










Services on Demand
Journal
Article
 text in
text in  English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista mexicana de ciencias agrícolas
Print version ISSN 2007-0934
Rev. Mex. Cienc. Agríc vol.9 n.3 Texcoco Apr./May. 2018
https://doi.org/10.29312/remexca.v9i3.1226
Investigation note
In vitro germination of biznaga cabuchera
1Instituto Tecnológico de Ciudad Victoria. Boulevard Emilio Portes Gil núm. 1301, Ciudad Victoria, Tamaulipas, México. CP. 87010. Tel. 01(834) 1532000, ext. 325. (erick-burrin@yahoo.com; wpoot@docentes.uat.edu.mx).
2Facultad de Ingeniería y Ciencias-Universidad Autónoma de Tamaulipas. Centro Universitario Adolfo López Mateos, Cd. Victoria, Tamaulipas, México. CP. 87149. Tel. 01(834) 3181800, ext. 2102.
3Posgrado en Estadística-Colegio de Postgraduados, Montecillo, Texcoco, México. CP. 56230. Tel. 01(595) 9520200, ext. 1459 y 1409. (antonio.rangel@itvictoria.edu.mx; hvaquera@colpos.mx; othonjavier@hotmail.com).
Ferocactus pilosus [(Galeotti ex Salm-Dyck) Werdermann] is a cactacea known as biznaga, which shows slow growth and is listed as a species at risk of extinction. The objective of the present study was to carry out an in vitro test in order to study the effect of chemical treatments and growth regulators, as promoters of seed germination collected in two consecutive years. The chemical products studied were H2SO4 and H2O2 and as the AG3, AIA and ANA phytoregulators, under different immersion and concentration times, respectively. The allocation of treatments was under a completely randomized design. For 30 days, the number of seeds germinated every 2 days was recorded and the standard germination was determined. Logistic regression was used to study the effect of the study factors on the proportion of germinated seeds. The standard germination of the biznaga is 82% with the H2SO4 treatment, while the germination of the seed starts between 2 and 6 days after sowing in both years and the representative logarithmic phase is between 8 and 20 days with H2SO4 in 2015. The biznaga seeds sampled in 2016 show greater slowness to cover the initial phase of germination, covered in 16 days. The application of H2SO4 shortened the germination time by 4 days and increased the standard germination by 82%, while the H2O kept the standard germination constant (±50%) and the germination response time was found between 6 and 10 days after the germination, planting in vitro.
Key words: Ferocactus pilosus; H2O2; H2SO4; germination time; growth regulator; standard germination
Ferocactus pilosus [(Galeotti ex Salm-Dyck) Werdermann] es una cactácea conocida como biznaga, la cual muestra crecimiento lento y se encuentra catalogada como una especie en riesgo de extinción. El objetivo del presente estudio fuel el realizar un ensayo in vitro con el fin de estudiar el efecto de tratamientos químicos y reguladores de crecimiento, como promotores de la germinación de semillas colectadas en dos años consecutivos. Los productos químicos que se estudiaron fueron el H2SO4 y H2O2 y como los fitoreguladores AG3, AIA y ANA, bajo diferentes tiempos de inmersión y concentración, respectivamente. La asignación de tratamientos fue bajo un diseño completamente al azar. Durante 30 días se registró número de semillas germinadas cada 2 días y se determinó la germinación estándar. Se utilizó la regresión logística para estudiar el efecto de los factores de estudio sobre la proporción de semillas germinadas. La germinación estándar de la biznaga es 82% con el tratamiento H2SO4, mientras que la germinación de la semilla inicia entre 2 y 6 días después de la siembra en ambos años y la fase logarítmica representativa se tiene entre 8 y 20 días con H2SO4 en 2015. Las semillas de biznaga muestreadas el año 2016 muestran mayor lentitud para cubrir la fase inicial de la germinación, cubierta en 16 días. La aplicación de H2SO4 acortó 4 días el tiempo de germinación y aumentó 82% la germinación estándar, mientras que el H2O mantuvo constante la germinación estándar (±50%) y el tiempo de respuesta de la germinación se localizó entre 6 y 10 días de la siembra in vitro.
