











 text in
text in  English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkIntroducción
El mango (Mangifera indica L.) es uno de los frutales tropicales más importantes en el país, se cultiva en cerca de 185 mil hectáreas con una producción aproximada de 1.5 millones de toneladas (SIAP, 2011). México es el cuarto productor mundial, superado sólo por India, Tailandia y China. La exportación de este fruto se basa principalmente en los cultivares de Florida como Tommy Atkins, Haden, Kent y Keitt; en los últimos años ingresó a este grupo el cultivar Ataulfo, un material originario de México, el cual tiene gran aceptación en los mercados internacionales por presentar algunas ventajas como mayor vida de anaquel que la de los cultivares de Florida. El fruto del cv. Ataulfo tiene calidad de exportación, por lo que representa 13% del volumen de exportación anual (Vázquez-Valdivia et al., 2006).
Uno de los problemas más importantes que presenta el mango ʻAtaulfoʼ, es la presencia de una gran cantidad de frutos que quedan pequeños y no tienen valor de exportación; estos mangos son conocidos técnicamente como frutos sin semilla y por su tamaño los productores los llaman "niño"; dichos frutos carecen de semilla y muchos de ellos presentan una cuarteadura longitudinal, algunos frutos caen y otros quedan adheridos al árbol hasta el momento de la cosecha.
La presencia de este tipo de frutos es variable dentro de las huertas, en algunas de ellas es mínima, pero en otras puede alcanzar hasta el 80 % de los frutos (Pérez et al., 2007). Las causas de la formación de estos frutos y la forma de evitarlo, hasta el momento se desconocen. Singh (2005) observó reducción en el tamaño y peso del fruto por efecto del aborto del embrión en los cultivares Glenn, Irwin, Haden, Kent y Kensington. Los problemas en la polinización y fecundación conducen al aborto de embriones y a la formación de frutos pequeños sin semilla (Beasley et al., 1999), los cuales pueden caer en etapas tempranas del desarrollo o bien llegar hasta cosecha.
En relación con los aspectos nutrimentales, es bien conocido que el boro (B) es un elemento necesario para la producción y germinación del grano de polen y el crecimiento del tubo polínico (Lovatt y Dugger, 1984; Delgado et al., 1994; Marschner, 1995), juega un papel importante para la producción de semilla, ya que bajo deficiencia moderada o severa de este nutrimento, las plantas dejan de producir flores funcionales y pueden dejar de producir semilla (Gupta, 2007). En las flores, generalmente concentraciones bajas de B reducen la fertilidad masculina por efecto del deterioro de la microsporogénesis y afectación del crecimiento del tubo polínico. Los efectos pos-fecundación, incluyen el deterioro de la embriogénesis, lo que resulta en el aborto de la semilla o la formación de embriones incompletos o dañados y frutos malformados (Dell y Huang, 1997).
El B también es importante para el crecimiento del fruto, por su efecto en la división celular y proceso de expansión (Díaz, 2002); algunos cultivares de mango como Tommy Atkins, Haden 2H y Van Dyke son sensibles a la baja concentración de boro, presentando intensa caída de frutos jóvenes, aborto de semilla y por ende baja producción; cuando estos cultivares recibieron la aplicación de Borax al suelo a dosis de 300 g por planta en un primer año y de 100 g al año siguiente, todos incrementaron considerablemente la producción (Rossetto et al., 2000). Xue-Qun et al. (2011) observaron que con la aspersión de borato (K2B4O7) a dosis de 15 mM, a árboles de mango en floración, disminuyó significativamente el porcentaje de flores con antracnosis (Colletotrichum gloeosporioides) y se incrementó el número de frutos por rama.
Por lo anterior, este estudio se planteó con los objetivos de: a) determinar si la aplicación de B al suelo puede disminuir la formación de frutos sin semilla; b) saber si el B aplicado mejora el contenido de macronutrimentos y B en hojas, flores, frutos con semilla y sin semilla; c) conocer la distribución de los macronutrimentos y del B en el fruto; y d) comparar la concentración nutrimental entre frutos con semilla y sin semilla.
