











 text in
text in  English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkIntroducción
El bióxido de carbono (CO2), metano (CH4) y el óxido nitroso (N2O), gases de efecto invernadero, se han incrementado en la atmósfera desde tiempos pre-industriales debido a factores antropogénicos y este incremento contribuye al cambio climático; el CO2 en específico, se incrementó en 40%; pasó de 278 ppm alrededor de 1 750 a 390.5 ppm en 2011 (IPCC, 2013). Las causas del incremento en la concentración de gases de carbono en la atmósfera están bien documentadas, en el caso del CO2 son el uso industrial y doméstico de combustibles que contienen carbono (petróleo, carbón, gas natural y leña), la deforestación -que provoca la descomposición de la materia orgánica- y la quema de la biomasa vegetal (Jaramillo, 2004). De acuerdo con Maass (2009), el incremento de CO2 en la atmósfera produce un aumento en la retención de la radiación emitida por la superficie terrestre, alterando el régimen térmico e induciendo el calentamiento global del planeta mediante el efecto invernadero.
El efecto invernadero se debe a los gases atmosféricos llamados justamente de efecto invernadero o termoactivos, los más importantes son el vapor de agua (H2O) y el CO2 (Garduño, 2004). La importancia del vapor de agua y del CO2 en la atmósfera es tal que sin su presencia la temperatura promedio actual del planeta sería aproximadamente 33 °C más fría y por lo tanto el planeta estaría congelado (Schlesinger, 1997). El CO2 es un componente natural del aire y siempre ha existido en la atmósfera, lo que ha hecho el hombre es acrecentarlo y esto es lo que conlleva al aumento del efecto invernadero (Garduño, 2004).
Para determinar la biomasa de los árboles y generar sus ecuaciones alométricas se utilizan métodos directos o destructivos que implican la destrucción de los árboles evaluados, ya que como lo indican Fehse, et al. (2002) los métodos directos se basan en el derribo y pesaje de la vegetación. Por ello, debido a que no siempre es posible derribar y destruir los árboles a evaluar para generar sus ecuaciones alométricas, la mayoría de los trabajos acerca del tema Acosta et al. (2002); Díaz et al. (2007); Juárez (2008); Avendaño et al. (2009); han utilizado métodos destructivos para estimar la biomasa y se han enfocado en evaluar principalmente especies de interés maderable, se consideró importante estimar la biomasa y carbono mediante un método no destructivo para una especie de escaza importancia comercial, pero de gran importancia ambiental, como lo son las especies de la vegetación riparia, ya que aunque que se asume que esta vegetación juega un papel crucial en el ciclo del carbono es poca la información disponible sobre los almacenes de este elemento en dichos ecosistemas.
El objetivo de esta investigación fue generar las ecuaciones alométricas de biomasa y carbono mediante un método no destructivo para la especie arbórea (Alnus acuminata K.) de un transecto del bosque de galería del río Texcoco.
Materiales y métodos
El trabajo de campo se desarrolló en un transecto del bosque de galería del río Texcoco, localizado específicamente en la sección del río que atraviesa la comunidad de San Pablo Ixayoc, municipio de Texcoco, Estado de México, México. El transecto fue seleccionado bajo el criterio mejor estado de conservación y abundancia de la vegetación riparia, midió 960 m de longitud, inició en el puente Zacasolco y terminó en el puente Aclamaxa. Las coordenadas geográficas de ambos puntos son los 19° 28' 44.9" y los 19° 28' 56.6" de latitud norte y los 98° 47' 55.7" y los 98° 48' 43.8" de longitud oeste, ubicados a 2 512 y 2 463 msnm respectivamente.
El clima del área de estudio es semifrío con una temperatura media anual entre 5 y 12 °C. La temperatura media del mes más frío varía de-3a18°C y la del mes más cálido entre 6.5 y 22 °C siendo este el mes más húmedo de los climas semi fríos sub húmedos con lluvias en verano. La precipitación del mes más seco es menor a 40 mm y el porcentaje de lluvia invernal menor a 5 mm. En la zona la precipitación total anual varía a lo largo del gradiente altitudinal, registrándose precipitaciones de 800 mm en la parte baja y de hasta 1 200 mm en la parte más alta donde se encuentra el bosque de pino y pradera alpina (INEGI, 2001).
