










Services on Demand
Journal

Article
 text in
text in  English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
 Cited by SciELO
Cited by SciELO Related links
 Similars in
SciELO
Similars in
SciELO Share

 Permalink
PermalinkRevista mexicana de ciencias agrícolas
Print version ISSN 2007-0934
Rev. Mex. Cienc. Agríc vol.10 spe 22 Texcoco Mar./Apr. 2019
https://doi.org/10.29312/remexca.v0i22.1856
Articles
Birds in agricultural systems with conservation tillage in northern Mexico
1Programa de Innovación en el Manejo de Recursos Naturales-Campus San Luis Potosí-Colegio de Postgraduados. Iturbide 73, Salinas de Hidalgo, San Luis Potosí, México. CP. 78600. (benjamin@colpos.mx; josep@colpos.mx).
2 Departamento de Suelos-Universidad Autónoma Chapingo. Carretera México-Texcoco km 38.5, Chapingo, Estado de México. CP. 56230. (biologo-ugalde@hotmail.com).
3 Programa de Desarrollo Rural-Campus Montecillo-Colegio de Postgraduados. Carretera México-Texcoco km 36.5, Texcoco, Estado de México. CP. 56230. (figueroa@colpos.mx).
4 Programa de Innovación Agroalimentaria Sustentable-Campus Córdoba-Colegio de Postgraduados. Carretera Federal Córdoba-Veracruz km 348, Congregación Manuel León, Amatlán de los Reyes, Veracruz, México. CP. 94946. (f-katia@hotmail.com).
In Mexico, the creation of new agricultural areas causes the loss of bird species. To know the role of farming systems with zero conservation tillage-live fence (ZCT-LF) to maintain diversity-habitat use by birds, during June-September 2011, a bird monitoring was conducted in central Mexico. Species richness was analyzed with Jacknife1, the similarity with Sorensen, Conglomerates; abundance with log-normal model, χ2; the diversity with Shannon-Wiener; and possible differences in these parameters with Kruskal-Wallis. Habitat use in the vertical (substrates)-horizontal planes (layers) was inferred with the frequency of observation (FO) and Poisson regression (ARP). The average results of Jacknife1 for UE’s were 38.7%, Sorensen 31%, a cluster with three subamalgamations (minimum, e= 2.7, maximum, e= 3.6) was formed; Lognormal (χ2= 130.09, Y= 0.3518, 3 dof), χ2 (P-value = 0.028); Shannon-Wiener H’= 2.99; Kruskal-Wallis P-value = 0.0248, 0.028, 0.4232, respectively. The FO suggest that the higher layer-substrate was most used (46.15%) - flight (27.95%); The ARP for layer, behavior, sex, age, substrate suggests that 2, 4, 1, 2 and 6 variables had significant coefficients. Mexico faces the problem of opening land to agriculture, arid and semiarid areas do not escape this phenomenon; therefore, the ZCT-LF systems are an option to maintain and preserve the avifauna.
Keywords: abundance; behavior; habitat
En México, la apertura de nuevas zonas agrícolas propicia la pérdida de especies de aves. Para conocer el rol de sistemas agrícolas con labranza de conservación cero-cerco vivo (LCC-CV) en el mantenimiento de la diversidad-uso de hábitat por la avifauna, durante junio-septiembre de 2014, se realizó un monitoreo de aves en el centro-norte de México. La riqueza de especies se analizó con Jacknife1, la similitud con Sorensen, Conglomerados, la abundancia con un modelo Log-normal, χ2, la diversidad con Shannon-Wiener y las posibles diferencias en dichos parámetros con Kruskal-Wallis. El uso de hábitat en los planos vertical (estratos)-horizontal (sustratos) se infirió con la frecuencia de observación (FO) y regresión Poisson (ARP). En los sistemas (LCC-CV) se registraron 52 especies de aves, distribuidas en cinco órdenes, 19 familias y 10 subfamilias. Los resultados promedio de Jacknife1 para UE’s fue de 38.7%, Sorensen 31%; asimismo, se conformó un Clúster con tres subamalgamaciones (mínima, e= 2.7; máxima, e= 3.6); Log-normal (χ2= 130.09, Y= 0.3518, 3 gdl), χ2 (p-value= 0.028); Shannon-Wiener H’= 2.99; Kruskal-Wallis de p-value= 0.0248, 0.028, 0.4232, respectivamente. Las FO sugieren que el estrato-sustrato más utilizado fue el superior (46.15%)-vuelo (27.95%). El ARP para estrato, comportamiento, sexo, edad, sustrato indicó que 2, 4, 1, 2 y 6 variables presentaron coeficientes significativos. México enfrenta el problema de apertura de tierras a la agricultura, las zonas áridas y semiáridas no escapan a este fenómeno, por ello, los sistemas de LCC-CV constituyen una opción para mantener y conservar la avifauna.
