas se encuentra presente la enzima nitrogenasa la cual cataliza la conversión de N2 a NH4. Carcaño et al. (2006) reportan a Klebsiella sp con una actividad nitrogenasa en un rango de 42-122.46 nmol de etileno por ml donde al comparar con el género Azospirullum sp. (8.62-70.08 nmol) presenta una significativa diferencia en la actividad nitrogenasa de etileno por ml, esto indica que el género Klebsiella sp., cuenta con un gran potencial como bacteria fijadora de nitrógeno.
Solubilización de fosfatos
El 100% de las cepas son solubilizadoras. La de mayor índice fue K. quasipneumoniae PB06 con índices de 10.7 mm, seguida por K. pneumoniae KA-2 con 9.66 mm y K. quasipneumoniae HPA-4-3 con 7.97 mm Tereja et al. (2013) evalúo cepas de Bacillus sp., en donde 29.31% son solubilizadoras de fosfatos, esto pone a Klebsiella como un género con gran potencial para efectuar este fenómeno. La liberación del fósforo soluble es esencial para el desarrollo de las plantas; desempeña un papel fundamental en la síntesis de DNA. Lara et al. (2011), reporta índice de solubilización con rango máximo entre 3 y 4.2 mm de diámetro de un total de 20 cepas entre las que se encuentran P. putida, P. luteola, E. sakasaki y E. cloacae, evaluadas en las mismas condiciones de este documento de trabajo. La diferencia de estos resultados está relacionada con la capacidad de fijar el nitrógeno atmosférico. Se han registrado altas tasas de solubilización en presencia de amonio como fuente de nitrógeno lo cual se relaciona con el porcentaje de fijación en las cepas evaluadas en nuestro trabajo (Beltrán, 2014).
Producción de ácido indol acético
En la producción de AIA se observó que 100% de las cepas son productoras de AIA entre las que destacan: K. pneumoniae RS10 y K. quasipneumoniae PB06 con una elevada concentración de 3.241 y 2.738 µg ml-1 respectivamente que es estadísticamente significativo con respecto al control positivo K. variicola PB con 2.169 µg ml-1. La concentración de AIA en la planta es responsable de incrementar el sistema radicular y follaje de la planta (Taiz-Zeiger, 2013). Además, esta fitohormona se relaciona con la inducción a la elongación de tallos y desarrollo de frutos.
Los resultados obtenidos en este estudio en comparación con los de Bautista-Gallardo (2008) son inferiores. Ellos trabajaron con A. vinelandii y obtuvieron alta producción de AIA. Carcaño et al. (2006) reportan cepas de Azospirillum y Klebsiella con una producción de hasta 26.55 µg ml-1, lo cual confirma que las especies del género Klebsiella son productoras de esta auxina, no obstante en este estudio se pueden comparar que las cepas de Klebsiella sp., que no tienen actividad nitrogenasa presentan una mayor producción de indoles y viceversa, esto confirma los resultados obtenidos dado que en las cepas fijadoras de nitrógeno se obtiene una producción de AIA en bajas concentraciones. Vega et al. (2016) mencionan que las cepas productoras de AIA presentan un mayor efecto promotor de crecimiento, aumentando la elongación de la raíz primaria y el número de raíces secundarias. No obstante, las que no producen AIA retardan la elongación de las raíces.
Produccion de sideroforos
La producción de sideroforos se realizó a las cepas identificadas molecularmente: el 100% de las cepas evaluadas son productoras de sideroforos que concuerda con lo mencionado por Arena et al. (2015) donde demuestra que K. pneumoniae produce sideróforos: enterobactina y aerobactina. Esto le confiere ventajas competitivas en la colonizacion de la raiz, brindando protección mediante el biocontrol que establece, debido a la captacion del hierro por los sideroforos, siendo atraidos a la membrana bacteriana limitando asi la disponibilidad del hierro para microorganismos que carezcan del sistema de asimilacion. Esto es concordante con lo mencionado por Molina et al. (2015) en relación al antagosnismo entre B. altitudinis y T. cucumeris.
K. quasipneumoniae HPA4-3 y PB06 y K. variicola PB02 las seleccionamos como candidatas en conjunto con el Tricoderma sp para ser inoculadas en S. lycopersicum Saladette cultivar DT-22, aunque la cepa de K. pneumoniae RS10 presenta varías características de promoción vegetal, se descartó por diversos estudios publicados donde demuestran las alteraciones en la salud humana.