Materiales y métodos
Este estudio se realizó en una huerta de mango ʻAtaulfoʼ con presencia de frutos sin semilla, ubicada en la localidad de Atonalisco, en Tepic, Nayarit, México; la cual se encuentra entre las coordenadas geográficas 21° 37' de latitud norte y 104° 48' de longitud oeste a una altitud de 601 m. El clima de la región es Aw2(w)(i') que corresponde a un cálido subhúmedo con lluvias en verano (García, 1973). Las características del suelo (0 a 40 cm de profundidad) fueron: pH de 6.0, conductividad eléctrica de 0.4 dS m-1, capacidad de intercambio catiónico de 12 cmolc kg-1, contenido de materia orgánica de 3.5%, textura franco-arcillo-arenoso y pobre (Castellanos et al., 2000) en contenido nutrimental (N 4.9, P 6.7, K 184, Ca 423, Mg 183 y B 0.27 mg kg-1). El huerto tenía una densidad de 156 árboles ha-1 de 8 años de edad (en marco real de 8 m), injertados sobre mango criollo de la región. Se realizaron dos aplicaciones (al inicio y finales de la temporada de lluvias) de 3 kg por árbol de la fórmula de fertilización 17-17-17.
El experimento consistió en la aplicación de tres dosis de boro al suelo: 25, 50 y 100 g por árbol. La fuente comercial utilizada fue Solubor (20.5% de B). Para lo cual se seleccionaron al azar 5 árboles para cada tratamiento, incluyendo un tratamiento control (sin fertilización con B). La aplicación se hizo a los tres meses después de la cosecha, en forma equitativa en los cuatro puntos cardinales de la copa y en la zona de goteo del árbol. El diseño experimental fue completamente al azar, con cinco repeticiones, la unidad experimental fue un árbol.
Antes de la aplicación del boro se hizo un muestreo foliar en los brotes del flujo vegetativo de verano, con la finalidad de conocer el estado nutrimental inicial de los árboles; para evaluar el efecto de los tratamientos durante la floración se hizo un muestreo de hojas de brotes con inflorescencias y de brotes sin inflorescencias, así como de inflorescencias; finalmente, dos semanas antes de la cosecha se realizaron muestreos de hojas, frutos sin semilla y frutos con semilla. Para los muestreos foliares se recolectaron 40 hojas completas (lámina con pecíolo) sanas y maduras por árbol de la parte media de los brotes de alrededor de la copa, semejante a como lo realizó Oosthuyse (2000).
Se muestrearon cuatro inflorescencias por árbol (una por punto cardinal de la copa) y de cada inflorescencia se recolectaron 6 ejes secundarios (2 basales, 2 intermedios y 2 apicales). Para los muestreos de frutos, se seleccionaron los que se encontraban en madurez fisiológica (epidermis color verde amarillento claro) de los cuatro puntos cardinales de la copa; se recolectaron 8 frutos sin semilla y 4 frutos con semilla (también llamados normales) por árbol. En el laboratorio los frutos normales se seccionaron en pulpa con epidermis y semilla, para realizarles el análisis nutrimental por separado.
Se evaluó la concentración de N, P, K, Ca, Mg y B en las muestras de cada órgano; para lo cual se usaron 0.5 g de muestra seca, que se sometieron a una digestión húmeda con una mezcla diácida (H2SO4:HClO4, 2:1 v/v) y peróxido de hidrógeno. La determinación de N se hizo por el método microkjeldahl, el resto de elementos se determinó en un espectrofotómetro de emisión atómica de plasma por inducción acoplada (ICP) AES de Varian Australia Pty Ltd (Victoria, Australia) en todos los casos se siguió la metodología descrita por Alcántar y Sandoval (1999). Por último, se calculó el porcentaje de frutos con y sin semilla por árbol del total de frutos por tratamiento.
Para analizar los resultados se realizó un análisis de varianza y la prueba de comparación de medias de Tukey con el paquete SAS versión 9.0 (SAS Institute, 1999).