Palabras clave: abundancia; comportamiento; hábitat
Introduction
Mexico is one of the 12 megadiverse countries, its avifauna is made up of 1054 species (22 orders and 78 families), equivalent to 11% worldwide (Navarro and Benítez, 1993). Its ornithofauna is associated with its forest cover, which covered (70’s), 72.05% of the national territory. This coverage has been subject to disturbances and disturbances (Masera, 2001). During 1970-1980, agricultural areas covered 16% of the territory, of which 4.58% were irrigated, 11% were seasonal forest crops, of which 93.5% were less than 20 ha (Navarro et al., 2000).
The opening of new agricultural areas has led to the loss of some species, particularly birds (Ramírez-Albores, 2010). Those of arid-semiarid environments do not escape these alterations due to their fragility and wide extension of 50-60% of the national territory (Pompa-García, 2008) of their xerophytic scrub, an important portion of this is distributed in the Mexican Altiplano (Rzedowski, 1978). The shrublands of the Potosino-Zacatecano highlands have been used to establish grazing areas and conventional agricultural crops themselves that have been managed under a scheme of intensive tillage with supply of inputs to increase soil fertility-crop yield.
This management has caused compaction, erosion, soil-water contamination; caused the loss of biodiversity (Figueroa-Sandoval, 1983), particularly of the avifauna, a taxonomic group that plays a transcendental role in the energy flows within and between ecosystems since they sustain diverse trophic chains, acting on the vegetative propagation as vectors of dissemination of seeds, some are in risk category (Herrera, 1985). In order to mitigate soil deterioration and increase the yield of traditional crops-environmental quality, mixed agricultural systems based on zero conservation tillage-live fence (ZCT-LF) technology have been established in the region (Navarro et al., 2008), which seem to mitigate the negative effects of agricultural intensification.
In order to plan the compatibility of productive systems with the conservation of biodiversity, it is essential to evaluate the changes they cause to their populations (Lantschner and Rusch, 2007). However, the importance (border effect) of these systems of mixed agricultural production and their benefit, in terms of conservation, for wildlife species, has not been studied, much less for the birdlife that makes use of them in this type of systems surrounding forests xerophytic. The objective of this research was to determine the role played by agricultural systems with LCC-CV on diversity and habitat use (vertical-horizontal) by bird communities in Central Mexico.
Materials and methods
The agricultural area of the Municipality of Salinas de Hidalgo, San Luis Potosí, is located in the Potosino-Zacatecano Altiplano, located in the physiographic region of the Central Plain of the Mexican Altiplano, at the southern end of the Chihuahuan Desert within the highlands of the center and south of San Luis Potosi and southeast of Zacatecas, in the coordinates 22° 45’, 22° 42’ North and 01° 57’, 101° 46’ West, between the heights of the 2 100 to 2 330 masl. The area borders North with Santo Domingo, Villa de Ramos, South with Zacatecas, West with Villa de Ramos, Zacatecas, East with Charcas, Venado, Moctezuma, Zacatecas (Luna-Cavazos et al., 2007).
Sampling design
Seven study units (UE’s) were chosen, the polygonal and surfaces of each of them (9, 10, 4, 2, 3, 10 and 8 ha, respectively) were delimited and quantified with a Geographic Information System (GIS) (Figure 1). The sampling design used was systematic sampling (MS) at convenience with predetermined distances of 250 m between each unit of choice (UEL) (Cochran, 1977).

Birds were monitored using a mixed counting scheme at points with fixed radius of 25 m and intensive search (Ralph et al., 1996). This combined system allows a better detection of some species of quiet, silent and conspicuous behavior (López-DeCasenave et al., 1998). The birds were observed with binoculars of 25 X 50 m (Bushnell) and the sighted species were identified with standard field guides (birds from Mexico and North America). The data referring to the arboreal vegetation were collected through the application of quadrants with a central point (Cottam and Curtis, 1956), for herbaceous-shrubbery embedded paintings (Oosting, 1956). Likewise, the habitat variables (land cover) were registered through the Canfield Line (Canfield, 1941). The samplings were carried out monthly (June-September 2011).
Statistic analysis
The specific wealth of birds was systematically ordered by the classification of American Ornithologists Union (AOU, 2009) for this purpose, information was used on the number of species recorded in a timely manner in time and space. Species richness was analyzed using Jacknife1 using the presence-absence data recorded in each UE-pulse, sampling, Magurran (1988). The similarity between the UE’s evaluated was estimated with Sorensen (Chao et al., 2005), these parameters were calculated in EstimateS 8.2.0. (Burnham and Overton, 1979; Colwell, 2009). To graphically visualize the similarity in the wealth recorded in the UE’s under study, a multivariate analysis of clusters by observations was carried out (Kaufman and Rousseeuw, 2009), which was carried out in Minitab 15.1.20 (2007).