Rendimiento de producción en Saladette cultivar DT-22
Se hicieron 11 cosechas (una por semana) en total se tomaron al azar de cinco cosechas en eventos distintos para analizar el rendimiento. La cosecha total de producción estimada fue de: 1 392 kg por todos los tratamientos. Los resultados obtenidos muestran que los tratamientos evaluados, no presentan el mismo efecto en la producción de jitomates de primera clase, el análisis comparativo Anova nos da un valor de p= 0.0008*, los datos completos se observan en la (Figura 3) donde se clasifica de acuerdo con el agrupamiento de Tukey, tratamientos con la misma letra no presentan diferencias significativas en su efecto, el T4 tiene el valor promedio más alto (p= 0.0007*) en producción (39.618 kg) por cosecha, respecto grupo testigo (T1). A diferencia de los tratamientos donde fueron evaluados de manera individual, T2 que no presenta diferencia significativa respecto al testigo (p= 0.464), el T3 tiene una menor producción (p= 0.999 * ) en comparación con los demás tratamientos y el testigo (T1), por otro lado, en el T5, no presenta diferencia significativa (p= 0.999) respecto al grupo testigo (T1), este último consorcio se realizó por que la cepa HPA43 fue capaz de promover el crecimiento radicular in vitro de S. lycopersicum L. (Gutiérrez et al., 2017).

Figura 3 Gráfico de comparación de medias del rendimiento de la producción (kg) jitomate Saladette cultivar DT-22. En las gráficas se muestra la barra del error estándar y la clasificación de acuerdo con el agrupamiento de Tukey que indica que los tratamientos con las mismas letras no presentan diferencias significativas.
Además de los consorcios microbianos se evaluó un biofertilizante comercial T6, ya estandarizado y se comprobó que, sí aumenta el rendimiento de producción de los cultivos respecto al T1; sin embargo, no es significativo en comparación con nuestro T4. Estos microorganismos son capaces de establecer una asociación mutualista (bacteria-hongo-planta) y potencializar el rendimiento de la producción, estimulan el desarrollo del sistema radicular de la planta influyendo en la diferenciación celular a nivel del meristemo, internalizándose a la zona de diferenciación e influyen en la diferenciación de las células de acuerdo a las necesidades de la planta y las condiciones ambientales, por lo tanto hay mayor número de pelos radiculares los cuales permiten el ingreso de nutrientes y minerales al sistema vascular de la planta (Scharf et al., 2016; Calzavara et al., 2018).
Análisis microbiológico de los frutos expuestos a los tratamientos microbianos
En el Cuadro 1 se muestra el recuento microbiológico de los frutos por tratamiento, se observan las medias y desviaciones estándar del Log de UFC g-1 de tres repeticiones en tres eventos distintos, se reporta el número de microorganismos g-1 presentes en la muestra, como la ISO 21528-2:2017, los valores obtenidos del recuento están en los límites máximos permisibles.
Cuadro 1 Recuento microbiológico en frutos de S. lycopersicum DT-22 en cada tratamiento.
| Tratamiento | Recuento microbiológico Log de UFC/g (X ±DS) | N (microorganismos g-1) | (Pr >F) | Referencia |
| T3 | 2.6 ±2.5 | 3262 a | p= 0.469 | NOM-113-SSA1-1994, ISO 4833-1:2013, ISO 21528-2:2017 |
| T4 | 2.2 ±2.1 | 1595 a | ||
| T5 | 2.5 ±2.5 | 2828 a | ||
| T1 | 2.2 ±2 | 1545 a |
X= media; DS= desviación estándar; N= número; T3= K. variicola y K. quasipneumoniae; T4= K. variicola, K. quasipneumoniae, Trichoderma sp,; T5= K. quasipneumoniae, Trichoderma sp.; T1= testigo. El subíndice a en cada valor, pertenece al agrupamiento de Tukey, letras iguales indican que no existe diferencia estadística entre los tratamientos.
