










Services on Demand
Journal
Article
 text in
text in  English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista mexicana de ciencias agrícolas
Print version ISSN 2007-0934
Rev. Mex. Cienc. Agríc vol.6 n.spe11 Texcoco May./Jun. 2015
https://doi.org/10.29312/remexca.v0i11.781
Investigation notes
Phenological variability of a population of bean Patol (Phaseolus coccineus L.) native of Durango, Mexico
1Campo Experimental Valle de México- Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias. Carretera Los Reyes-Texcoco, km 13.5. A. P. 307, C. P. 56250, Tel: (595) 92 126 57. Coatlinchán, Estado de México. México.
2Instituto Politécnico Nacional, CIIDIR-Unidad Durango, Avenida Sigma 119, Fracc. 20 de Noviembre II, Durango, Durango, México. C. P. 34220. (ghiturbide@hotmail.com).
3Centro de Biotecnología Genómica-Instituto Politécnico Nacional. Blvd. Del maestro, esquina Elías Piña s/n, Col. Narciso Mendoza. Reynosa, Tamaulipas, México. (nmayek@ipn.mx).
The collection of native varieties of Phaseolus coccineus L. of the National Research Institute of Forestry, Agriculture and Livestock has 798 sheltered ex situ accessions. Representing varieties of the centre and south of the country and are still needed to include those the northern territory. The plant P. coccineus L. has a mixed mating system with predominance of crosslinking so that a population can have multiple colours of seeds and the plants display different phenology. The genebank received a collection of the Ejido San José de las Corrientes, Vicente Guerrero, Durango, Mexico. Seed colour variants were defined and plated on June 25, 2010 and May 17, 2011 in Chapingo, Mexico. We recorded: photoperiod, days and accumulated temperature of seedling emergence at flowering, and days and temperature required to start flowering at the beginning of maturity. Short-cycle variants were identified, 123-144 days and long cycle of 165 days. In May plantings, the plant bloomed for 26-36 days with 13.17 and 13.19 hours of light; and plantings on June 40-56 days to 12.33 and 12.49 hours of light due to natural photoperiod decreased over time. When the growing season increases, so does the accumulation of temperature due to the increase occurring in spring and summer at this latitude. When planting in May accumulated in plants growing season, 468-719 °C, and in June 713-990 °C.
Keywords: Phaseolus coccineus L.; domesticated; plant genetic resources; plant phenology; sowing date
La colección de variedades nativas de Phaseolus coccineus L. del Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias tiene 798 accesiones resguardadas ex situ. Están representadas las variedades del centro y sur del país y hace falta introducir las del norte del territorio. La planta de P. coccineus L. tiene sistema de apareamiento mixto con predominancia del entrecruzamiento por lo que una población puede tener varios colores de semilla y las plantas mostrar diferente fenología. El banco de germoplasma recibió una colecta del Ejido San José de las Corrientes, Municipio de Vicente Guerrero, Durango, México. Se definieron variantes por colores de semilla y se sembraron 25 de junio de 2010 y el 17 de mayo 2011 en Chapingo, México. Se registró: fotoperiodo, días y temperatura acumulada de emergencia de plántula a inicio de floración, y días y temperatura requerida de floración a inicio de madurez. Se identificaron variantes de ciclo corto, 123-144 días, y de ciclo largo de 165 días. En siembras de mayo la planta floreció a los 26-36 días con 13.17 y 13.19 horas luz; y en siembras de junio a los 40-56 días con 12.33 y 12.49 horas luz debido a la disminución de fotoperíodo natural a través del tiempo. Al alargarse el período vegetativo aumentó la acumulación de temperatura debido al incremento que ocurre en primavera-verano en esta latitud. Al sembrar en mayo las plantas acumularon en período vegetativo, 468-719 °C, y al sembrar en junio 713-990 °C.