La vegetación arbórea del área de estudio varía también según el gradiente altitudinal, a la altura de San Pablo Ixayoc se presentan los bosques de encino, dominados por diferentes especies del género Quercus (encinos o robles), estos bosques se encuentran como una transición entre los bosques de coníferas y las selvas pudiendo alcanzar desde los 4 hasta los 30 m de altura; pueden ser abiertos o muy densos y se desarrollan en diversas condiciones ecológicas (Martínez, 2010). El área de trabajo se localiza sobre la cordillera Sierra Nevada, se presentan los suelos Feozem en la parte alta baja y Cambisol en la parte más alta ambos de textura media (INEGI, 2001).
Se realizó un barrido florístico cuantitativo del transecto, para ello se delimitaron parcelas de muestreo de 600 m2 (12 m de ancho por 50 m de largo), estas se establecieron de manera longitudinal y zigzagueante de acuerdo al cauce del río y abarcaban tanto la corriente de agua, que en esa época del año (nov. - oct., 2012) medía entre 1 y 1.5 m de ancho, así como ambas franjas del bosque de galería. En cada parcela de muestreo se contabilizaron todas las especies arbóreas presentes con un diámetro a la altura de pecho (DAP), medido a 1.3 m, mayor a 5 cm y se calcularon sus índices de valor de importancia relativa (IVIR) mediante la siguiente ecuación tomada de Stiling (1999).
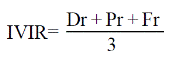
Donde: IVIR= índice de valor de importancia relativa; Dr= densidad relativa; Pr= predominio relativo; y Fr= frecuencia relativa.

Donde: D= densidad de cada especie; D1= densidad de todas las especies.
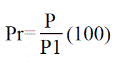
Donde: P= área basal de cada especie; P1= área basal de todas las especies.

Donde: F= número de parcelas en que ocurre cada especie; F1= número de parcelas muestreadas.
Las ramas se clasificaron en cuatro categorías diamétricas: delgadas, medianas, gruesas y extra gruesas. Se realizó un inventario de ramas por copa, se contabilizó el número de ramas presente en estas, se registró su categoría diamétrica respectiva y se podó en cada copa una rama de cada categoría diamétrica existente empleando una segueta forestal.
En el árbol 1 sólo se podó una rama ya que este presentó únicamente ramas delgadas, en el 2 se podaron dos ramas, este presentaba ramas delgadas y medianas. Del árbol 3 al 8 se cortaron tres ramas por ejemplar, estos árboles contaban con ramas delgadas, medianas y gruesas. Los árboles 9 y 10 presentaron las cuatro categorías de ramas, pero las primeras tres categorías formaban parte de la cuarta, por ello se consideró que estos árboles sólo presentaban el tipo de ramas extra gruesas, de esta categoría solo se cortó una rama por árbol.
En total se podaron 23 ramas: 8 delgadas, 7 medianas, 6 gruesas y 2 extra gruesas, todas fueron separadas en los componentes madera de las ramas (mr) y hojas (h). Los componentes de cada rama se pesaron con una báscula de reloj de 15 kg de capacidad y sus valores se agruparon y promediaron por tipo de rama para generar el peso fresco (PF) por componente según cada categoría diamétrica. Con esos valores y el inventario de ramas por árbol se calculó el aporte en peso fresco de cada componente a la biomasa de cada copa.
En campo se colectó una muestra de hojas de 500 g por árbol y de las 23 ramas se extrajo una rodaja basal de madera de aproximadamente 5 cm de grosor. En el laboratorio se obtuvo el PF de las rodajas de madera con una báscula marca Volke SF- 400 con capacidad de 5 kg y se colocó todo el material vegetal en una estufa de secado a 90 °C hasta que alcanzó un peso constante o peso seco (PS), esto sucedió a las 24 h en las muestras de hojas, a las 48 h en las rodajas con diámetros entre 5 y 20 cm y a las 96 h en las rodajas con diámetros mayores a 20 cm.
Los valores del PF y PS por componente se utilizaron para determinar su contenido de humedad en porcentaje (CH%). Los contenidos de humedad resultantes se promediaron por componente y los resultados se asumieron como su respectivo contenido promedio de humedad en porcentaje. Con los porcentajes promedio de humedad y el inventario de ramas se determinó el contenido de biomasa en peso seco de cada componente por copa (Cuadro 1) empleando para ello la siguiente fórmula:
Cuadro 1 Características dasométricas y contenido de biomasa y carbono de 10 árboles de aile (Alnus acuminata K).
