











 text in
text in  English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkIntroducción
La alta variedad de componentes y metabolitos secundarios que existen entre especies de plantas e incluso hasta genotipos evita que exista un solo método universal estandarizado para el aislamiento de ácidos nucleicos en las plantas (Friar, 2005).
Para lograr la amplificación de regiones blanco se requiere partir de DNA con cantidad y calidad adecuadas (Rocha-Salavarrieta, 2002; Mafra et al., 2008; Sharma et al., 2013). Durante la elección del método de aislamiento de DNA vegetal, se debe de considerar a) el tipo de material del cual se parte (semilla, hoja, tallo u otros); b) tipo de DNA (genómico o de organelos); y c) el tipo de análisis que se llevará a cabo (Rocha-Salavarrieta, 2002).
Los reportes de metodologías en tejidos foliares frescos son más comunes que en semillas (Sharma et al., 2013) y se han publicado varios métodos para reducir las limitantes a las que se enfrentan durante el aislamiento de los ácidos nucleicos (Saghai-Maroof et al., 1984; Doyle y Doyle, 1990; Lopes et al., 1995; López-Mora et al., 2011; Pérez-Urquiza et al., 2013; Huang et al., 2013; Youssef et al., 2015).
No obstante, debido a la limitada eficiencia de algunos métodos de obtención de DNA (López-Mora et al., 2011), se realizan trabajos de comparación de protocolos para obtener ácidos nucleicos con buen rendimiento, pureza, integridad y funcionalidad (Demeke y Jenkins, 2010). Estos cuatro aspectos están determinados por la cantidad, el grado de degradación y el tamaño molecular de los ácidos nucleicos y la presencia de sustancias contaminantes inhibidoras de PCR (Elsanhoty et al., 2011; Sharma et al., 2013).
En cuanto a soya, existen reportes de métodos aplicados en alimentos y bebidas (Mafra et al., 2008) y a partir de las propias semillas (Demeke et al., 2009; Sharma et al., 2013). Aunque también se han empleado métodos comerciales por su rapidez (Chandu et al., 2016), la principal desventaja es el incremento en el costo (Demeke y Jenkins, 2010).
La concentración de DNA se puede estimar por espectrofotometría mediante la medición a 260 nm de longitud de onda, sabiendo que, una unidad de densidad óptica a 260 nm corresponde a 50 µg de DNA de doble cadena (Rocha-Salavarrieta, 2002; López-Mora et al., 2011).
Por otra parte, la pureza se mide con los parámetros de las relaciones A260/280 y A260/230. En la relación A260/280, el rango de valores esperados es superior a 1.7 a 2 y representa la cantidad de DNA en relación con las proteínas contenidas en la muestra (López-Mora et al., 2011); mientras que, en la relación A260/230, se espera que los valores sean cercanos a 2 e indica la presencia de polisacáridos y polifenoles que no fueron retirados durante el proceso de purificación del DNA (Demeke et al., 2009; Demeke y Jenkins, 2010) pero que, es importarte retirarlos porque pueden inhibir la acción de la Taq polimerasa durante el proceso de amplificación por PCR (Friar, 2005; Sharma et al., 2013).
La integridad es la medida del tamaño del peso molecular del DNA obtenido que puede ser determinado por comparación con un marcador de peso molecular de referencia, regularmente lambda DNA con un tamaño 48 Kb; a través, de la técnica de electroforesis (Sanger et al., 1982).
Finalmente, la funcionalidad está determinada por la capacidad de amplificación de un locus o varios loci. En la actualidad, muchas de las metodologías que se emplean en el análisis molecular se basan en la PCR, de tal manera que, asegurar que el material obtenido está libre de inhibidores de PCR es una medida de la funcionalidad. En el caso de las plantas, para determinar la presencia o ausencia de inhibidores pueden emplearse los genes rbcL o matK (Levin et al., 2003).
En el presente trabajo se compararon tres protocolos de aislamiento de DNA a partir de semilla de soya para obtener ácidos nucleicos de concentración y calidad adecuadas para realizar amplificación por PCR.
Materiales y métodos
Materiales y pulverización de la semilla
Los experimentos se llevaron a cabo en el Centro Nacional de Recursos Genéticos, Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias, dentro del laboratorio de ADN y Genómicas. Para el estudio se incluyeron seis muestras de diez semillas de soya.
Extracción de ácidos nucleicos
Las semillas se pulverizaron de manera mecánica en el equipo TissueLyser II de la marca Qiagen. Después, el polvo se transfirió a tubos de microcentrífuga de 2 mL y se almacenaron a -80 ºC hasta la extracción del DNA.