Resultados y discusión
Concentración nutrimental. En el Cuadro 1 se presentan las concentraciones de los elementos evaluados en las hojas antes de la aplicación del boro. De acuerdo con Agustí (2004) la concentración de suficiencia de N foliar en mango está entre 1 y 1.5%, lo que implica que en los árboles de ʻAtaulfoʼ utilizados en esta investigación la concentración de N fue baja, lo cual tiene relación con el muy bajo contenido de este elemento en el suelo de la huerta como lo demostró el análisis realizado (4.9 mg kg-1); en tanto que las concentraciones foliares del resto de los elementos, a pesar de que se encontraban en bajas concentraciones, según la interpretación de análisis de suelo de Castellanos et al. (2000), se encontraron dentro de los intervalos de suficiencia.

El N es uno de los principales elementos estructurales de las plantas y juega un papel muy importante en el crecimiento vegetativo y reproductivo Zong-min et al. (2012), este elemento representa 2% del total de la materia seca de las plantas y es un componente de proteínas, ácidos nucleicos, coenzimas y numerosos metabolitos secundarios (Miller y Cramer, 2004). Su insuficiencia reduce el crecimiento de la planta y del área foliar con la consecuente disminución en la tasa fotosintética. El manejo adecuado del N en las plantas hortícolas conduce a obtener alta calidad de flores, frutos y semillas (Zong-min et al., 2012).
En las especies frutales la deficiencia de nitrógeno puede inducir el aborto del polen; en condiciones de abasto adecuado el N mejora la longevidad del óvulo (Díaz, 2002), lo que conlleva a mejorar el amarre de fruto. En mango se ha observado que el mal manejo del suministro de N al árbol afecta la producción de flores, fructificación y calidad del fruto; el exceso de este elemento en el suelo promueve un crecimiento vegetativo vigoroso en detrimento del reproductivo (Medeiros et al., 2004).
Al analizar el efecto de los tratamientos sobre la concentración nutrimental en las hojas, se observó que la aplicación de B sólo influyó en la concentración de Ca y B; la mayor concentración de Ca fue obtenida con la dosis de 100 g de B; se pudo observar que las tres dosis de boro al suelo mejoraron la concentración de B en la planta (Cuadro 2). Esto significa que el B aplicado fue absorbido y transportado vía xilema a la copa del árbol, con mejora en la absorción del Ca, el cual se encontraba en muy baja concentración (Castellanos et al. , 2000) en el suelo de la huerta (423 mg kg-1 de suelo).
Cuadro 2 Concentración nutrimental en hojas de mango ʻAtaulfoʼ con aplicaciones de boro al suelo en Nayarit, México.
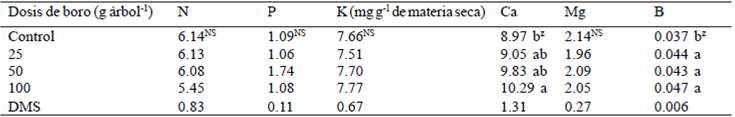
zMedias con la misma letra dentro de columnas no son diferentes de acuerdo a la prueba de medias de Tukey (p≤ 0.05). DMS= diferencia mínima significativa. NS= no significativo.
Fageria (2001) señala que el B ayuda en la traslocación del Ca absorbido hacia la parte aérea de las plantas. Sin embargo, esto no coincide con Marschner (1995) y Castellanos et al. (2000), quienes señalan que a nivel de suelo y de reacciones en la superficie de la raíz el B reduce la utilización de Ca. Sin embargo, ninguno de los tratamientos con B mejoraron la concentración foliar de N, lo cual no coincide con Marschner (1995), quien indica que el B puede tener un efecto indirecto favorable sobre la absorción y asimilación del nitrato, debido a que induce a un incremento en la actividad de la nitrato reductasa, como lo observaron Bellaloui et al. (2010) en soya [Glycine max (L.) Merr.].