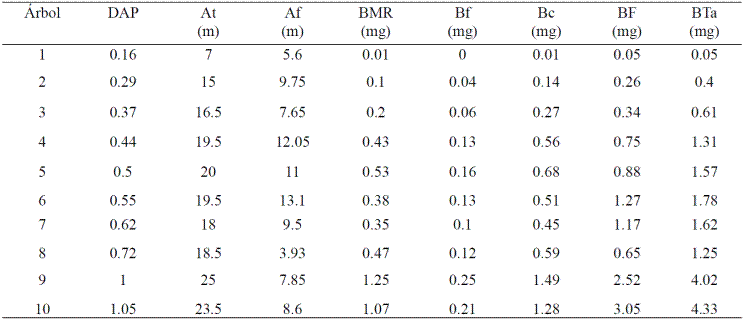
DAP= diámetro a la altura de pecho (1.3 m), At= altura total, Af= altura de fuste limpio, BMR= biomasa de la madera de las ramas, Bf= biomasa del follaje, Bc= biomasa de la copa, BF= biomasa del fuste, BTa= biomasa total por árbol.

Donde: B= biomasa en peso seco; Pf= Peso fresco; CH %= contenido de humedad en porcentaje.
Al sumar la biomasa en peso seco de la madera de las ramas (Bmr) y de las hojas (Bh) se obtuvo la biomasa total en peso seco de la respectiva copa (BC) (Cuadro 1).
De las 23 rodajas de madera se seleccionó al azar una por árbol y se les determinó su volumen empleando arena (mezcla comercial de piedra molida de la empresa Cantocreto Palsa S. A de C. V.) y un recipiente de plástico de forma neiloide (diámetro basal: 16 cm, diámetro superior: 13 cm, altura: 11.5 cm) al cual se le determinó su volumen (1 905.77 cm3) mediante la siguiente fórmula:

Donde: V= volumen (cm3); π=3.1416; h= altura; R= radio mayor; r= radio menor.
El procedimiento consistió en tamizar con una malla No. 60 una cantidad de arena del doble del volumen del contenedor, obteniéndose 3 500 g de arena tamizada que fueron depositados en una cubeta de plástico. Después en el contenedor neiloide se vertieron los 3 500 g de arena tamizada y el exceso de material formó un montículo que, sin presionarlo, fue desbastado con una regla metálica hasta el ras del contenedor, todo el material desbastado fue colectado y desechado. En seguida se pesó el contenedor junto con la arena que quedó en su interior después del desbaste (2 842.1 g) y este material fue regresado a la cubeta de plástico. Posteriormente, por cada determinación de volumen, en el contenedor neiloide se vertía directamente de la cubeta una capa de arena de aproximadamente 5 cm de espesor y sin presionarla se colocaba sobre esta la rodaja de madera a evaluar, después se vertía al interior del contenedor toda la arena restante en la cubeta.
Los fustes se subdividieron ascendentemente en secciones lo más cilíndricas posible y procurando que el cambio de dimensiones entre el diámetro basal y apical de cada sección fuera lo menos drástico posible. Los extremos de cada sección fueron marcados sobre los fustes con líneas horizontales de aproximadamente 15 cm hechas con una segueta forestal. Se midió el diámetro basal y apical de cada sección y el promedio de estos se consideró como el diámetro general de la sección respectiva mientras que la longitud existente entre ambos diámetros se consideró como su altura. Con estos datos se calculó el volumen de cada sección como si estas hubieran sido de forma cilíndrica perfecta, empleando para ello la siguiente formula:
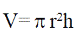
Donde: V= volumen; π=3.1416; r= radio; h= altura
La suma de las longitudes individuales de las secciones en que se subdividió el fuste se asumió como la altura de fuste limpio (Af l), y la suma de los volúmenes individuales de sus respectivas secciones se asumió como su volumen real o total. Con estos datos se determinó el factor de forma de los fustes el cual es un factor de reducción que indica la conicidad de un árbol y se expresa como el volumen real del árbol dividido por el volumen de un sólido cilíndrico de referencia cuyas dimensiones corresponden al DAP y altura de fuste limpio del árbol que representa (Romahn et al., 1994).