Se evaluaron tres protocolos para determinar el método adecuado de obtención de DNA: a) método convencional con solución buffer con CTAB (Saghai-Maroof et al., 1984) con modificaciones; b) método con solución buffer con sarcosina al 1% (Lopes et al., 1995) con modificaciones; y c) método con solución de fenol/cloroformo (Pérez-Urquiza et al., 2013) con modificaciones.
Método convencional con solución buffer con CTAB
Este protocolo se evaluó con dos diferentes concentraciones de CTAB, al 1% y 3%. El procedimiento fue el mismo en ambos casos, partiendo de 50 mg de semilla pulverizada en un tubo de 2 mL con 1 mL de solución buffer de extracción (TRIS-HCl 100 mM, pH 7.5, NaCl 700 mM, EDTA 50 mM, pH 8, 2-betamercaptoetanol (BME) 140 mM, polivinilpirrolidona al 1% y CTAB al 1% o 3%) previamente calentado a 65 ºC.
Los tubos se agitaron vigorosamente con un mezclador de vórtice y se incubaron a 65 ºC durante 90 minutos con agitación lenta y constante. Después, se dejaron a temperatura ambiente (TA) por cinco minutos, se agregaron 700 μL de solución cloroformo-octanol (24:1) y se mezclaron por inversión suavemente durante 10 min. Se centrifugaron a 4 500 x g durante 15 min a TA; se transfirió la fase acuosa a un nuevo tubo que contenía 700 μL de solución cloroformo-octanol (24:1) y se repitió este paso. El sobrenadante se transfirió a un nuevo tubo con 10 μL de RNAsa (10 mg mL-1), se mezcló por inversión y se incubó a 37 ºC durante 30 min.
Posteriormente, se adicionaron 800 μL de isopropanol frío y se mezcló por inversión hasta la precipitación del DNA. Los tubos se centrifugaron a 4 500 x g durante 10 min a TA para la formación del pellet, se decantaron y se agregó 1 mL de solución que contenía acetato de amonio 10 mM y etanol al 76%, se dejó a TA durante 20 min, se centrifugó a 4 500 x g durante 10 min a TA, se decantaron los tubos y se realizó un lavado con solución con acetato de amonio 10 mM y EDTA 0.25 mM. Posteriormente, las muestras se dejaron secar y se agregaron 200 μL de solución buffer TE (TRIS 10 mM y EDTA 1 mM, pH 8) hasta disolver totalmente el DNA.
Método con solución buffer con sarcosina al 1%
Se transfirieron 250 mg del pulverizado de semilla en un tubo de microcentrífuga de 2 mL, luego, se adicionó 1 mL de solución buffer de extracción (TRIS-HCl 100 mM, pH 8.5, NaCl 100 mM, EDTA 20 mM, pH 8, N-laurilsarcosina al 1%), se mezclaron permitiendo que la solución tuviera contacto completo con el polvo y se dejaron a TA durante 10 min.
Después, las muestras se mezclaron con 600 μL de fenol y se centrifugaron a 10 000 x g durante 15 min. El sobrenadante se transfirió a un tubo de microcentrífuga de 2 mL nuevo, que contenía 30 μL de RNAsa (10 mg mL-1), se dejaron incubar a 37 ºC durante 120 min. Nuevamente, se agregaron 600 μL de fenol y se centrifugaron a 10 000 x g durante 10 min. El sobrenadante se transfirió a un tubo nuevo con 600 μL de solución cloroformo-alcohol isoamílico (24:1), se mezcló vigorosamente y se centrifugaron las muestras a 10 000 x g durante 10 min.
Posteriormente, el sobrenadante se recuperó en un tubo nuevo con 600 μL de isopropanol frío, se mezclaron suavemente por inversión hasta observar el DNA precipitado, se centrifugaron a 10 000 x g durante 5 min para la obtención de pellet, después se descartó el sobrenadante y a través de absorción capilar con papel absorbente se eliminó la mayor cantidad posible del sobrenadante.
A continuación, el pellet se purificó con dos lavados, el primero con etanol al 70% y el segundo con etanol al 100% para eliminar proteínas y carbohidratos contaminantes. Las muestras se centrifugaron a 10 000 x g durante 5 min, se descartó el etanol y se dejaron a TA hasta que el etanol se evaporó completamente. Finalmente, el pellet de DNA se disolvió en 100 μL de solución buffer TE (TRIS 10 mM y EDTA 1 mM, pH 8).