The distribution of abundance (number of individuals per species) registered in each UE was interpreted ecologically through Log-normal models, as these were the ones that better adjusted the distribution of the data of the present work (Preston, 1948), said models were carried out in Microsoft Excel (2003). To determine if the number of individuals registered per species is potentially distributed in each UE, a goodness-of-fit test of (2 was used for contingency tables for independent samples (Miller, 2003b). This analysis was carried out in JMP IN version 8.0.2. (2009).
The diversity of bird species in each UE was analyzed using Shannon-Wiener using information on the frequencies of the species recorded in each of them (Tramer, 1969). Said estimator was calculated in EstimateS 8.2.0. The possible differences in wealth, abundance and diversity among UE’s were determined by Kruskal-Wallis (Miller, 2003a), which were carried out in JMP IN 8.0.2 (2009).
The proportion of individuals per species that make use of each vertical layer and substrate (horizontal plane) was estimated with observation frequency indexes (FO); Curts (1993) modified for the present. These were calculated in Microsoft Excel (2003). To determine the possible relationship between recorded bird abundance (dependent variable ‘y’) and vertical strata of vegetation; behaviors exhibited by birds (based on the criteria proposed by Remsen and Robinson, 1990), sexes ages and horizontal substrates (each of them as independent variables, ‘x’), Poisson regression models (ARP) were used. The adjustment of the dependent variable to the independent variables was carried out using generalized linear models (GLM); McCullagh and Nelder (1989). In all the ARPs, the criterion of selection of variables by backward steps (Stepwise) was used, with criterion of classification of the minimum Akaike (AIC); Akaike (1969), these analyzes were performed in R-version 2.12.1 (2010).
In all cases an α= 0.05 was used, so that those in which p< 0.05 were considered as statistically significant coefficients.
Results and discussion
Based on the AOU (2009), 52 bird species were registered in the systems (ZCT-LF), distributed in five orders, 19 families and 10 subfamilies. The most representative orders in number of species were passerine with 35 (67%), falconiform with seven (13%), columbiform with five (10%), piciform with four (8%). Of the species recorded in this study, one species is in danger of extinction (P), two threatened (A), five subject to special protection (Pr), 44 not included in any category; only four are endemic (SEMARNAT, 2010). The Jacknife1 estimator showed that with the sampling effort by UE’s under study it was possible to detect 100% (UE1), 46.7 (UE2), 23.26 (UE3), 25 (UE4), 30.3 (UE5), 19.9 (UE6) and 26% (EU7) of the bird species that are theoretically present in each EU (Figure 2).
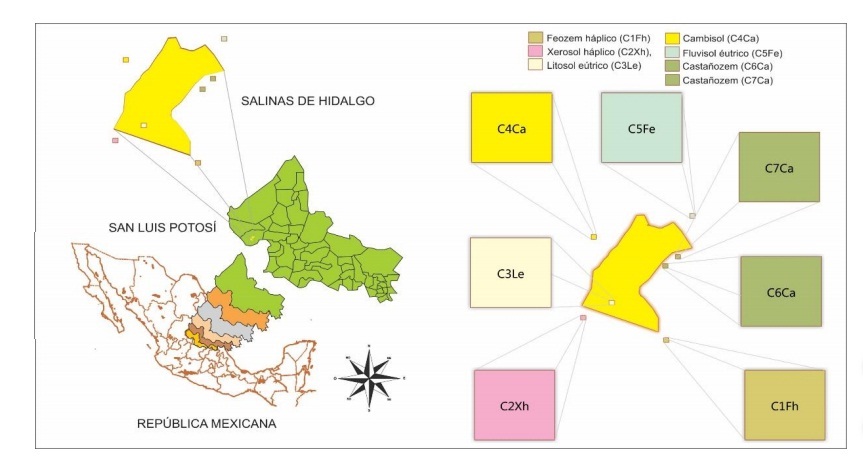
Figure 2 Observed and estimated bird species accumulation curves (Jackknife index1) for each UE in agricultural systems of Salinas de Hidalgo, San Luis Potosí, Mexico.
Likewise, in the case of samplings (pulses) Jacknife1 managed to detect 100% (P1), 41.6 (P2), 39 (P3), 30.4% (P4) in each of them (Figure 3). The Kruskal-Wallis results (p-value= 0.0248) identified significant differences in the number of species registered in each of the UE’s under study.
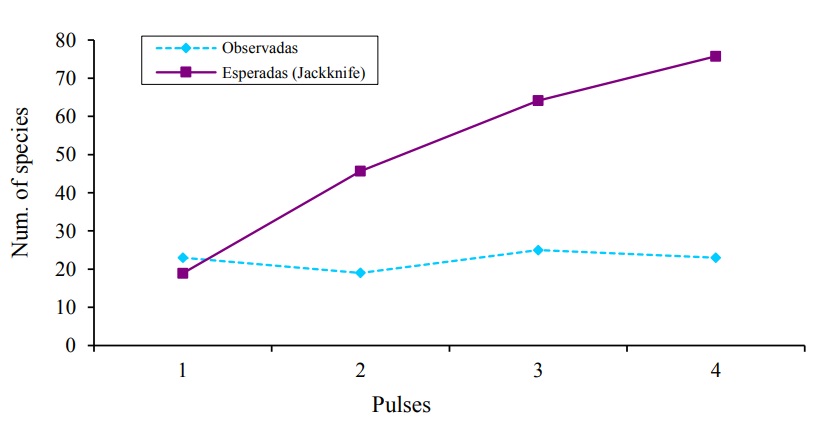
Figure 3 Observed and expected bird species accumulation curves (Jackknife index1) for each pulse in agricultural systems of Salinas de Hidalgo, San Luis Potosí, Mexico.