La biomasa de los fustes (BF) (Cuadro 1) se obtuvo con la siguiente fórmula propuesta por Fehse et al. (2002):

Donde: BF= biomasa del fuste (Mg); DM= densidad de la madera de la especie (Mg m-3); DAP= diámetro a la altura del pecho (m); Af l= altura de fuste limpio (m); Ffp= factor de forma promedio de la especie.
Al sumar la biomasa de la copa (BC) a la biomasa del fuste respectivo (BF) se obtuvo la biomasa total por árbol (BTA).
Adicionalmente se cuantificó el porcentaje de carbono de la madera de las ramas y las hojas, para ello, de los 10 árboles evaluados se eligieron cinco cubriendo equitativamente el rango de DAP y se analizó por cada árbol una muestra representativa de la madera de las ramas y una de las hojas. De cada muestra se molieron 100 g de material vegetal hasta pulverizarlo empleando un molino Thomas Wiley Mill Modelo ED-5; en el caso de las rodajas de madera, estas primero tuvieron que ser convertidas en astillas delgadas usando un formón de carpintero y un mazo. Las 10 muestras se analizaron en el auto determinador automático de carbono marca Shimadzu Modelo TOC SSM 5050A de acuerdo a los procedimientos del Laboratorio de Fertilidad de Suelos del Colegio de Postgraduados (Etchevers, 1992).
El contenido de carbono de cada muestra se usó para obtener el porcentaje promedio de carbono por componente. Debido a que no se derribaron árboles, el promedio de carbono de la madera de las ramas se empleó como el porcentaje promedio de carbono de los fustes. Con los valores de carbono por componente y los datos de biomasa en peso seco de la copa y de los fustes se determinó el contenido de carbono de estas estructuras mediante la siguiente expresión tomada de Díaz et al. (2007):

Donde: CCC= contenido de carbono del componente; BTC= biomasa total del componente (kg); (%) C= porcentaje de carbono del componente. Al sumar el contenido de carbono de la copa al contenido de carbono del fuste respectivo se obtuvo el contenido de carbono total por árbol (CCA) (Cuadro 1). Para generar las ecuaciones alométricas de biomasa y carbono se emplearon dos modelos: un lineal y un exponencial, como lo indican Rügnitz et al. (2008); Gayoso (2002) respectivamente y expresados de la siguiente manera:
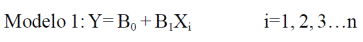
Donde: Y= biomasa (Mg); X= DAP (cm) y B0 y Bi son los parámetros del modelo.

Donde: Y= biomasa (kg); X= DAP (cm); y b y k son los parámetros de la función.
El ajuste de los modelos para generar las ecuaciones alométricas propuestas se realizó empleando el programa de análisis estadístico SAS (2002).
Resultados y discusión
Se inventariaron 762 árboles pertenecientes a 9 especies con los siguientes IVIR, aile (Alnus acuminata K.): 36.81%, fresno (Flaxinus uhdei W.): 14.73%, encino (Quercus rugosa N.): 14.12%, sauce llorón (Salix spp.): 11.92%, tepozán (Buddleia cordata K.): 7.62%, pirul (Schinus molle L.): 6.37%, cedro (Cupressus lusitanica M.): 4.63%, tejocote (Crataegus spp.): 3.33% y eucalipto (Eucaliptus spp.): 0.46%.
Respecto a la biomasa de las copas, las ramas de categoría diamétrica: delgadas, medianas, gruesas y extra gruesas presentaron diámetros basales que midieron entre 5 y 10, 10 y 15, 15 y 20, y entre 20 y 25 cm respectivamente, con un peso fresco promedio de la madera de 3.5, 15.2, 49.5 y 174.6 kg y de las hojas de 2.5, 7.5, 23.5 y 46.6 kg respectivamente. Mientras que sus valores promedio en peso seco para la madera de las ramas y las hojas fueron de 1.79, 7.75, 25.25, y 89.05 kg y 0.95, 2.85, 8.93 y 17.71 kg respectivamente. La madera de las ramas y las hojas presentaron porcentajes de humedad que variaron de 43 a 57% y de 55 a 68% con promedios de 49 y 62% respectivamente. La madera de las ramas aportó una mayor cantidad de biomasa a la copa en comparación con las hojas, en promedio la madera de las ramas aportó a cada árbol 0.42 Mg de biomasa y las hojas 0.1 Mg.