Método con solución de fenol/cloroformo
En un tubo con 250 mg de tejido pulverizado se adicionaron 1.6 mL de solución buffer de extracción (tris base 50 mM, EDTA 5 mM y dodecilsulfato sódico 30 g L-1, pH 8) y 20 μL de proteinasa K (10 mg mL-1). Los tubos se dejaron incubar a 65 ºC en agitación lenta y constante durante 90 min. Posteriormente, se dejaron a TA por cinco minutos y se adicionaron 20 μL de RNAsa (10 mg mL-1).
Las muestras se centrifugaron a 5 000 x g durante 30 min a TA. Se recuperó el sobrenadante en un tubo con 800 μL de fenol, se mezcló de manera vigorosa y se centrifugó a 10 000 x g durante 5 min. Después, se recuperó la fase acuosa en un tubo con 800 μL de solución fenol/cloroformo-alcohol isoamílico (25:24:1) y se mezcló vigorosamente. Posteriormente, se centrifugaron a 10 000 x g durante 10 min, se recuperó la fase acuosa y se repitió este paso para obtener una interfase limpia.
Cada tubo con sobrenadante se mezcló con 800 μL de solución cloroformo-alcohol isoamílico (24:1) y se centrifugaron a 10 000 x g durante 5 min; se recuperó el sobrenadante y se mezcló por inversión con 30 μL de solución acetato de potasio 3 M (pH 5.2) y 750 μL de etanol al 96%. Se incubaron las muestras a -20 ºC durante 30 min. Después, se centrifugaron a 10 000 x g durante 5 min y se descartó el sobrenadante con cuidado de no verter el pellet y tratando de eliminar la mayor cantidad de líquido por absorción capilar con una toalla de papel.
A continuación, se agregó 1 mL de etanol al 70% y se agitaron los tubos hasta despegar el pellet para descartar las sales, se centrifugaron a 5 000 x g durante 5 min, luego se descartó el sobrenadante y se dejaron las muestras a TA hasta evaporar el etanol completamente. El DNA se disolvió en 100 μL de solución buffer TE (TRIS 10 mM y EDTA 1 mM, pH 8).
El DNA obtenido a través de los tres protocolos se almacenó en refrigeración a 4 ºC hasta el análisis de concentración y calidad.
Rendimiento de los métodos de extracción de DNA
El rendimiento de los métodos se determinó mediante la relación de la concentración de DNA y la cantidad de semilla pulverizada empleada. La cuantificación de ácidos nucleicos se realizó por espectrofotometría en un equipo NanoDrop 2000 TM (Thermo Scientific); a través, de la medición de la densidad óptica a 260 nm de longitud de onda. Previamente a la medición, las muestras se retiraron de refrigeración y se incubaron a TA durante 10 min.
Pureza de los ácidos nucleicos
La pureza de los ácidos nucleicos se determinó mediante los parámetros de las relaciones A260/280 y A260/230. Ambas se estimaron mediante espectrofotometría en un equipo NanoDrop 2000TM (Thermo Scientific); a través, de las medidas de densidad óptica a 230, 260 y 280 nm de longitud de onda.
Integridad de ácidos nucleicos
Con la finalidad de verificar el peso molecular del DNA genómico, se realizó electroforesis en gel de agarosa al 1% teñido con GelRed (Biotium®). En el gel se aplicaron 30 ng de DNA de cada muestra y se utilizó como marcador de referencia 3 µL de λ DNA (Promega®) sin cortar (10 ng µL-1). El corrimiento electroforético se realizó a 100 volts durante 45 min. Los geles se visualizaron con luz UV en un equipo transiluminador y capturó la imagen mediante fotografía digital.
Funcionalidad del DNA
La funcionalidad se verificó con la amplificación del gen rbcL mediante PCR. La mezcla de reacción se preparó con RedTaq 0.8X (Sigma-Aldrich®), iniciador sentido 0.35 μM, iniciador antisentido 0.35 μM y 10 ng de DNA. Se emplearon las secuencias de los iniciadores previamente descritas (Levin et al., 2003). Las condiciones de amplificación fueron: desnaturalización inicial a 95 ºC por 10 min, 40 ciclos de desnaturalización a 95 ºC por 40 s, hibridación a 55 ºC por 40 s y extensión a 72 ºC por 40 s, extensión final a 72 ºC por 1 min y mantenimiento a 4 ºC (Levin et al., 2003). Los productos de PCR se visualizaron por corrimiento electroforético a 100 volts durante 60 min sobre gel de agarosa al 2% teñido con GelRed (Biotium®). Se aplicaron 5 μL de producto del amplificado y se utilizó como marcador de referencia escalera de 100 pb (Promega®). Después, se preparó la dilución de trabajo a 10 ng µL-1 con un volumen de 100 µL.