According to Sorensen, the average similarity of species registered among (UE’s), expressed as a percentage, was 31%. The highest similarity was presented in the UE’s 1-7, with 50%, with 11 shared species. Similarly, the lowest similarity was observed between the UE’s 2-4 and 2-6, with 15% among them, seven and six shared species, respectively. The Cluster analysis for the data of species registered by UE exhibited a single cluster; however, three sub-amalgamations were formed with a minimum euclidean distance of e= 2.7 and maximum e= 3.6 (Figure 4).
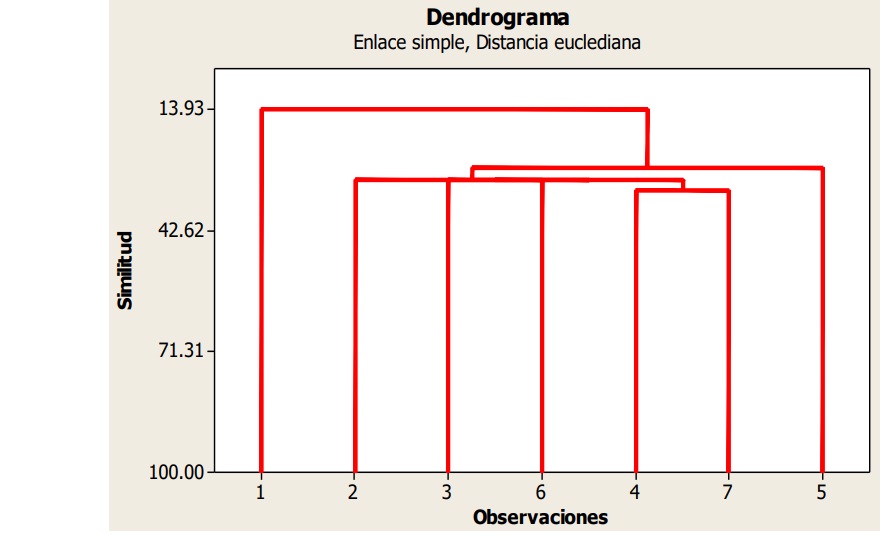
Figure 4 Cluster of similarity for species by UE’s in agricultural systems of Salinas de Hidalgo, San Luis Potosí, Mexico.
The Log-normal model indicates that the logarithms of the bird frequency data recorded by UEs are distributed in a normal way (χ2= 130.09, Y= 0.3518, 3 dof), which shows an adjustment in the distribution of abundances to said series (Figure 5). The results of χ2 show that the highest proportion of individuals per bird species was recorded that could potentially be expected in the UE’s under study, which is supported by the Kruskal-Wallis analysis (p-value= 0.028), which detected differences in abundances per species recorded in each of them.
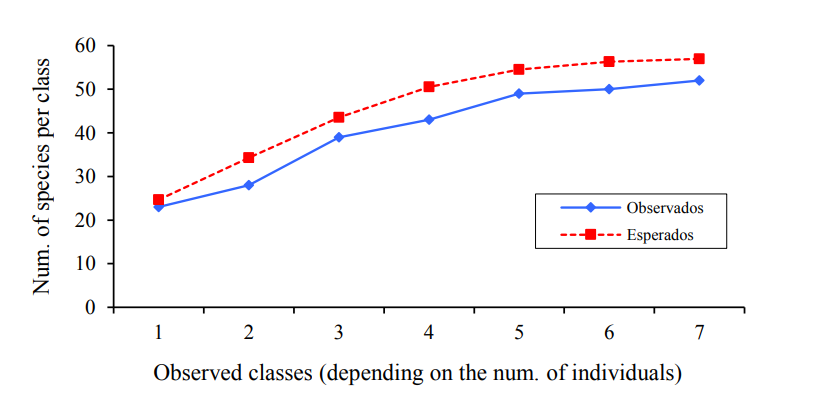
Figure 5 Distribution of abundances of birds registered in the UE's under study adjusted to a log-normal series in agricultural systems of Salinas de Hidalgo, San Luis Potosí, Mexico.
Shannon-Wiener considering the number of individuals per species recorded in each UE’s, estimated a minimum value of H’= 2.84 (UE1) and maximum of H’= 3.15 (UE7) (Figure 6). While Kruskal-Wallis comparing the diversity of species did not detect significant differences (p-value= 0.4232) between the species diversity recorded and estimated in each of the UE’s under study.