Palabras clave: Ferocactus pilosus; H2O2; H2SO4; germinación estándar; regulador de crecimiento; tiempo de germinación
The arid zones occupy almost a third of the planet’s surface and are represented by hyper-arid, arid, semi-arid and sub-humid zones, with a diversity of soils, fauna, flora, water balance and human activity. There are 2 900 species of Cactaceae worldwide (Bravo-Hollis and Schneivar, 1999), while the American continent groups 2000 species (Jiménez, 2011). Of these, 1500 species develop in almost 56% of the Mexican territory (Hernández, 2006), with 73 and 78% of generic and specific endemism (Guzmán et al., 2003) and 35% are threatened (Hernández and Godinez, 1994). Ayudaran con las cosas de México
In this situation, there is the capercaillie [Ferocactus pilosus (Galeotti ex Salm-Dyck) Werdermann], distributed in Coahuila, Durango, Nuevo Leon, San Luis Potosí, Tamaulipas and Zacatecas (Bravo-Hollis and Sánchez-Mejorada, 1991). Which is subject to intense pressure for the collection and destruction of their habitat (Alanis and Velazco, 2008), which has forced it to modify structure and population density. In addition, its spread is limited it is vulnerable to predators and depends on nurses, competes with other species, suffers pathogen attack and is affected by temperature, humidity, light and soil (Lara et al., 2016). Consequently, it is a species at risk and under special protection (DOF, 2010), which is why it is necessary to recover and conserve its populations.
The ecological and biological knowledge of F. pilosus is scarce: little explored sexual reproduction, slow germination rate, poor establishment of seedlings and slow growth (Mandujano et al., 2007). Therefore, sexual reproduction under controlled conditions has been explored through the use of chemical compounds and growth regulators. In this way, Ferocactus has achieved 70 to 90% germination of the seed immersed 1.5 min in HCl (Navarro and González, 2007), which also accelerates the germination rate of F. histrix (Navarro et al., 2008). It also reached 80% of standard germination by immersing the seed in gibberellic acid (Rojas-Arechiga and Vázquez-Yanes, 2000). On the contrary, indole acetic and naphthaleneacetic acids do not promote the germination of F. histrix and F. latispinus (Amador-Alferez et al., 2013).
The propagation by seed of the biznaga would be convenient for obtaining individuals with better genetic characteristics (Ortiz-Hernández and Carrillo-Salazar, 2012), therefore, it is suggested to increase knowledge in seed germination (Sánchez-Villegas and Rascón-Chu, 2017), critical stage for the survival of F. pilosus. The objective of this work was to demonstrate that the application of chemical products and regulators of plant growth, represent an efficient means to increase the speed and standard germination of seeds of Ferocactus pilosus collected in two consecutive years.
The fruit collection was carried out in the common Estanque of the Walle, Miquihuana, Tamaulipas, Mexico (23° 34’ 15” north latitude and 99° 51’ 24” west longitude), at 1 560 meters above sea level. The climate is dry, annual precipitation of 350 to 500 mm and average temperature of 17 to 19 °C. The soil is Calcaric Regosol (Rc) of alluvial origin (SPP-INEGI 1983). Among the vegetation, Yucca filifera, Larrea tridentata and Agave lechugilla (Mora-Donjuan et al., 2014). The selection of individuals for fruit collection was made according to number (≤10) and length of shoots (≤1.5 m), in July 2014. Randomly selected 50 physiologically mature fruits that were kept between 4 and 5 °C by 30 days.
Seed extraction was carried out manually: the pulp was compressed against a mesh of 1 mm opening, with constant rinsings of drinking water. They were then distributed on brown paper until dried, at room temperature (25 °C) and stored in waxed envelopes until October 2015. That same year, another fruit harvest was made with the same protocol of 2014, and the seeds were preserved until October 2016. Seed selection was made considering the morphology parameters, color of the testa, without apparent damage and viability, the latter being measured with the precipitation/flotation test in distilled water reported by Emongor et al. (2004).