Análisis estadístico
El diseño de experimento aplicado fue bloques completamente al azar con seis repeticiones, siendo los bloques las semillas de soya. Tres variables de respuesta fueron estudiadas: a) rendimiento; b) relación A260/280; y c) relación A260/230. El análisis estadístico se llevó a cabo con el uso del programa SAS, Versión 9.3 del sistema SAS para Windows (SAS Institute Inc., Cary, NC, USA). Se realizó el análisis Anova de tres variables cuantitativas y la comparación de las medias mediante la prueba Tukey con un nivel de significancia del 0.05.
Resultados y discusión
Rendimiento de los métodos de extracción de DNA
El rendimiento más alto se obtuvo con los protocolos que emplearon solución de CTAB al 3% y 1%, seguidos de los protocolos con soluciones de sarcosina y fenol/cloroformo (Cuadro 1). En todos los métodos se obtuvo buena cantidad de DNA, aunque no es el único parámetro a considerar para la realización de estudios moleculares posteriores.
Cuadro 1 Valores promedio del rendimiento y pureza de ácidos nucleicos obtenidos a partir de semillas de soya Glycine max (L.) Merrill. con los protocolos de extracción evaluados.
| Protocolo de extracción | Rendimiento (mg g-1) | Pureza | ||||
|---|---|---|---|---|---|---|
| 260/280 | 260/230 | |||||
| μ | s | μ | s | μ | s | |
| Solución con CTAB al 1% | 5.65 | 2.578 | 1.22 | 0.095 | 2.41 | 0.254 |
| Solución con CTAB al 3% | 11.93 | 4.058 | 1.14 | 0.027 | 0.62 | 0.46 |
| Solución con sarcosina al 1% | 0.76 | 0.165 | 0.76 | 0.08 | 1.6 | 0.328 |
| Solución de fenol/cloroformo | 0.07 | 0.016 | 1.97 | 0.016 | 2.14 | 0.247 |
μ= media; s= desviación estándar.
Aunque con el protocolo de fenol/cloroformo se obtuvo menor rendimiento de DNA (152 a 263 µg mL-1) con un promedio de 186 µg mL-1, ésta concentración es suficiente cuando el propósito es realizar amplificación de marcadores por PCR, como ha sido reportado en otros estudios en lo que también se ha obtenido menor cantidad de DNA a partir de semilla de soya con el método de CTAB (Sisea y Pamfil, 2007; King et al., 2014).
En los resultados del Anova se observaron diferencias significativas entre los protocolos utilizados, p> 0.0001 (Cuadro 2), mientras que, en la comparación de medias, se observan tres grupos diferentes: a) solución de CTAB al 3%; b) solución de CTAB al 1%; y c) soluciones de sarcosina y fenol/cloroformo (Cuadro 3).
Cuadro 2 Cuadrados medios y significancia de las variables rendimiento y pureza de DNA de semillas de soya Glycine max (L.) Merrill.
| Fuente de variación | GL | Cuadrado medio de rendimiento | Cuadrados medios de pureza | ||||
|---|---|---|---|---|---|---|---|
| A260/280 | A260/230 | ||||||
| Protocolo | 3 | 180 | *** | 1.5 | *** | 3.7 | *** |
| Error | 15 | 5.1 | 0 | 0.1 | |||
| CV (%) | - | 49.2 | 3.8 | 19.4 | |||
Gl= grados de libertad; CV= coeficiente de variación; ***= significativo con p≤ 0.0001.
Cuadro 3 Comparación de medias con la prueba de Tukey de las variables rendimiento y relaciones A260/280 y A260/230.
| Grupos | Rendimiento | Pureza | |||||
|---|---|---|---|---|---|---|---|
| A260/280 | A260/230 | ||||||
| Media | Protocolo | Media | Protocolo | Media | Protocolo | ||
| A | 11.93 | CTAB 3% | 2.09 | Sarcosina | 2.41 | CTAB 1% | |
| B | A | 2.14 | Fenol/cloroformo | ||||
| B | 5.65 | CTAB 1% | 1.97 | Fenol/cloroformo | 1.6 | Sarcosina | |
| C | 0.76 | Sarcosina | 1.22 | CTAB 1% | 0.62 | CTAB 3% | |
| C | 0.08 | Fenol/cloroformo | 1.14 | CTAB 3% | |||
Medias con la misma letra no tienen diferencias significativas (α= 0.05).