Figure 6 Shannon-Wiener index for each UE’s in agricultural systems of Salinas de Hidalgo, San Luis Potosí, Mexico.
The analysis of FO of birds recorded in each of the strata (heights) of the vegetation; showed that birds prefer, in that order, to the upper (46.15%), low (37.31%) and medium (16.54%) vegetation stratum. For its part, the analysis of FO of birds recorded in each of the substrates (horizontal plane), indicated a preference for flight (27.95%) using soil substrate (24.58%), herbaceous (15.15%), fence live (nopal) (7.07%), pirul (Shinus spp., 6.4%), huizache (Acacia spp., 5.72%), mesquite (Prosopis spp., 5.39%), maguey (Agave spp., 3.03%), cassava (Yucca spp. 2.69%), nopal (Opuntia spp. 2.02%). The Poisson regression between the abundance of birds and the vertical strata of the vegetation, behaviors, sexes, ages and horizontal substrates, showed that variables 2, 4, 1, 2 and 6 had an effect on the dependent variable (y) with values of AIC of 58.192, 52.373, 53.574, 54.272 and 55.888, for each of the analyzes, respectively (Table 1).
Table 1 ARP results using GLM models for bird abundance and vertical strata, behaviors, sexes, ages and horizontal substrates in the UE’s under study in agricultural systems of Salinas de Hidalgo, San Luis Potosí, Mexico.
| ARP | Estimate | Std. error | z value | Pr(>|z|) | Significance |
| ARP abundance of birds and vertical layers of vegetation | |||||
| Low stratum | 0.059316 | 0.011019 | 5.383 | 7.33E-08 | *** |
| Upper stratum | 0.01498 | 0.00526 | 2.848 | 0.0044 | ** |
| ARP abundance of birds and their behavior | |||||
| Pairing | 1.03963 | 0.14044 | 7.403 | 1.33E-13 | *** |
| Courting | -0.85504 | 0.2494 | -3.428 | 0.000607 | *** |
| Courting 2 | 0.54178 | 0.11861 | 4.568 | 4.93E-06 | *** |
| Hanger | 0.08689 | 0.01027 | 8.464 | < 2E-16 | *** |
| ARP of abundance of birds and their sex | |||||
| H | 0.10231 | 0.02614 | 3.914 | 9.06E-05 | *** |
| ARP abundance of birds and their ages | |||||
| A | 0.017296 | 0.004362 | 3.965 | 7.33E-05 | *** |
| J | 0.062126 | 0.01277 | 4.865 | 1.14E-06 | *** |
| ARP of bird abundance and horizontal substrates | |||||
| Soil | 1.005138 | 0.400328 | 2.511 | 0.01205 | * |
| Soil 2 | -0.047395 | 0.020329 | -2.331 | 0.01974 | * |
| Suscerc | 0.626164 | 0.25402 | 2.465 | 0.0137 | * |
| Suscerc 2 | -0.065653 | 0.021738 | -3.02 | 0.00253 | ** |
| Susther | -0.236201 | 0.111782 | -2.113 | 0.0346 | * |
| Susther 2 | 0.019102 | 0.006991 | 2.732 | 0.00629 | ** |
In all cases a Poisson distribution was assumed in the abundance (y), so a logarithm was applied as a function of the link between it and the independent variables (X)= H= female; A= adult; J= juvenile; Soil 2= quadratic soil effect; Suscerc= fencing substrate; Suscerc 2= quadratic effect of Suscer; Susther= herbaceous substrate; Susther 2= quadratic effect of Susthe
The trends of the specific richness and conservation status of some of the registered birds coincide partially with that reported by Cárdenas et al. (2003) who conducted a study in agricultural areas finding that the most representative family in number of species and frequencies of these was Tyrannidae; meanwhile, Ibarra et al. (2001) conducted an investigation on the avifauna associated with two cacao plantations in Tabasco (Mexico), concluding that the most represented families were Emberizidae and Tyrannidae.
These results are very similar to those obtained in the present study; however, the inferences obtained in the estimator of species richness contrast with those obtained by Bolwig et al. (2004) who report avifaunal species richness data for three types of agricultural ecosystems in Uganda, where the average intensity of crop shares the anthropic disturbance characteristic of agricultural activity, in our UE’s; nevertheless, the patterns in the rarefaction curves allowed to affirm that the ZCT-LF presupposes a lower pressure.
Thus, the trends in species richness, for the present, seem to respond to the sampling effort made and the bird's own migration phenology, which can vary even from one year to another, which may affect its detectability, as suggested by Bojorges and López-Mata (2005) who, despite having good approximations (90%) in the estimation of this parameter, recommend increasing the monitoring effort on a spatial-temporal scale, considering the environmental co-operations that may affect in the presence of accidental or migratory birds (Carrascal, 1987).