Por el recuento de UFC g-1 encontrada se infiere que se trata de la microflora endémica que está asociada a los sitios anatómicos del fruto, que tienen afinidad por sitios específicos de la planta de acuerdo con la composición nutricional de cada sitio anatómico (Ottesen et al., 2013). Es de vital importancia la vigilancia microbiológica y el estudio de la ecología microbiana de estos cultivos de interés comercial, como el jitomate DT-22 que se comercializa directamente al mercado y es altamente consumido en fresco, debido que los desinfectantes utilizados son de uso superficial en el fruto, no ejercen un efecto sobre los tejidos internos (Barak et al., 2012). Debemos vigilar que los frutos no representen un riesgo para la salud del consumidor.
Análisis molecular de la huella genómica de K. variicola PB02 y K. quasipneumoniae HPA43
Se obtuvieron en total 114 cepas (12 de frutos y 102 raíz), identificados fenotípica y bioquímicamente como Klebsiella sp. Los perfiles obtenidos en base al ERIC-PCR permitió clasificar clonas (38) y subclonas (16) de las cepas de K. variicola PB02 y K. quasipneumoniae HPA43, todas a partir de raíz, de las cepas aisladas de frutos no se determinó ningún aislamiento con el mismo patrón de bandas similar al de las cepas control, como se observa en la (Figura 4).

Figura 4 Se observan los patrones de clonalidad de las secuencias intergénicas repetitivas de enterobacterias. a) M= marcador de peso molecular de 1Kb; 1= cepa K. variicola PB02; 2= K. quasipneumoniae HPA43; 3= control negativo. Cepas de frutos; 4= T5R3, 5= S2, 6= R4, 7= V4, 8= P3, 9= Z3, 10= T3R2, 11= R3, 12= N3. Cepas de raiz; 13= T5R5, 14= T5R1, 15= T3R4: b) M= marcador de peso molecular de 1Kb; 1= 3R1, 2= 3R2, 3= 3R4, 4= 3R4, 5= 3R5, 6= 5R1, 7= 5R2, 8= 5R3, 9= 5R4, 10= 5R5, 11= T5R3, 12= T5R4, 13= T3R2, 14= V5, 15= W5, 16= T5, 17= T3R1, 18= T3R4, 19= T3R5, 20= T3R, 21= 5R1, 22= A1, 23= A6, 24= B6, 25= C6, 26= D6, 27= E6.
Este método se ha utilizado principalmente para la determinar brotes hospitalarios con el objetivo de comprobar si los aislamientos similares fenotípicamente tienen una relación genética, dado a la alta tasa de variabilidad genética en procariontes ocasionada por factores ambientales, se amplifican secuencias intergénicas conservadas en enterobacterias que permitan identificar por medio de su huella digital que se derivan de un ancestro en común, en un mismo sitio y en un periodo definido (Tenover et al., 1995). Estos perfiles son útiles para analizar la propagación y distribución de las bacterias en el ambiente, como es que se mueven de un lugar a otro y las condiciones a las cuales persisten.
En los resultados obtenidos, se encontró similitud genética entre las cepas Klebsiella aisladas de raíz y las cepas K. variicola PB02, K. quasipneumoniae HPA43 inoculadas; no obstante, hacen faltaestudios moleculares especificos que permitan confirmar que estas bacterias u otras BPCV no traslocan a los frutos, que no han intregado mecanismos de adaptacion, por transferencia horizontal que permitan la traslocación al fruto como es el caso de E. coli, en respuesta a los factores ambientales, geneticos o ambientales (Ottesen et al., 2013). Para comprender mejor la interaccion de la ecología microbiana en el fruto y determinar si la colonización de estas bacterias es un factor determinante para desarrollar un problema de salud a los consumidores.
Conclusiones
El uso de bacterias promotoras de crecimiento vegetal en consorcios con hongos antagonistas potencializa el rendimiento de producción de cultivos agrícolas de interés comercial ayudando a satisfacer las demandas de consumo y reduciendo los costos de producción en un tiempo menor en comparación con la agricultura convencional. Se requieren más estudios sobre los efectos de la cantidad de inoculo usado en el rendimiento de los frutos y usar herramientas moleculares más finos de las cepas aisladas de los frutos para corrobora en el patrón clonal, y proponerlos con mayor certeza para el estudio de translocación de BPCV desde la rizosfera a los frutos.