Palabras clave: Phaseolus coccineus L. domesticado; fecha de siembra; fenología vegetal; recursos genéticos vegetales
A population is a community of organisms of cross and sexual reproduction that share frequency of genes adjusted to the prevailing conditions (Dobzhansky, 1951). In cross-pollinated plants, the high percentage of natural cross-pollination fosters a constant genetic exchange through several generations so their populations are highly heterogeneous. This paper studies the phenology of a population of P. coccineus L., which has a mixed mating system with predominance of crosslinking (Coello and Escalante, 1989). Identify the processes that define their vegetative and reproductive periods (flowering and maturity) and infers the best time of year to regenerate and increase their seed that would be part of the genebanks´collections for long term safekeeping.
This is a kind of bean native of the highlands of Mexico and Central America, where the cultivated form grows near the wild (Miranda-Colín, 1967). It is found from Chihuahua to Panama in humid highlands at altitudes 1 500-3 000 m where coexist cultivated and wild and its centre of origin and domestication is Mesoamerica (Delgado-Salinas, 1988). It is a climbing perennial plant, which can also be grown as an annual, with a cross-pollination reproduction system and is the only species of the genus pollinating using hummingbirds (Búrquez and Sarukhan, 1980), although other pollinators are bees and bumblebees. Recent studies indicated that, the molecular genetic markers in Mesoamerica, the cultivated acquis is closely related to the wild, the current gene flow between wild and domesticated is high and continuous, and that wild contribute to the gene pool cultivated.
Moreover, it has been suggested that domestication occurred, or across the Mexico-Guatemala, or that there were two separate domestication events area: Guatemala-Honduras, and Mexico (Spataro et al., 2011). The domestication process seems to have eroded the levels of genetic variability of P. coccineus, as both were cultivated and wild maintain high variability in both populations of the Central Plateau as in Chiapas, in southern Mexico. The high variability is due to crossover rates ranging from intermediate to high in contrast to other species of bean (Escalante et al., 1994). These authors note that similar levels of genetic variation in wild and cultivated may also be due to: high levels of gene flow between populations; large effective populations generated by pollinators (bees and hummingbirds) over long distances; because wild subpopulations grow together; because the cultivated grow near wild; or the survival of individuals that can reach up to 10 years (Fernández, 1981).
In addition, the genetic distance between populations of Central Mexico is lower (0.024-0.027 Nei index), that found between populations of Chiapas and Central Plateau (0.339-0.445) indicating the existence of geographic genetic differentiation. There have been no changes in the reproductive system of P. coccineus L. and perhaps it has not evolved to the self-reproduction by: 1) slower rate between generations because the plant can reach 10 years; 2) the existence of little genetic variability in the reproductive system of P. coccineus L. and that the variation is more due to environmental effects, such as plants most at risk and most visited flowers have higher rates of crossover; and 3) the gene flow between wild and cultivated make the artificial selection to the self-reproduction is slow (Escalante et al.,1994;Ayala et al., 2006; Castillo et al., 2006).
This work was conducted in 2010 and 2011 crop seasons in the Valley of Mexico Experimental Field in Chapingo, State of Mexico, at an elevation of 2250 meters at the coordinates 19° 29' north latitude and 98° 53' west longitude. In Chapingo, the maximum annual average temperature is 25.1 °C, the average annual minimum is 7.6 °C and, the annual precipitation is 620 mm (http://smn.cna.gob.mx/climatologia/normales5110/normal15170.TXT). 16 variants of a runner bean collection from the Ejido San Jose de las Corrientes, Vicente Guerrero, Durango, at an elevation of 2120 meters, and coordinates 23° 46' 07" north latitude and 103° 51' 30" west longitude. In this locality, the maximum annual average temperature is 26.2 °C, the average annual minimum 7.5 °C and total annual rainfall of about 500 mm (http://smn.cna.gob.mx/climatologia/Normales5110/normal10088.TXT). The variants were selected by the colour of testa of seeds contained in the collected population with different color of testa, 6 and 12 single color combinations. Plain colors were white, light, cream, yellow mustard cream, lilac and color suede or leather.