Germination tests were carried out under aseptic conditions in the laminar flow hood (ESCO, model AVC-4D2). The seeds were subjected to a cleaning process consisting of immersion for 10 min in 70% alcohol (Purex, Mexico), followed by three washes with sterile deionized water in Petri dishes with 5 min stirring. They were then immersed in a 15% sodium hypochlorite solution (Cloralex, Alen, Mexico) for 5 min, then washed in sterile deionized water as mentioned above.
The drying was carried out on absorbent paper before starting with the pre-germinative treatments which consisted of 1) chemical reagents: a) immersion in hydrogen peroxide (H2O2) at 3.34% for 10, 20 and 30 min in 2015 and 2016; b) sulfuric acid (H2SO4) at 97.30% for 5, 10 and 15 minutes in both years; and 2) plant growth regulators (RCV): a) gibberellic acid (AG3); b) indole acetic acid (AIA) and; c) naphthaleneacetic acid (ANA). In the latter, the seeds were immersed for 30 min in concentrations of 125, 250 and 500 ppm in both years of collection. The control treatment consisted of soaking the seeds in sterile deionized water for 30 min. The combination of the treatments allowed to obtain 16 in total for both years of collection.
The first germination experiment was established in October 2015. The Petri dishes were sterilized in autoclave (All American, 1941X, USA) at 1 atm. of pressure for 30 min. Each Petri box represented the experimental unit and was composed of 25 seeds and repeated four times. The boxes were incubated in a growing room with a relative humidity of 70% and a temperature of 25 ±2 °C, photoperiod of 16 h light (30 µmol m-2 s-1) per 8 h dark, emitted by cold-light fluorescent lamps (Illux, T5 Mod. 1 4100 °K, 50 W/120 V). The humidity in the Petri dishes was maintained by adding 10 ml of sterile deionized water every 48 h for 25 days, to recover the water consumed or lost, after counting the germinated seeds.
Germinated seed was considered, when exhibiting rupture of the testa and radicle emergence of 1 mm (ISTA, 2011). The second germinative experiment was carried out in September 2016 following the same procedure as in 2015. The statistical analysis included the logistic regression or binary regression in order to study the effect of the study factors: chemical compounds and phytoregulators, immersion time, concentration and years of collection on the proportion of germination. For this, the GENMOD procedure of the SAS ver program was used, 9.1.3 (SAS, 2007). The probabilistic model that is supposed to model the number of germinated seeds was that of the Binomial Distribution, with a logarithmic league function (Myers et al., 2002).
The model used is:
Donde: es la probabilidad de que una semilla germine dado los factores de estudio (x), Y= variable binario (1 germinar, 0 no germinar), es una función que refleja el efecto de factores son los parámetros asociados.
Where: P(Y=1|x) is the probability that a seed will germinate given the study factors (x), Y= binary variable (1 germinate, 0 not germinate), β0+ β1x1+…+ βp+xp is a function that reflects the effect of factors xi and βi are the associated parameters.
The chi-square test (χ2) verified specific differences between treatments. The choice of the best treatment was made by orthogonal contrasts according to the GENMOD procedure (De Menezes et al., 2016).
Latency is a strategy that seeds have to ensure germination when environmental conditions allow, however, this can be broken by the use of chemical compounds or RCV (Flores and Jurado, 2011) that help water imbibition and activation of cellular metabolism. The analysis of the maximum likelihood estimator of the logistic regression, for the year 2015, indicated that sulfuric acid was the compound that presented the highest percentage of germination (82%) of biznaga (p< 0.01) in comparison to the other treatments, including the control (Table 1, Figure 1). Likewise, in 2015 sulfuric acid as a pre-germinative treatment of biznaga seed, improved the germination time at the beginning of the 6th day and later stabilized until reaching 82% of germination; however, this behavior was different in the remaining treatments where germination started on the 9th day and stabilized after the 20th day, reaching 59% germination.