Las diferencias altamente significativas del rendimiento entre los protocolos se pueden deber a que, otras sustancias también son detectadas a 260 nm de longitud de onda, como puede ser DNA de cadena simple, fenoles, RNA y nucleótidos, las cuales no son distinguibles del DNA de cadena doble (Demeke y Jenkins, 2010). Esto coincide con los valores de la relación A260/280 inferiores observados con el uso de los protocolos con soluciones de CTAB y sarcosina (Cuadro 1), lo cual dificulta la determinación precisa de la cantidad de ácidos nucleicos (López-Mora et al., 2011).
Pureza de ácidos nucleicos aislados
Además de obtener la cantidad suficiente de DNA para realizar los análisis deseados, es de alta importancia tomar en cuenta la pureza. La relación A260/280 nos permite conocer la cantidad de ácidos nucleicos en relación con la cantidad de proteínas presentes en la muestra.
Con el uso de solución fenol/cloroformo, las muestras obtuvieron valores en la relación A260/280 de alta pureza (1.97 en promedio), mientras que con el uso de soluciones de CTAB al 1% y 3% y sarcosina, los valores indican pureza del DNA de baja calidad (Cuadro 1). Resultados similares fueron obtenidos por Kamiya y Kiguchi (2003) quienes a partir de semillas de soya, obtuvieron DNA de buena calidad (A260/280 de 1.74 a 1.81) con el uso de fenol/cloroformo/alcohol isoamílico.
Así mismo, con la solución con fenol/cloroformo, los valores de la relación A260/230 obtenidos tuvieron un valor promedio de 2.14. En contraste, con el uso de soluciones de CTAB al 1 y 3% y sarcosina, los valores sugieren muestras de DNA con pureza de baja calidad, como se observa en el Cuadro 1. De acuerdo con reportes previos, valores bajos en esta relación indican la presencia de inhibidores de PCR, como pueden ser los polisacáridos (Demeke y Jenkins, 2010), lo cual puede deberse al contenido de sacarosa, fructosa, rufinosa y estaquiosa, que corresponden al 25% de la composición de las semillas de soya (De Luna-Jiménez, 2007) incrementando hasta 48% en algunas variedades (Zhang et al., 2017), y que no son eliminados por completo durante el proceso de aislamiento. Con el uso de CTAB, cloroformo y las enzimas α-amilasa, pectinasa, celulasa y hemicelulasa los azúcares pueden ser removidos (Demeke y Jenkins, 2010).
En el análisis de varianza, se observan diferencias altamente significativas de las relaciones A260/280 y A260/230, entre los protocolos estudiados, p< 0.0001 (Cuadro 2). A través de la comparación de medias, en la relación A260/280, se obtienen tres grupos: a) solución de sarcosina; b) solución de fenol/cloroformo; y c) soluciones con CTAB al 1% y 3%; mientras que, en la relación A260/230, se agrupan los protocolos de la siguiente manera: a) soluciones de CTAB al 1% y fenol/cloroformo; b) soluciones de fenol/cloroformo y sarcosina; y c) solución de CTAB al 3% (Cuadro 3).
Integridad de ácidos nucleicos aislados
Con el objetivo de determinar si las muestras obtenidas cuentan con alto peso molecular y pueden ser útiles para conservación a largo plazo, se realizó corrimiento electroforético sobre gel de agarosa al 1%. En la electroforesis de ácidos nucleicos obtenidos con CTAB al 1% (Figura 1a) y fenol/cloroformo (Figura 1c) se obtienen bandas que indican DNA de alto peso molecular, de acuerdo con el marcador λ DNA, mientras que, con sarcosina no se observan bandas en el gel (Figura 1b). En el caso de las muestras que se obtuvieron con el protocolo que utiliza solución de CTAB al 3% no se consideró útil realizar la electroforesis debido a que se presentaron valores bajos en la relación A260/280 (rango de 1.11 a 1.17), que indican mala calidad de ácidos nucleicos.