Respecto a la biomasa del fuste, el número mínimo y máximo de secciones en que estos fueron delimitados varió de 4 a 7, en todas el diámetro basal fue mayor al apical, no hubo dos secciones que midieran lo mismo ni dentro de un mismo árbol ni dentro de todos los árboles, la sección más corta midió 30 cm de longitud y la más larga 3.5 m. Entre mayor fue la altura de los fustes mayor fue también el número de secciones en que estos se delimitaron, la altura de fuste limpio vario de 3.93 a 13.10 m y el factor de forma (Ffp) de 0.5 a 0.75 con un valor promedio de 0.66. Los resultados mostraron, con excepción de los árboles 7 y 8, que el volumen de los fustes limpios no fue proporcional a su longitud si no a su DAP, entre mayor fue el DAP mayor fue también su volumen. Los volúmenes variaron de 0.08 a 4.38 m3 para los árboles con menor y mayor DAP respectivamente y el volumen promedio fue de 1.66 m3. Su biomasa varió de 0.05 a 3.05 Mg para los árboles con el menor y mayor DAP respectivamente.
El porcentaje de la biomasa en los árboles se distribuyó de la siguiente manera: 64.92, 27.06 y 8.02 en el fuste, la madera de las ramas y las hojas respectivamente. El contenido de carbono varió de 48.2 a 50.3% y de 46.5 a 47.1% en la madera de las ramas y el follaje con promedios de 48.9% y 47.1% respectivamente. Estos resultados varían a los reportados en trabajos similares donde el follaje presenta mayor contenido de carbono (Díaz et al., 2007; Juárez, 2008; Gómez et al., 2011). Esto pudo deberse a la alta proporción (50%) de ramillas en las muestras de follaje analizadas. Al multiplicar los valores de la biomasa en peso seco del componente madera de las ramas y follaje por sus respectivos porcentajes promedio de contenido de carbono se obtuvo el contenido de carbono de estos componentes por árbol (CCMR y CCf) y al sumar estos se definió el contenido de carbono de cada copa (CCc) (Cuadro 1).
Cuadro 1 Contenido de carbono en 10 árboles de aile (Alnus acuminata K.).

DAP= diámetro a la altura de pecho (1.3 m); CCMR= contenido de carbono en la madera de las ramas; CCf= contenido de carbono en el follaje; CCc= contenido de carbono de la copa; CCF= contenido de carbono del fuste; CCa= contenido de carbono por árbol.
El contenido de carbono de los fustes (CCF) se obtuvo al multiplicar el valor de su biomasa por el porcentaje promedio del contenido de carbono en la madera de las ramas. Se utilizó este último porcentaje ya que al no haberse derribado árboles no se contaba con muestras de madera del fuste y en general el contenido de carbono en los fustes siempre es más similar al de las ramas en comparación con el del follaje (Gómez et al., 2001; Díaz et al., 2007; Juárez, 2008). Finalmente la suma del contenido de carbono de la copa (CCc) y del fuste (CCF) generó el valor del contenido de carbono en cada árbol (CCa) (Cuadro 1).
Las ecuaciones alométricas generadas con los valores del DAP de los 7 árboles y determinadas mediante el modelo lineal presentaron un coeficiente de determinación altamente satisfactorio (R2= 0.99) y quedaron expresadas como:

Donde: B y CCAson el contenido de biomasa (Mg) y carbono por árbol (Mg) respectivamente y DAP es el diámetro a la altura de pecho (1.3 m). Sin embargo, ambas ecuaciones generaron valores negativos de biomasa y carbono al emplearlas en DAP menores a 23 cm, a diferencia de las ecuaciones generadas con el modelo exponencial que aunque con coeficientes de determinación menores (R2= 0.95) estas no generaron valores negativos al emplearlas en DAP menores a 23 cm. La ecuación alométrica para estimar biomasa en aile (Alnus acuminata K.) utilizando el modelo exponencial, quedó expresada como:

Donde: B es el contenido de biomasa por árbol (Mg) y DAP es el diámetro a la altura de pecho.
Los parámetros del modelo exponencial y la dispersión de puntos de los valores de biomasa observados se presentan en la Figura 1.