Table 1 Analysis of the estimation of the maximum likelihood parameter in the binomial distribution test due to the effect of chemical compounds and phytoregulators on the standard germination of biznaga seed in two years.
| Treatments | 2015 | 2016 | |||||
| VE | EE | χ2 | VE | EE | χ2 | ||
| H2O2 | -0.09 | 0.17 | 0.3 | -0.71 | 0.12 | 31.5 * | |
| H2SO4 | 0.39 | 0.13 | 8.99 * | -0.53 | 0.11 | 23.5 * | |
| AG3 | -0.06 | 0.17 | 0.13 | -0.71 | 0.12 | 31.5 * | |
| AIA | 0.05 | 0.15 | 0.14 | -1.21 | 0.17 | 46 * | |
| ANA | 0.27 | 0.19 | 2.8 | -0.47 | 0.1 | 20.6 * | |
| H2O | -0.59 | 0.11 | 26.2* | -0.17 | 0.04 | 15.9 * | |
VE= estimated value; EE= standard error; χ2= Chi square; *= significant (α≤ 0.05).
This type of response could be due to the biological phenomenon that normally occurs in seeds that germinate under environmental conditions of aridity (Jurado and Flores, 2005). That is, the germination of the seed would express the dependence of soil water retained among soil particles, minimal but fundamental compared to that required by other plant species (Loustalot et al., 2014). The greatest germination response is due to the scarification of the testa by the corrosive action of sulfuric acid (Flores and Jurado, 2011), morphological situation that favors the flow of water to accelerate the imbibition and the germinative process (Navarro and González, 2007).

Figure 1 Effect of chemical pre-treatments and growth regulators on the standard germination of seeds of Ferocactus pilosus sampled in 2015 and 2016. The lines indicate the standard deviation.
It has been observed that among the diet of species such as Neotoma sp., Scelophorus sp., Corvux corax, Melanerpes aurifrons and Campylorhynchus brunneicapillus are the fruits of biznaga or cabuches (personal observation) and that in turn contribute to the dispersing action (endozoocoric) of seeds. Therefore, the immersion of the seed in sulfuric acid simulates the scarification process that occurs in the digestive tract of these and other vertebrates, thus favoring germination (Escobar and Huerta, 1999).
In 2016, the germination of the seeds treated with pre-germinative compounds started approximately on the 2nd day of sowing, but with a slow germinative response until the 18th day. The RCV, gibberellic acid presented higher percentage of germination (48%) than the remaining treatments (p< 0.01) and the control (Table 1). The differential effect of AG3 on the germination response was slightly higher with 9% higher than the control (39%) and it was maintained from the beginning of germination. This same effect was reported in seeds of F. acanthodes, F. wislizeni, Echinocereus viridiflorus (Deno, 1994), Stenocereus griseus (Martínez-Cárdenas et al., 2006) and in the case of Harrisia fragrans, at a higher concentration (1 000 ppm) of AG3 (Dehgan and Pérez, 2005) obtained better response in the percentage of germination.
In this sense, AG3 improves germination through the hydrolysis of starch in the endosperm and promotes cell division in the meristematic tissues. It has been suggested that it can replace other stimuli such as light and temperature to initiate germination (Sánchez-Villegas and Rascón-Chu, 2017) and increases the germination response in seeds with some type of latency, that present physiological barriers or that have remained stored for long periods of time (Amador-Alférez et al., 2013).
The differences observed in the percentages of germination between 2015 and 2016 for H2SO4 (Figure 2) is attributed to variations in the thickness of the seed heads, which would allow greater or less resistance to scarification by this compound and presence of seeds with degrees of polymorphic latency within the same fruit or latency at the embryo level (Sánchez et al., 2015). It has been documented that H2SO4 in diluted solutions stimulates and accelerates the germination time, because it softens the membranes of the seed coat that may be suberized or impregnated with substances that make them impervious to water and oxygen.