On the other hand, Rappole et al. (1993) report that in agricultural systems and in some natural systems, the increase of species during the migratory period and the displacement of resident species is minimal, since migratory birds can be flexible in the use of the habitat, being able to find them in disturbed systems as is was the case with this study. However, the patterns of wealth registered between UE’s contrast with that reported by Sánchez and López (1988) who point out that two faunal communities are similar if their similarity index is less than 66.6%, this is for different types of traditional agricultural type plots; however, the composition and structure of the bird community recorded in this research was more heterogeneous.
The distribution of abundance recorded in the present evidence is an avifauna community made up of few but abundant species, followed by more or less abundant species and rare species, without the presence of dominance (monopoly) over the use of resources. These results differ with that reported by Ugalde-Lezama et al. (2009) suggest that in semiconserved areas of a forest stand of Pinus greggii Englem, in Zoquiapan, Mexico, the distribution of abundances was adjusted to a Log-normal series; however, the trends recorded in this one showed that the ZCT-LF systems support bird communities similar to the disturbance gradient that these forests exhibited.
Similarly, the proportions of individuals registered by species in each UE could be attributed to their physiognomy, which provides a wide range of resources that allow the spatial distribution of bird species registered in them. Similar patterns are reported by Enríquez-Lenis et al. (2007) who point out that seasonality can modify positively or negatively the structure and floristic composition, which condition the use of ecological niches in the vertical-horizontal plane, determining the patterns of abundance and diversity, particularly in birds.
The diversity of species recorded in this research was relatively high, especially considering that the UE’s under study are agricultural systems with ZCT-LF. However, this contrasts with that reported by Carrascal et al. (2002) who estimated low diversity patterns in different disturbance gradients, including forest stands and agricultural areas.
The frequencies of birds that use or have preference for a vertical stratum in particular, contrast with that reported by Emlen (1977) who recorded homogeneous patterns of use from the lower stratum to the upper stratum. On the other hand, Carrascal and Telleria (1985) report the use of the middle and lower strata, noting that the plant substrate is more relevant to birds compared to the variable height; however, the birds recorded in this work did separate vertically, which coincides with Cueto and López-DeCasenave (1999) who reported an effect of the vertical structure of the vegetation on the abundances-distribution of this taxonomic group, particularly in the herbaceous and arboreal strata.
On the other hand, the frequencies of birds that use or have preference for some substrate in the horizontal plane coincide with that reported by Becerra and Grigera (2005) who point out that these have an effect on diverse patterns of bird activity such as feeding, so the variation in their availability and productivity determines the structure and composition of bird communities. The birds in this study had live fences, coupled with the presence of trees and shrubs, which had an effect on their abundances, which provided shore habitats between two or more different environments as suggested by Salinas et al. (2007) who point out that the increase in diversity is largely due to the presence and establishment of live fences, since they function as connectivity corridors in the conservation of wildlife by offering spaces for perches, feeding, shelter, nesting for some species; particularly birds, showing that they can harbor, sometimes very particular, more species, even than traditional agricultural areas.
The ZCT-LF systems are beneficial for avifaunal biodiversity in the parameters evaluated between UE’s, which were acceptable; however, it is recommended to increase the monitoring effort on a spatio-temporal scale that complements the inferences made. There was effect of the strata (vertical) and substrates (horizontal), finding differential patterns of use of these. This study, is the first of its kind, which shows that these agricultural systems (ZCT-LF) are convenient in the maintenance-conservation of birds in the Potosino-Zacatecano Altiplano, for the multiple advantages and services offered to this taxonomic group compared to traditional agricultural systems.
Conclusions
In this study, 52 bird species of five orders, 19 families and 10 subfamilies were recorded. The averages of Jacknife1 for UE’s were 38.7% and Sorensen 31%.
A Cluster was formed with three sub-amalgamations (minimum, e= 2.7, maximum, e= 3.6). The frequencies of observation suggest that the most used stratum-substrate was the higher one (46.15%) - flight (27.95%). The ZCT-LF systems are an option to maintain and conserve birdlife in northern central Mexico.
Acknowledgments
To the owners of the agricultural systems of Salinas de Hidalgo, San Luis Potosí.