Los tratamientos aplicados sólo tuvieron efecto en la concentración de B en las hojas de brotes con inflorescencias y en las hojas recolectadas previo a la cosecha de los frutos. En ambos tipos de hojas la concentración de 100 g de B al suelo duplicó la concentración de boro, comparada con la concentración de las hojas del control (Cuadro 3). Esto quiere decir que el B se transportó a ese tipo de hojas debido a que había puntos de demanda del elemento, como son las flores y frutos en crecimiento. En árboles de olivo Delgado et al. (1994) observaron mayor concentración de B en hojas jóvenes de árboles en floración que en las de árboles que no estaban en floración, lo que indica una necesidad mayor de B por los árboles durante el proceso reproductivo.
Cuadro 3 Concentración de boro en hojas de mango ʻAtaulfoʼ con aplicaciones de boro al suelo en Nayarit, México.

zMedias con la misma letra dentro de columnas no son diferentes de acuerdo a la prueba de medias de Tukey (p≤ 0.05). DMS= diferencia mínima significativa.
La concentración de N foliar inicial (antes de la aplicación de tratamientos), disminuyó 50% durante la floración, como lo indican las concentraciones en las hojas de los brotes vegetativos y reproductivos; la concentración en las inflorescencias fue igual a la de las hojas antes de aplicar los tratamientos (inicial) y a la de las hojas antes de la cosecha; los frutos normales fueron los que presentaron la mayor concentración de N, los frutos sin semilla presentaron la misma concentración que las hojas de brotes vegetativos y florales (Cuadro 4). La mayor concentración de P y K se observó en los frutos normales, seguida por la de las inflorescencias; la concentración de P de las hojas de las diferentes etapas y la de los frutos sin semilla fue semejante; la de K de las hojas de todos los muestreos fue similar; en tanto que la de los frutos sin semilla fue 50% menor a la de los frutos normales.
Cuadro 4 Concentración nutrimental (mg g-1 de materia seca) en hojas, inflorescencias y frutos de mango ʻAtaulfoʼ en Nayarit, México.

zMedias con la misma letra dentro de columnas no son diferentes de acuerdo a la prueba de medias de Tukey (p≤ 0.05). DMS= diferencia mínima significativa.
La concentración foliar inicial de Ca y la de las hojas al momento de la cosecha de los frutos fueron similares entre sí, y superiores a la del resto de los órganos; la menor concentración de este elemento se registró en las inflorescencias y en los dos tipos de frutos. La mayor concentración de Mg se observó en las hojas de los brotes vegetativos y en la inflorescencia, seguida por la de las hojas de los brotes florales y la de los frutos normales; la menor concentración la presentaron los frutos sin semilla. La concentración de B inicial fue la mayor y disminuyó en el resto de las estructuras evaluadas, incluyendo a los frutos normales; los frutos sin semilla presentaron la menor concentración de este elemento (Cuadro 4).
Estos resultados indican que el N, P, K y Mg fueron transportados a los puntos de demanda, como lo fueron las inflorescencias y los frutos normales; ya que sus concentraciones fueron iguales o superaron a la concentración foliar inicial (Cuadro 4), debido a que todos ellos son elementos móviles en floema (Marschner, 1995). En el caso del Ca y B, los resultados indican que ambos elementos se concentraron en las hojas, principalmente antes de la floración (valor inicial); en tanto que las inflorescencias y frutos (puntos de demanda) de los dos tipos recibieron muy poco aporte; esto se debe a que el transporte de ellos es por el xilema y depende de la transpiración del órgano, y las hojas son las estructuras con mayor tasa transpiratoria; además de que el Ca y el B son considerados de muy baja movilidad en el floema (Epstein y Bloom, 2005; Fageria, 2009), de tal forma que la mayoría del Ca y B demandados por los puntos en crecimiento, como ápices de brotes, hojas jóvenes o frutos tienen que ser cubiertos por el transporte vía xilema (Marschner, 1995). Medeiros et al. (2004) al evaluar el contenido nutrimental en hojas en cuatro etapas fenológicas del cultivar Tommy Atkins, encontraron que el Ca fue el nutrimento de mayor concentración en las hojas, no así en las flores y frutos.