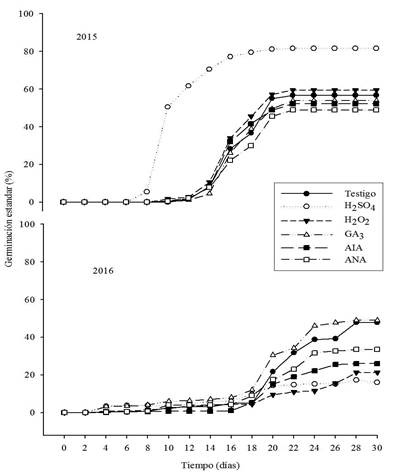
Figure 2 Accumulated germination of seeds of Ferocactus pilosus sampled in 2015 and 2016 and pre-treated with chemical compounds and growth regulators.
These variations in the positive germinative response have been reported in species of Mammillaria zephyranthoides (Navarro and Juárez 2006), M. myxtax (Navarro et al., 2010) and M. mainiae (Sánchez-Villegas and Rascón-Chu, 2017). Another effect due to the use of compounds with scarification properties is the inactivation of enzymes related to germination (Martínez-Cárdenas et al., 2006). In this study, it was detected that a time of immersion greater than 10 min in H2SO4, caused necrosis and death of the newly emerged radicle; therefore, it is suggested to reduce the concentration and immersion time of F. pilosus in H2SO4 at 1.5 min (Navarro and González, 2007).
The pre-germinative compounds applied in F. pilosus seeds in both years, decreased the germination time of approximately 10 to 3 days with respect to the control (Figure 1 and 2). The thickness and chemical composition of the seed coat may interfere with the water permeability in the seed and affect germination; this interference has been reported in cactus seeds (Navarro and Deméneghi, 2007), where the term physical latency is used to refer to the absence of germination as a result of water impermeability due to the seed coat. It has been suggested that F. pilosus presents hydration memory (Contreras-Quiroz et al., 2016a).
That is, after the seeds are dispersed in the field, they remain on the surface of the soil exposed to discontinuous hydration cycles being a key factor in the dynamics of desert plants, as an adaptive strategy to postpone germination in arid areas where climatic conditions are unpredictable, with annual rainfall less than 500 mm and that rainfall in a short time can produce a physiological effect in seeds with different degrees of dormancy synchronizing their germination in an accumulated way (Contreras-Quiroz et al., 2016a, 2016b).
Therefore, an advantage of the seed is to accelerate germination, due to the short time that the soil surface can retain moisture. The time and percentage of germination seems to be similar to what occurs when using H2SO4 or continuous application of H2O, in the same way that it responds with discontinuous hydration (Contreras-Quiroz et al., 2016a). So the seeds of biznaga under natural conditions require a disperser to soften the testa (gastric juices) and take advantage of the scarce rainwater retained in the soil particles to germinate (Loustalot et al., 2014; Sánchez-Villegas and Rascón-Chu, 2017).