The combined were: light brown to dark brown, purple with black, yellow with black, yellow with coffee, coffee with cream, black and cream, black with gray and cream, black with purple, cream with black and gray, and black coffee and yellow. Planting dates were in 2010; June 25 and 17 May 2011. Planting was done in a furrow plots 3 m long at a distance of 0.80 m between rows leaving a groove between each variant of “ayocote”. Each plot contained 5 plants (repetitions) of runner bean spaced 0.60 m between plants and three plants per hill.
Date and photoperiod were recorded at the beginning of flowering (first flower open on the ground), beginning of maturity (first ripe pod on the ground) as well as the length of the period of maturation of the pods. The calculation of photoperiod was obtained from the website: http://www.tutiempo.net/silvia_larocca/Temas/fotoperiodo.htm Lammi (2014). Using data from daily minimum and maximum temperatures obtained from the meteorological station of Chapingo (UACH) accumulated temperature in the growing season and reproductive period of each plant or recurrence was determined.
The analysis of variance that included 16 variants, two sowing dates and 5 repetitions showed significant difference between planting dates and among variants for days to flowering, light hours and accumulated temperature emergence to flowering. Likewise for cumulative days elapsed and temperature from the start of the start of flowering to maturity. The variability included within this collection not only reflected in the external features of seed as mottled colours and patterns of the testa, but also in plant phenological characteristics (Table 1).
Table 1 Analysis of variance of phenological variables and mean temperature accumulated in two periods of growth of runner bean plant. Average of two planting dates.
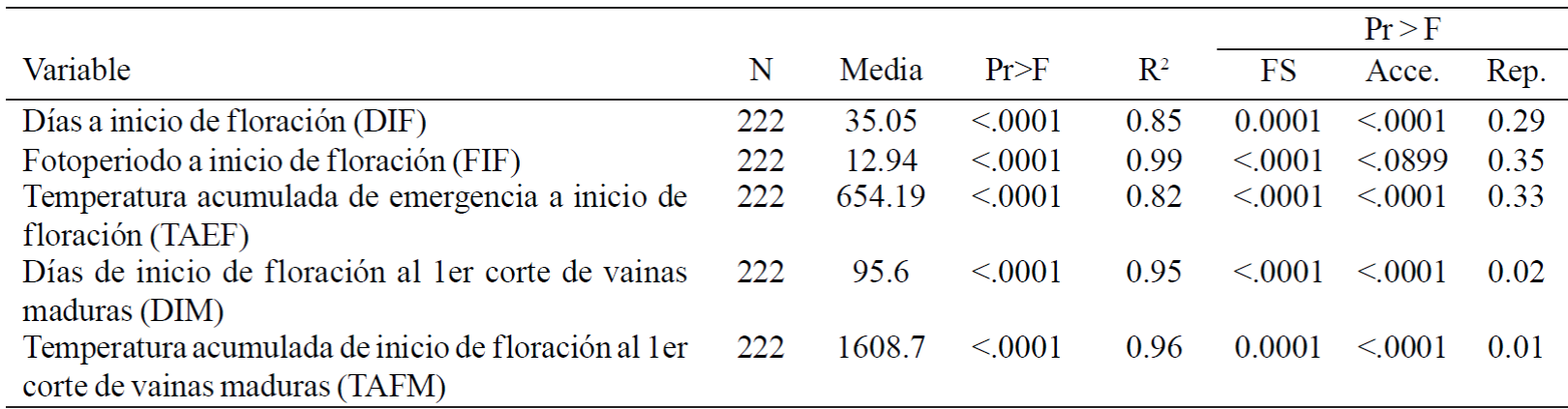
N= número de observaciones.
With the data obtained in 2011, variants of short cycle (123 and 144 dae) and long cycle (164-165 dae) were identified. The short cycle flourished between 26 and 36 DAE, began maturity between 123 and 139 dae, and pod maturity period varied between 7 and 21 days. Long cycle variants flourished between 26 and 38 DAE, began maturity at 164-165 dde, and pod ripening period was 27-41 days.