Porcentaje de frutos con y sin semilla. Las dosis de 50 g de B por árbol disminuyó a la mitad el porcentaje de frutos sin semilla con respecto al control; en tanto que la dosis de 100 g presentó un 35% menos de estos frutos (Cuadro 5). Esto se debe a que el B es un micronutrimento esencial para la producción de frutos con semilla. En las flores, la concentración baja de B reduce la fertilidad masculina por deterioro de la microsporogénesis y crecimiento del tubo polínico al promover la formación de calosa en el estilo. Los efectos pos-fecundación incluyen el deterioro de la embriogénesis, lo que resulta en el aborto de la semilla o la formación de embriones incompletos o dañados y frutos malformados (Dell y Huang, 1997; Díaz, 2002).
Cuadro 5 Porcentaje de frutos sin semilla y con semilla en árboles en mango ʻAtaulfoʼ con aplicaciones de boro al suelo en Nayarit, México.

zMedias con la misma letra dentro de columnas no son diferentes de acuerdo a la prueba de medias de Tukey (p≤ 0.05). DMS= diferencia mínima significativa.
En flores de vid (Vitis vinifera L.), cuyos estigmas contenían de 8 a 20 μg de B por g de materia seca tuvieron pobre fecundación, en comparación con aquellas donde el estigma contenía de 50 a 60 μg de B (Díaz, 2002). Rossetto et al. (2000) observaron que los cultivares de mango tienen diferente comportamiento en relación al nivel de boro en la hoja. El mango ʻWinterʼ presentó buena producción y retención de frutos normales bajo condiciones de concentración baja de boro en las hojas (10 mg kg-1 de ms); mientras que los mangos ʻHaden 2Hʼ y ʻVan Dykeʼ fueron sensitivos a esa concentración, y en consecuencia, presentaron caída intensiva de frutos jóvenes, aborto de semilla y producción baja.
Con los resultados obtenidos en este estudio en mango ʻAtaulfoʼ, las concentraciones de B kg-1 de ms. corresponden a 60 mg en las hojas antes de la floración (valor inicial); en floración, 41 mg en las hojas de brotes sin inflorescencias y 48 mg en hojas de brotes con inflorescencias (Cuadro 4). Con el tratamiento de 100 mg de B, la concentración foliar de brotes productivos se elevó a 63 mg kg-1 de ms (Cuadro 3); lo que indica que en ʻAtaulfoʼ las flores requieren de más B que en otros cultivares y que con las dosis de 50 y 100 g de B se alcanzaron concentraciones en los brotes productivos que pudieron evitar el aborto del embrión.
Concentración nutrimental en frutos. La concentración de N, P y Mg fue significativamente mayor en los frutos con semilla que en los frutos sin semilla. La concentración de K fue similar en ambos tipos de frutos y la de Ca fue superior en los frutos sin semilla. En los frutos con semilla, ésta fue la estructura que concentró más N, P, K, Ca y Mg que la pulpa con epidermis. La concentración de B fue similar entre los dos tipos de frutos y entre la semilla y la pulpa con epidermis de los frutos normales (Figura 1).
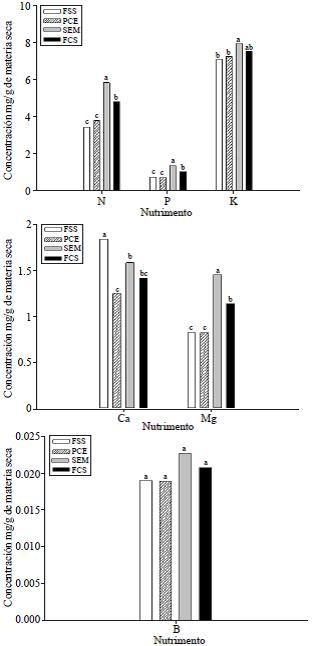
Figura 1 Concentración nutrimental en fruto sin semilla (FSS), pulpa con epidermis (PCE), semilla (SEM) y en fruto con semilla completo (FCS). Medias con la misma letra son estadísticamente iguales de acuerdo a la prueba de medias de Tukey (p≤ 0.05).