Literatura citada
Alanís, F. G. J. y Velasco, M. C. G. 2008. Importancia de las cactáceas como recurso natural en el Noreste de México. Ciencia UANL. 11(1):5-11. [ Links ]
Amador, A. K. A.; Díaz, G. J.; Loza, C. S. y Bivián, C. E. 2013. Efecto de diferentes reguladores de crecimiento vegetal sobre la germinación de semillas y desarrollo de plántulas de dos especies de Ferocactus (Cactaceae). Polibot. 35(1):109-131. [ Links ]
Bravo, H. H. y Sánchez, M. H. 1991. Las cactáceas de México. Volumen II. Universidad Nacional Autónoma de México (UNAM). México, DF. 404 p. [ Links ]
Bravo, H. H. y Scheinvar, L. 1999. El interesante mundo de las Cactáceas. Fondo de Cultura Económica (FCE). Segunda edición. México, DF. 223 p. [ Links ]
Contreras, Q. M. Del R.; Pando, M. M.; Flores, J. and Jurado, E. 2016a. Effects of wetting and drying cycles on the germination of nine species of the Chihuahuan Desert. Bot. Sci. 94(2):221-228. [ Links ]
Contreras, Q. M.; Pando, M. M.; Jurado, E.; Bauk, K.; Gurvich, D. E. and Flores, J. 2016b. Is seed hydration memory dependent on climate? Testing this hypothesis with Mexican and Argentinian cacti species. J. Arid Environ.. 130(1):94-97. [ Links ]
De Menezes, G. L.; Machado, M. de F. P. S.; Ballesta, P.; Mora, F.; Milaneze, G. M. A. y Aparecida, M. C. 2016. Suplementos orgánicos para el cultivo in vitro del híbrido Laeliocattleya (Orchidaceae). IDESIA. 34(1):47-54. [ Links ]
Dehgan, B. and Pérez, H. E. 2005. Preliminary study shows germination of Caribbean Apple Cactus (Harrisia fragrans) improved with acid scarification and gibberellic acid. Native Plants J. 6(1):91-95. [ Links ]
Del Castillo, R. 1986. Semillas, germinación y establecimiento de Ferocactus histrix. Cact. Suc. Mex. 31(1):5-11. [ Links ]
Deno, N. C. 1994. The critical role of gibberellins in germination and survival of certain cacti. Cact. Succ. J. 66(1):28-30. [ Links ]
DOF. 2010. (Diario Oficial de la Federación). Norma Oficial Mexicana NOM-059-SEMARNAT-2010. Protección ambiental-especies nativas de México de flora y fauna silvestres-categorías de riesgo y especificaciones para su inclusión, exclusión o cambio-lista de especies en riesgo. 30 de diciembre de 2010. México, DF. 78 p. [ Links ]
Emongor, V. E.; Mathowa, T. and Kabelo, S. 2004. The effect of hot water, sulphuric acid, nitric acid, gibberellic acid and ethephon on the germination of Corchorus (Corchorus tridens) seed. J. Agron. 3(3):196-200. [ Links ]
Escobar, V. y Huerta, F. 1999. Relaciones ecológicas de Ferocactus histrix (DC.) Lindsay en los Llanos de Ojuelos, Jalisco-Zacatecas. Cact. Suc. Mex . 44(2):40-48. [ Links ]
Flores, J. y Jurado, E. 2011. Germinación de especies de cactáceas en categoría de riesgo del desierto chihuahuense. Rev. Mex. Cienc. Fores. 8(2):59-70. [ Links ]
Guzmán, U.; Arias, S. y Dávila, P. 2003. Catálogo de cactáceas Mexicanas. Universidad Nacional Autónoma de México (UNAM)-Comisión Nacional para el Conocimiento y Uso de la Biodiversidad (CONABIO). México, DF. 315 p. [ Links ]
Hernández, M. H. 2006. La vida en los desiertos mexicanos. Colección: la ciencia para todos No. 213. Fondo de Cultura Económica. México, DF. 188 p. [ Links ]
Hernández, M. H. y Godínez, A. H. 1994. Contribución al conocimiento de las cactáceas mexicanas amenazadas. Acta Bot. Mex. 26(1): 33-52. [ Links ]
ISTA. 2011. International Seed Testing Association Rules proposals for the international rules for seed testing 2011 edition ISTA. Bassersdorf, Switzerland. 53 p. [ Links ]
Jiménez, S. C. L. 2011. Las cactáceas mexicanas y los riesgos que enfrentan. Rev. Digital Universitaria. 12(1):1-23. [ Links ]