Sowing 40 days before June 25, caused the plant to lengthen the growing season (DIF), which was reflected in an increase in accumulated temperature (TAEF) in that period. Also started blooming with less daylight hours (FIF) due to decreased natural photoperiod occurred over time (Figure 1). The occurrence of consecutive frost from 5 to 13 November 2010, with minimum temperatures ranging between 0 and -5.6 °C, forced to harvest all variants on November 8, 126 days after seedling emergence so no was possible to know the actual number of days needed to start plants mature pods (Figure 2 and Table 2).
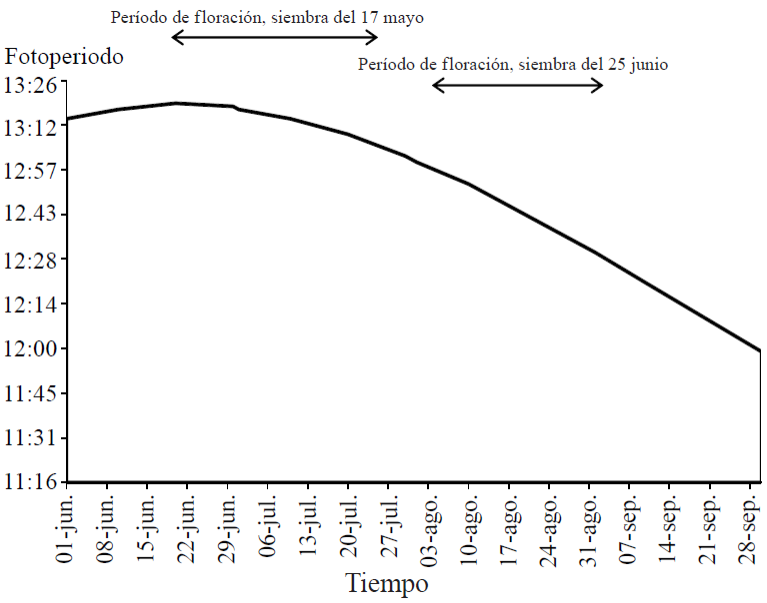

Table 2 Means and standard deviation of phenological variables and average temperature accumulated in two periods of plant growth of runner bean in two sowing dates.

FS= fecha de siembra; N= número de obseraciones; DT= desviación típica; DIF= días a inicio de floración; FIF= fotoperíodo a inicio de floración; TAEF= temperatura acumulada de emergencia a floración; DIM= días a inicio de madurez; TAFM= temperatura acumulada de floración a madurez
Plant response to photoperiod and temperature
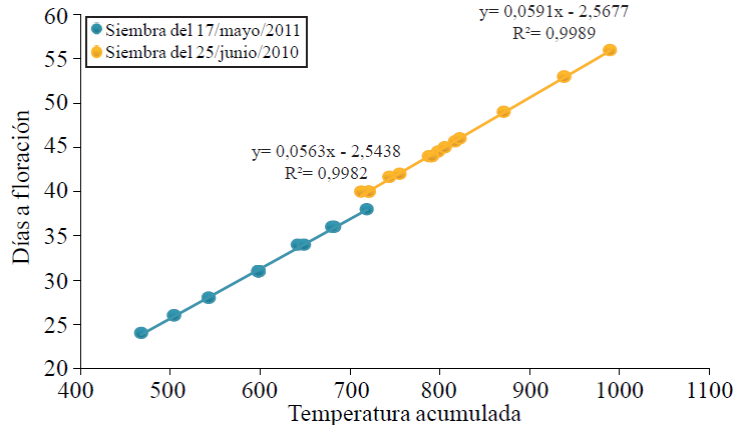
The temperature and photoperiod are the most important factors that induce plant development through its growing cycle. In both planting dates, the number of days from seedling emergence to the beginning of flowering, decreased as photoperiod is reached (Figure 3). The response of the plant to be planted in early May was the beginning of flowering rush (between 26 and 36 DAE) with long days between 13.17 and 13.19 hours light. While in June planting flowering plant began many days (between 40 and 56 days) with shorter photoperiods between 12.33 and 12.49 hours light (Figure 1).