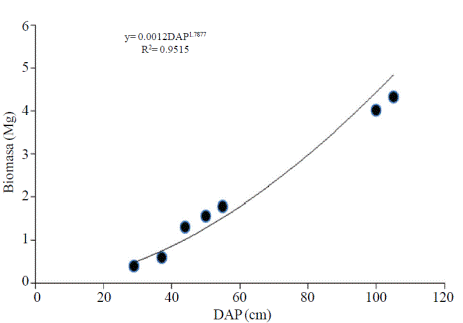
Figura 1 Dispersión de los valores de Alnus acuminata K. y la línea de regresión generada con los datos de contenido de biomasa utilizando el modelo exponencial.
Al ajustar la ecuación para determinar la biomasa en función del DAP se presentó un coeficiente de determinación de 0.95.
La ecuación alométrica para estimar el contenido de carbono en aile (Alnus acuminata K.) con el modelo exponencial, quedó expresada como:

Donde: CCA es el contenido de carbono por árbol (Mg) y DAP es el diámetro a la altura de pecho.
Los parámetros del modelo exponencial y la dispersión de puntos de los valores de carbono observados se presentan en la Figura 2.
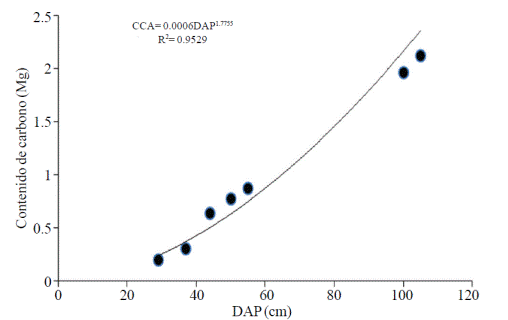
Figura 2 Dispersión de los valores de Alnus acuminata K. y la línea de regresión generada con los datos de contenido de carbono utilizando el modelo exponencial.
Al ajustar la ecuación para determinar el contenido de carbono en función del DAP se presentó un coeficiente de determinación de 0.95.
El alto IVIR del aile (36.81%) estuvo influenciado por sus porcentajes de densidad (27.82%) y frecuencia relativa (26.32%), pero fundamentalmente fue determinado por su porcentaje de predominio (56.30%), esto indicó un desarrollo sobresaliente de sus fustes en comparación con los de las otras ocho especies. Estos valores coincidieron con lo observado en campo donde sus DAP fueron desde los 5 hasta los 105 cm, y la altura de un gran número de sus elementos fue mayor a 20 m, mientras que en fresno (Flaxinus uhdei W.), la especie con el segundo mayor IVIR (14.73 %), los DAP de la especie sólo midieron de 5 a 74.80 cm y su altura en general fue menor a la de los ailes. El marcado desarrollo de los ailes se atribuyó a que por su altura captaban mayor radiación solar y podían fotosintetizar mas, además de encontrarse creciendo en un medio ambiente favorable para su crecimiento, ya que como lo señala Rzedowski (1978), al igual que en muchas otras partes del mundo, los bosques de Alnus (“aile”) en México son de dos afinidades ecológicas principales: viven a lo largo de arroyos y pequeños ríos, o bien constituyen comunidades sucesionales, surgidas como consecuencia de la destrucción de otros tipos de bosque.
Los valores de biomasa observados en las copas y derivados del aporte de madera de las ramas y hojas mantuvieron el patrón reportado en otros trabajos (Acosta et al., 2002; Díaz et al., 2007; Avendaño et al., 2009) en donde estos aumentan de forma directa al incrementarse los valores dasométricos de los árboles. Sin embargo, se considera que para obtener una mayor precisión al determinar la biomasa de la copa mediante el método empleado es conveniente podar un mayor número de ramas por copa en vez de solo podar una por categoría diamétrica o en su defecto realizar una clasificación más eficiente de ramas como puede ser el combinar con el diámetro la variable longitud de ramas.
La biomasa contenida en el fuste aumentó al aumentar su DAP más que su altura y este patrón se conservó en la mayoría de los árboles excepto en el 7 y 8. El fuste del árbol 7, a pesar de tener 10 cm más de DAPque el árbol 6, presentó un contenido de biomasa (1.17 Mg) ligeramente menor al del árbol 6 (1.27 Mg). El árbol 8 a pesar de su DAPde 0.72 m se vio severamente afectado por la escaza longitud de su fuste (3.93 m) lo que incidió para que este presentara una marcada disminución en la biomasa de esta estructura, ya que aunque por su DAP se esperaría que su fuste presentara mayor altura y en consecuencia alrededor de 1.5 Mg de biomasa, en realidad solo presentó 0.65 Mg; es decir, la mitad de lo esperado.