REFERENCES
AOU. 2009. Check-list of North American birds. 7th (Ed.). American Ornithologists Union, Washington, DC. 131 pp. [ Links ]
Akaike, H. 1969. Fitting autoregressive models for prediction. Annals of the Institute of Statistical. Mathematics. 21(1):243-247. [ Links ]
Becerra, S. R. M. y Grigera, D. 2005. Dinámica estacional del ensamblaje de aves de un bosque Norpatagónico de Lenga (Nothofagus pumilio) y su relación con la disponibilidad de sustratos de alimentación. El Hornero. 20(2):131-139. [ Links ]
Bojorges, B. J. C. y López, M. L. 2005. Riqueza y diversidad de especies de aves en una selva mediana subperennifolia en el Centro de Veracruz, México. Acta Zoológica Mexicana (ns). 21(1):1-20. [ Links ]
Bolwig, S.; Mushabe, D.; Nkuutu, D.; Pomeroy, D. and Tushabe, H. 2004. Biodiversity in Uganda’s Farming Systems in Relation to Agricultural Intensification. Makerere University Institute of Environment and Natural Resources (MUIENR) PO. Box 7298, Kampala, Uganda. 105 p. [ Links ]
Burnham, K. P. and Overton, W. S. 1979. Robust estimation of population size when capture probabilities vary among animals. J. Ecol. 60(5):927-936. [ Links ]
Canfield, R. H. 1941. Application of the line interception method in sampling range vegetation. J. Forest. 39(4):388-394. [ Links ]
Cárdenas, G.; Harvey, C. Ibrahim, M. y Finegan, B. 2003. Diversidad y riqueza de aves en diferentes hábitats en un paisaje fragmentado en Cañas, Costa Rica. Agroforestería en las Américas. 10:(39-40):78-85. [ Links ]
Carrascal, L. M. 1987. Relación entre avifauna y estructura de la vegetación en las repoblaciones de coníferas de Tenerife (Islas Canarias). Ardeola. 34(2):193-224. [ Links ]
Carrascal, L. M. y Tellería, J. L. 1985. Estudio multidimensional del uso de espacio en un grupo de aves insectívoras forestales durante el invierno. Ardeola. 32(1):95-113. [ Links ]
Carrascal, L. M.; Palomino D. y Lobo, J. M. 2002. Patrones de preferencias de hábitat y de distribución y abundancia invernal de aves en el centro de España. Análisis y predicción del efecto de factores ecológicos. Animal Biodiversity and Conservation. 25(1):7-40. [ Links ]
Chao, A.; Chazdon, R. L.; Colwell, R. K. and Shen, T. J. 2005. A new statistical approach for assessing similarity of species composition with incidence and abundance data. Ecol. Letters. 8(2):148-159. [ Links ]
Cochran, W. G. 1977. Sampling techniques. John Wiley and Sons, Nueva York, USA. 448 p. [ Links ]
Colwell, R. K. 2009. EstimateS: statistical estimation of species richness and shared species from samples. Version 8.2.0. Department of Ecology and Evolutionary Biology, University of Connecticut. Connecticut, USA. User’s guide and application. http://viceroy.eeb.uconn.edu/estimates. [ Links ]
Cottam, G. and Curtis, J. T. 1956. The use of distance measures in phytosociological sampling. Ecology. 37(3):451-460. [ Links ]
Cueto, V. R. y López de Casenave, J. 1999. Determinants of bird species richness: role of climate and vegetation structure at a regional scale. J. Biog. 26(3):487-492. [ Links ]
Curts, J. 1993. Análisis exploratorio de datos. In: Salas, P. M. A. y Trejo, C. O. (Eds.) Las aves de la Sierra Purépecha del estado de Michoacán. SARH, Instituto Nacional de Investigaciones Forestales y Agropecuarias, Centro de Investigación Pacífico Centro. Boletín divulgativo núm. 79, Coyoacán, D. F., México. 1-14 pp. [ Links ]
Emlen, J. T. 1977. Estimating breeding season bird densities from transect count. The Auk. 94:455-468. [ Links ]
Enríquez-Lenis, M. L.; Sáenz, J. C. e Ibrahim, M. 2007. Riqueza, abundancia y diversidad de aves y su relación con la cobertura arbórea en un agropaisaje dominado por la ganadería en el trópico subhúmedo de Costa Rica. Agroforestería en las Américas. 45:49-57. [ Links ]
Figueroa-Sandoval, B. 1983. La investigación en labranza en México. Terra. 1:37-43. [ Links ]
Herrera, C. M. 1985. Determinants of plant-animal coevolution: the case of mutualistic dispersal of seeds by vertebrates. Oikos. 44(1):132-141. [ Links ]
JMP IN 8.0.2. 2009. Statistics for the Apple MacIntosh. Statistics and Graphics Guide, Version 8.0.2. Academic SAS Institute Inc. Cary, Carolina del Norte, USA. [ Links ]
Ibarra, A. C.; Arriaga-Weiss, S. y Estrada, A. 2001. Avifauna asociada a dos cacaotales tradicionales en la región de la Chontalpa, Tabasco, México. Universidad y Ciencia. 17(34):101-112. [ Links ]
Kaufman, L. and Rousseeuw, P. J. 2009. Finding groups in data: an introduction to cluster analysis. Wiley Series in Probability and Statistics, Nueva York, USA. 368 p. [ Links ]