Figure 3 Response in days to flowering to photoperiod of 16 variants of a collection of native patol grown in two planting dates in Chapingo, State of Mexico. Cycles spring-summer, 2010 and 2011.
Conclusions
In both planting dates, increasing the days of seedling emergence to flowering increased the accumulation of temperature in this period. By sowing May 17, while the plants grew and developed to initiate flowering accumulated between 468 and 719 °C of temperature, lower quantity than the accumulated over the same period to sow on June 25, which was 713-990 °C.
Literatura citada
Ayala, G. O. J.; Pichardo, G. J. M.; Estrada,G. J. A.; Carrillo, S. J. A. y Hernandez, L. A. 2006. Rendimiento y calidad de semilla de frijol ayocote en el Valle México. Agric. Téc. Méx. 32(3):313.321. [ Links ]
Búrquez, A. y Sarukhán, J. 1980. Biología floral de poblaciones silvestres de Phaseolus coccineus L. I. Relaciones planta-polinizador. Boletín Sociedad Botánica de México 39:5-25. [ Links ]
Castillo, M. M.; Ramírez, V. P.; Castillo, G. F. y Miranda, C. S. 2006. Diversidad morfologica de poblaciones nativas de frijol común y frijol ayocote del oriente del estado de México. Rev. Fitotec. Mex. 29(2):111-119. [ Links ]
Coello, C. J. G. y Escalante, G. A. M. 1989. Estructura genética y determinación de los parámetros del sistema de apareamiento en poblaciones silvestres y cultivadas de Phaseolus coccineus. Tesis de Biólogo. Facultad de Ciencias, Universidad Nacional Autónoma de México. 98 p. [ Links ]
Delgado-Salinas, A. 1988. Variation, taxonomy, and germplasm potentialities in Phaseolus coccineus . In: Gepts, P. (Ed.). Genetic resources of Phaseolus beans. Kluwer, Dordrecht. 441-463 pp. [ Links ]
Dobzhansky, T. 1951. Genetics and the origin of species. 3rd (Ed.). New York: Columbia University Press. [ Links ]
Escalante, A. M.; Coello, G.; Equiarte, L. E.; Pinero, D. 1994. Genetic structure and mating systems in wild and cultivated populations of Phaseolus coccineus and P. vulgaris (Fabaceae). Am. J. Bot. 81(9):1096-1103. [ Links ]
Fernández, P. 1981. Ciclos de vida comparativos de poblaciones de Phaseolus coccineus L. (Leguminosae). PhD. thesis, Universidad Nacional Autónoma de México (UNAM). D. F. México. [ Links ]
Lammi, J. 2014. Cálculo del fotoperiodo. http://www.tutiempo.net/silvia_larocca/Temas/fotoperiodo.htm. [ Links ]
Miranda, C. S. 1967. Infiltración genética entre Phaseolus coccineus L. y Phaseolus vulgaris L. Serie de Investigación Núm. 9. Colegio de Postgraduados en Ciencias Agrícolas, Chapingo, Estado de México. [ Links ]
Servicio Meteorológico Nacional. Normales Cllimatológicas. 2014. http://smn.cna.gob.mx/climatologia/Normales5110/NORMAL10088.TXT. [ Links ]
Servicio Meteorológico Nacional. Normales Cllimatológicas. 2014. http://smn.cna.gob.mx/climatologia/Normales5110/NORMAL15170.TXT. [ Links ]
Spataro, G.; Tiranti, B.; Arcaleni, P.; Bellucci, E.; Attene, G.; Papa, R.; Spagnoletti, Z. P. and Negri, V. 2011. Genetic diversity and structure of a worldwide collection of Phaseolus coccineus L. Theor. Appl. Genet. 122:1281-1291. [ Links ]
Received: February 01, 2015; Accepted: May 01, 2015
 Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