Jurado, E. and Flores, J. 2005. Is seed dormancy under environmental control or bound to plant traits? J. Veg. Sci. 16(5):559-564. [ Links ]
Lara, J. E. I.; Treviño-Carreón, J.; Estrada D. B.; Poot, P. W. A.; Vargas-Tristán, V. y Ballesteros-Barrera, C. 2016. Determinación de las especies nodriza de Ferocactus pilosus (Galeotti) Werderm. (Cactaceae) en Miquihuana, Tamaulipas, México. Rev. Mex. Agroecosist. 3(2):184-194. [ Links ]
Loustalot, L. E.; Malda, B. G. X.; Suzán, A. H.; Hernández, S. L.G. y Guevara, E. A. 2014. Estudio de germinación y crecimiento en semillas de Ferocactus histrix (De Candolle). Cact. Suc. Mex . 59(3):79-95. [ Links ]
Mandujano, M. C.; Golubov, J. y Rojas-Aréchiga, M. 2007. Efecto del ácido giberélico en la germinación de tres especies del género Opuntia (Cactaceae) del Desierto Chihuahuense. Cact. Suc. Mex . 52(2):46-52. [ Links ]
Martínez-Cárdenas, M. de L.; Cabrera-Jiménez, M. del C.; Carmona, A. y Varela-Hernández, G. J. 2006. Promoción de la germinación de semillas de Stenocereus griseus (Haworth) Buxbaum y Escontria chiotilla (Weber) Rose. Cac. Suc. Mex. 51(4):111-121. [ Links ]
Mora, D. C. A.; Rubio, C. E. A.; Alanís, R. E.; Jiménez, P J.; González, T. Mata, B. M. A. y Mora, O. J. M. 2014. Composición y diversidad vegetal de un área de matorral desértico micrófilo con historial pecuario en el noreste de México. Polibot. 38(1):53-66. [ Links ]
Myers, R. H.; Montgomery, D. C. and Vining, G. G. 2002. Generalized linear models, with applications in engineering and the sciences. John Wiley and Sons Press. New York, USA. 342 p. [ Links ]
Navarro, C. M. C.; Cervantes, O. G. y Lázaro, C. J. O. 2008. Efecto de la escarificación de semillas en la germinación de dos especies de Mammillaria. Zonas Áridas. 12(1):97-105. [ Links ]
Navarro, C. M. C.; Saldívar, S. S y Eliosa, L. H. R. 2010. Efecto de la escarificación y de la edad de semillas en la germinación de Mammillaria mystax. Zonas Áridas 14(1):196-205. [ Links ]
Navarro, M. C. y Deméneghi, A. P. 2007. Germinación de semillas y efecto de las hormonas en el crecimiento de Mammillaria pectinifera. Zon. Ári. 11(1):233-23. [ Links ]
Navarro, M. C. y González, E. M. 2007. Efecto de la escarificación de semillas en la germinación y crecimiento de Ferocactus robustus (Pfieff.) Britton & Rose (Cactaceae). Zonas Áridas. 11(1):195-205. [ Links ]
Navarro, M. C. y Juárez, M. S. 2006. Evaluación de algunos parámetros demográficos de Mammillaria zephyranthoides en Cuautinchán Puebla, México. Zon. Ári . 10:74-83. [ Links ]
Ortiz, H. H. Y. D. and Carrillo, S. J. A. 2012. Pitahaya (Hylocereus spp.): a short review Comunicata Sci. 3(4):220-237. [ Links ]
Rojas, A. M. y Vázquez, Y. C. 2000. Cactus seed germination: a review. J. Ar. Environ. 44(1):85-104. [ Links ]
Sánchez, J.; Muro, G., Flores, J.; Jurado, E. y Sáenz, M. J. 2015. Los bancos de semillas y su germinación en ambientes semiáridos. Ciencia UANL . 18(73):69-76. [ Links ]
Sánchez, V. A. y Rascón, C. A. 2017. Efecto de la escarificación química y del ácido giberélico en la germinación de Mammillaria mainiae. Cact. Suc. Mex . 62(1):4-12. [ Links ]
SPP-INEGI. 1983. (Secretaría de Programación y Presupuesto, Instituto Nacional de Estadística, Geografía e Información). Síntesis geográfica del estado de Tamaulipas. Dirección General de Geografía. México, DF. 158 p. [ Links ]
SAS (Statistical Analysis System). Institute Inc. 2007. SAS 9.1.3 for Windows Microsoft. SAS Institute Inc., Cary. North Carolina, USA. 212 p. [ Links ]
Received: April 00, 2018; Accepted: May 00, 2018
 Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