La concentración de N, P, K, Mg y B de los frutos sin semilla fue similar a la encontrada en la pulpa con epidermis de los frutos normales; sólo la concentración de Ca en el fruto sin semilla superó a la de la semilla y pulpa con epidermis (1.2 mg) (Figura 1).
La distribución nutrimental en el fruto varía de acuerdo al cultivar y sobre todo entre la pulpa y epidermis, ya que en mango 'Manila' Guzmán et al. (1996) encontraron que la epidermis del fruto fue el tejido más rico en Mg, Fe y Mn, la pulpa en K, el hueso en Ca y la semilla en N, P, Cu y Zn; por lo que la extracción nutrimental del fruto fue en el orden siguiente: K>N>Ca>Mg>P. Por otra parte, Singh (2005) indica que no encontraron diferencias significativas entre las concentraciones de N, K y Mg en la epidermis y la pulpa, ni en las de P y Ca en pulpa de frutos sin semilla y frutos con semilla en cinco cultivares evaluados. En el caso del cv. Ataulfo, se observó que la semilla fue la que concentró más a todos los elementos evaluados en comparación con la pulpa con epidermis.
Como ya se mencionó, existe una correlación positiva alta entre la distribución de Ca y la tasa de transpiración de los órganos, lo que explica el contenido bajo de Ca (<0.3%) en los frutos (estructura con poca transpiración), comparada con la de las hojas (3 a 5%) en la misma planta (Marschner, 1995); por lo cual, en los frutos de mango ʻAtaulfoʼ se tuvo baja concentración de Ca y mucho más alta en las hojas. La distribución de B está relacionada también con la pérdida de agua del órgano, por lo que las hojas acumulan más boro que las semillas y tejidos del fruto, como también se observó en este estudio con mango ʻAtaulfoʼ (Cuadro 4). Aunque estudios en árboles de olivo (Olea europea L.) han mostrado que las flores y frutos, en sus primeros estados de desarrollo, pueden promover la movilización de B foliar para satisfacer su demanda por este elemento (Delgado et al., 1994).
En el caso de los frutos sin semilla, la ausencia del embrión en la semilla, hace que la demanda por los elementos disminuya considerablemente, y por lo tanto se concentren menos. La presencia de semillas representa un factor importante en el crecimiento de los frutos, ya que es el órgano regulador del desarrollo de los mismos (Díaz, 2002) y afecta la acumulación de nutrimentos, particularmente la de Ca en pulpa y epidermis (Singh, 2005). Durante el desarrollo de las semillas, el embrión produce fitohormonas como las auxinas y giberelinas, que estimulan el desarrollo de la semilla y de los tejidos del fruto, como la pulpa (Díaz, 2002) ejerciendo una fuerte demanda nutrimental. El orden de prioridad entre demandas de un árbol en la distribución de fotosintatos y nutrimentos, lo determina la tasa de crecimiento (actividad de la demanda) y el tamaño de las demandas; en este contexto, la semilla tiene mayor fuerza de demanda que las partes del fruto, brotes en crecimiento y tejidos de almacenamiento del tallo (Wolstenholme, 1990). La mayor concentración de Ca registrada en los frutos sin semilla de ʻAtaulfoʼ se debe a que es un fruto de menor tamaño que el fruto con semilla y por ello el Ca se concentró más.
Conclusiones
La aplicación de B al suelo en dosis de 50 y 100 g disminuyó la formación de frutos sin semilla. Los árboles de mango ʻAtaulfoʼ usados en este trabajo, presentaron deficiencia de N y habrá que valorar si esto influye en la formación de frutos sin semilla. La aplicación de 100 g de B mejoró únicamente la concentración de Ca y B en la planta, y sólo mejoró la concentración de B en las hojas de brotes florales y en las del final del desarrollo del fruto. La distribución de los nutrimentos en el fruto normal fue la siguiente: la semilla acumuló más N, P, K Ca y Mg que la pulpa con epidermis y ambas estructuras tuvieron la misma concentración de B. Los frutos con semilla superaron a los sin semilla en la concentración de N, P y Mg; en tanto que éstos últimos concentraron más Ca, y ambos tipos de frutos tuvieron la misma concentración de K y B.