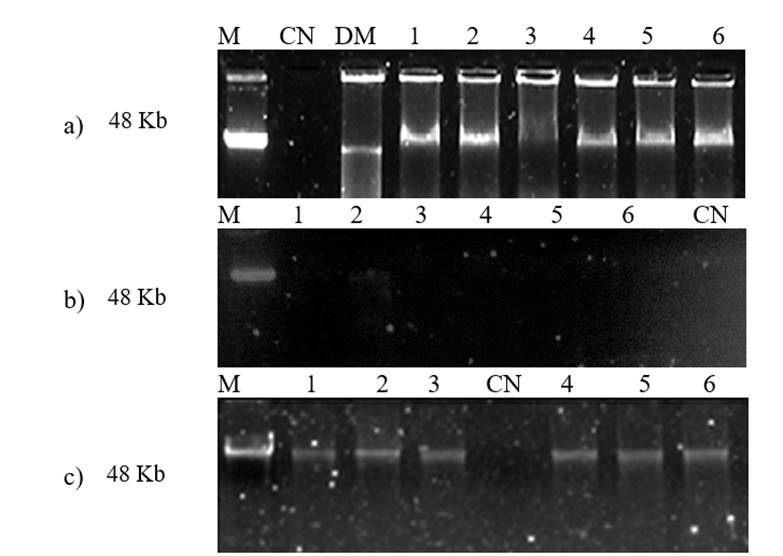
Figura 1 Electroforesis en gel de agarosa al 1% de los ácidos nucleicos obtenidos con tres protocolos de extracción. a) muestras de DNA aisladas con solución de CTAB al 1%; b) muestras de DNA aisladas con solución de sarcosina al 1%; y c) muestras de DNA aisladas con solución de fenol/cloroformo. M= marcador de referencia λ DNA; CN= control negativo; DM, DNA de maíz; 1-6= DNA de soya.
Durante el análisis de integridad de los ácidos nucleicos, se espera observar bandas sobre el gel con cierta intensidad y alto peso molecular revelado por el uso de un marcador de referencia. La presencia de barrido o manchas, es indicativo de DNA degradado, fragmentado o presencia de RNA contaminante (Figura 1). En el presente trabajo se observan bandas de intensidad similar al marcador de referencia cuando se empleó fenol/cloroformo, menor intensidad con CTAB y en sarcosina no se observan bandas, a pesar de que se presentaron lecturas altas a A260nm, lo cual puede deberse a productos contaminantes que son detectados a la misma longitud de onda.
Los resultados del presente estudio son similares a los reportados por Kamiya y Kiguchi (2003) quienes obtuvieron DNA con buena integridad a partir de semillas de soya con el uso de solución con fenol/cloroformo/alcohol isoamílico
Funcionalidad del DNA
La funcionalidad se evaluó únicamente con las muestras obtenidas con el protocolo con solución de fenol/cloroformo debido a que, los resultados de las pruebas de rendimiento, pureza e integridad de DNA fueron acordes con los parámetros esperados. En el corrimiento electroforético de la Figura 2 se observan las bandas de amplificación del gen rbcL en las seis muestras de DNA, lo que sugiere la ausencia de sustancias inhibidoras de PCR.

Figura 2 Electroforesis en gel de agarosa al 2% de los productos amplificados por PCR del gen rbcL. M= marcador de referencia escalera 100 pb; 1-6= amplicones con DNA de soya; CP1 y CP2= controles positivos 1 y 2 respectivamente, con DNA de maíz; CN= control negativo.
A pesar de que, el método con CTAB produjo mayor cantidad de DNA y es el más ampliamente utilizado en especies vegetales (Demeke et al., 2009), en el presente estudio se obtuvo mejor calidad con el método de fenol/cloroformo. A diferencia de los resultados que encontramos en el presente estudio, el método de CTAB fue efectivo para obtener DNA de alta calidad (Pinto et al., 2011) e incluso con tejido de cotiledón de semilla de soya (Al-Amery et al., 2016).
Con los resultados del presente trabajo, los laboratorios podrán optimizar recursos con la implementación del método apropiado para la obtención de ácidos nucleicos a partir de semilla de soya, pero también, puede ser aplicado en semillas de varias especies vegetales.
Conclusiones
La obtención de concentraciones elevadas de DNA con alguna metodología en particular, no necesariamente es indicativo de que el DNA sea útil para análisis posteriores como amplificación por PCR.
El método con solución de fenol/cloroformo es adecuado para obtener DNA en cantidad, pureza, integridad y funcionalidad apropiadas para realizar amplificaciones por PCR.
Los métodos de CTAB y sarcosina arrojaron valores bajos en las relaciones A260/280 lo que indica la presencia de proteínas, DNA degradado o de cadena sencilla o RNA, y en A260/230 que representa la presencia de carbohidratos o polifenoles inhibidores de PCR.