Esto pudo deberse a que muchos árboles a pesar de tener un DAP pronunciado sus fustes en un cierto punto tienden a bifurcarse o en algún punto de su desarrollo pudieron ser podados por los pobladores de la comunidad. Sin embargo, se mantuvo el patrón reportado en trabajos relacionados al tema (Acosta et al., 2002; Díaz et al., 2007; Avendaño et al., 2009) en donde se reporta que el aporte del fuste a la biomasa del árbol aumenta de forma directa al incrementarse los valores dasométricos de los árboles. Respecto al factor de forma obtenido (0.66) se consideró que al ser menor a 1 fue calculado de manera satisfactoria ya que como lo señala Romahn et al. (1994), tratándose de fustes el valor del coeficiente mórfico o factor de forma siempre es menor que la unidad.
El contenido de carbono total por árbol tendió a incrementarse al aumentar el DAP, la altura total del árbol (At) y la altura de fuste limpio (Af l), estos valores f luctuaron de 0.03 a 2.12 Mg para los árboles con el mayor y menor DAP respectivamente. El fuste aportó la mayor parte del carbono a los árboles en comparación con la copa, en promedio el fuste aportó 0.54 y la copa 0.29 Mg por árbol (1), estas proporciones coinciden con las reportadas en trabajos referentes al tema (Díaz et al., 2007; Juárez, 2008; Gómez et al., 2011). Por lo que se consideró que no hubo un efecto adverso al utilizar el porcentaje de carbono de las ramas para estimar el porcentaje de este elemento en los fustes.
Respecto al contenido promedio de carbono en la madera de las ramas y las hojas los valores obtenidos (48.9 y 47.1%), variaron ligeramente a los reportados en trabajos similares en donde las hojas presentan mayor contenido de carbono (Díaz et al., 2007; Juárez, 2008; Gómez et al., 2011). Esto pudo deberse a la alta proporción de ramillas, alrededor de 50%, en las muestras de hojas analizadas en este trabajo. Sin embargo, se conservó el patrón observado en relación al aporte parcial de carbono por parte del fuste, ramas y hojas al contenido total de este elemento en el árbol.
La eliminación de los árboles 1, 7 y 8 del análisis, generó ecuaciones alométricas con mayores coeficientes de determinación en ambos modelos. Los coeficientes de determinación obtenidos, mayores a 0.9, indicaron que en la determinación de biomasa y carbono en Alnus acuminata K. es confiable tomar el DAP como variable explicatoria. Se consideró que el mejor modelo para generar las ecuaciones alométricas fue el exponencial, coincidiendo con Gayoso (2002) quien señala que existe preferencia por este modelo debido a que expresa una proporcionalidad de los incrementos relativos entre dos componentes del árbol, aunado a que es ampliamente consistente para distintas formas de crecimiento.
Conclusiones
El aile (Alnus acuminata K.) fue la especie arbórea con el mayor IVIR (36.81%) del transecto del bosque de galería estudiado.
El método del recipiente de volumen conocido y arena empleando para determinar el volumen de las muestras de madera fue efectivo debido a que la arena se amoldó eficazmente a cualquier forma de las muestras y la cantidad de arena desplazada fue siempre similar al volumen de la muestra evaluada.
El método no destructivo empleado para el cálculo de la biomasa fue ef icaz al haber permitido determinar esta variable sin necesidad de derribar los árboles y al haberse mantenido el patrón observado en estudios similares en donde el fuste es la estructura que aporta al árbol la mayor cantidad de biomasa y carbono, seguido de la madera de las ramas y las hojas.
Los coeficientes de determinación obtenidos (R2) mayores a 0.9 indican que las ecuaciones determinadas para estimar biomasa y carbono para Alnus Acuminata K. son confiables para dicha especie en condiciones semejantes y dentro de rango de DAP evaluados.
El modelo exponencial fue el que mejor estimó el contenido de biomasa y carbono ya que no genera valores negativos al emplearlo en arboles con DAP menores a 23 cm. El contenido promedio de carbono en la especie fue de 48.84%.