Lantschner, M. V. y Rusch, V. 2007. Impacto de diferentes disturbios antrópicos sobre las comunidades de aves de bosques y matorrales de Nothofagus antarctica en el No Patagónico. Ecol. Austral. 17:99-112. [ Links ]
López-DeCasenave J.; Pellot, J. P.; Caziani, S. M.; Mermoz, M. an Protomastro, J. 1998. Responses of avian assemblages to a natural edge in a chaco semiarid forest in Argentina. The Auk. 115(2):425-435. [ Links ]
Luna-Cavazos, M.; Romero-Manzanares, A. y García-Moya, E. 2007. Distribución geográfica y ecológica de papas silvestres (Solanum l.), del altiplano potosino-zacatecano, México. Rev. Chapingo Ser. Hortic. 13(1):35-41. [ Links ]
Magurran, A. E. 1988. Ecological diversity and its measurement. Princeton University Press, Nueva Jersey, USA. 179 p. [ Links ]
Masera, O. R. 2001. Deforestación y degradación forestal en México. Departamento de Ecología Campus Morelia. Universidad Nacional Autónoma de México. Morelia, México. 13 p. [ Links ]
McCullagh, P. and Nelder, J. A. 1989. Generalized linear models. Second (Ed.). Chapman and Hall, Londres. 256 p. [ Links ]
Microsoft Excel. 2003. Excel. Microsoft Office for Windows. [ Links ]
Minitab 15.1.20. 2007. Minitab, Inc. [ Links ]
Molinero, L. M. 2003a. Análisis de la varianza. Asociación de la Sociedad Española de Hipertensión. Liga española para la lucha contra la hipertensión arterial. Madrid, España. 5 p. [ Links ]
Molinero, L. M. 2003b. Análisis de tablas de contingencia de más de dos variables cualitativas. Asociación de la Sociedad Española de Hipertensión. Liga Española para la lucha contra la Hipertensión Arterial. Madrid, España. 9 p. [ Links ]
Navarro, B. A.; Figueroa, S. B.; Martínez, M. M.; González, C. F. y Osuna, C. E. S. 2008. Indicadores físicos del suelo bajo labranza de conservación y su relación con el rendimiento de tres cultivos. Agric. Téc. Méx. 34(2):151-158. [ Links ]
Navarro, B. A.; Figueroa, S. B.; Ordaz, CH. V. M. y González, C. F. V. 2000. Efecto de la labranza sobre la estructura del suelo, la germinación y desarrollo de maíz y frijol. Terra. 18(1):61-69. [ Links ]
Navarro, S. A. G. y Benítez, D. 1993. Patrones de riqueza y endemismos de las aves. In: Flores, V. O. y Navarro, A. S. (Eds.). Biología y problemática de los vertebrados en México. Ciencias. 7:45-53. [ Links ]
Oosting, H. J. 1956. The study of plant communities. Freeman, San Francisco, USA. 185 p. [ Links ]
Pompa-García, M. 2008. Distribución especial de la pérdida de vegetación en ecosistemas áridos de México. MES. 3(3):13-22. [ Links ]
Preston, F.W. 1948. The commonness, and rarity, of species. Ecology. 29(3):254-283. [ Links ]
R 2.12.1. 2010. The R foundation for statistical computing version 2.11.0. (2010-03-15). ISBN 3-900051-07-0. [ Links ]
Ralph, C. J.; Geupel, G. R.; Pyle, P.; Martín, T. E.; DeSante, D. F. y Milá, B. 1996. Manual de métodos de campo para el monitoreo de aves terrestres. Department of Agriculture and Forest Service, USA. 46 p. [ Links ]
Ramírez-Albores, J. E. 2010. Diversidad de aves de hábitats naturales y modificados en un paisaje de la Depresión Central de Chiapas, México. Rev. Biol. Trop. 58(1):511-528. [ Links ]
Rappole, J. H.; Morton, E. S.; Lovejoy, T. E. y Ruos, J. L. 1993. Aves migratorias Neárticas en los Neotropicos. Ed. Conservation and research. Center National Zoological Park, Smithsonian Institution. Virginia, USA. 341 p. [ Links ]
Remsen, J. V. Jr. and Robinson, S. K. 1990. A classification scheme for foraging behavior of birds in terrestrial habitats. Studies in Avian Biology. 13:144-160. [ Links ]
Rzedowski, J. 1978. Vegetación de México. Editorial Limusa, México, Distrito Federal. 432 p. [ Links ]
Salinas, L., C. Arana y V. Pulido. 2007. Diversidad, abundancia y conservación de aves en un agroecosistema del desierto de Ica, Perú. Rev. Peruana de Biología. 13(3):155-167. [ Links ]
Sánchez, O. y López, G. 1988. A theoretical analysis of some indices of similarity as applied to biogeography. Folia Entomológica Mexicana. 75:119-145. [ Links ]
SEMARNAT. 2010. Norma Oficial Mexicana 059. Secretaría del Medio Ambiente y Recursos Naturales. México, DF. 78 p. [ Links ]
Tramer, E. J. 1969. Bird species diversity: components of Shannon’s formula. Ecology. 50(5):927-929. [ Links ]
Ugalde, L. S.; Valdez, H. J. I.; Ramírez, V. G.; Alcántara, C. J. L. y Velázquez, M. Y J. 2009. Distribución vertical de aves en un bosque templado con diferentes niveles de perturbación. Madera y Bosques. 15(1):5-26. [ Links ]
Received: January 2019; Accepted: April 2019
 Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
