










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkAgricultura, sociedad y desarrollo
Print version ISSN 1870-5472
agric. soc. desarro vol.2 n.1 Texcoco Jan./Jun. 2005
¿Fueron manipulados los datos de Mendel? (Argumentos en contra)
Were Mendel's data manipulated? (counterarguments)
Fidel Márquez-Sánchez 1 y Jaime Sahagún-Castellanos 2
1 Centro Regional Universitario de Occidente. Universidad Autónoma Chapingo. Rosario Castellanos. 2332. Colonia Residencial la Cruz. 44950. Guadalajara, Jalisco. (fidelmqz@hotmail.com).
2 Departamento de Fitotecnia. Universidad Autónoma Chapingo. 56230. Chapingo, Estado de México. (jsahagun@correo.chapingo.mx)
Resumen
Se discuten las opiniones de varios autores sobre la crítica de R. A. Fisher al ajuste de los datos de Mendel a sus hipótesis sobre la herencia en varios caracteres del chícharo (Pisum sativum). Datos demasiado buenos para ser ciertos, se ha argumentado. Tres autores basan su alegato en la forma inadecuada en que Fisher empleó la prueba de ji-cuadrada para sugerir que Mendel falseó sus datos experimentales. Otro autor señala cómo debe interpretarse el uso de la ji-cuadrada, ya que cuando se usa como lo hizo Fisher, para detectar datos falsificados, cualesquier conjunto de datos aceptables, congruentes con la hipótesis, parecerán demasiado buenos cuando se combinan.
Palabras clave: Datos manipulados, Mendel.
Abstract
The opinions of several authors on R. A. Fisher's criticism to the fit of Mendel's data to his hypotheses on the heredity of several character of peas (Pisum sativum) are discussed. These data are too good to be true, it has been argued. Three authors base their line of arguments on the inadequate way in which Fisher used the chi square test to suggest that Mendel lalsilied his experimental data. Another author indicates how the chi square should be interpreted; since if it is used, as Fisher did, in order to detect falsified data, any set of acceptable data, if congruent with the hypothesis, will appear to be too good when combined.
Key words: Manipulated data, Mendel.
Introducción
Este ensayo no puede tener las divisiones tradicionales de los escritos científicos o tecnológicos; 'no es un trabajo de investigación. Se trata de presentar a los lectores de Agricultura, Sociedad y Desarrollo, varios artículos que han aparecido acerca de los trabajos de Mendel, hacer algunos comentarios y concluir que los datos de Mendel no fueron manipulados.
El razonamiento de Fisher
Al análisis estadístico que R. A. Fisher hizo de los datos obtenidos por Mendel, que lo llevó a concluir que la probabilidad de que éstos puedan obtenerse en una nueva serie de experimentos idénticos de acuerdo con la ji-cuadrada de 7 en 100 000, se le ha llamado la paradoja de los datos demasiado buenos para ser ciertos (Pilgrim, 1984). Es decir, los datos se ajustaban tan bien a la hipótesis nula, que se podía dudar que fueran ciertos. Otros dos genetistas coincidieron con la argumentación de Fisher.
En genética, la ji-cuadrada se usa para probar la hipótesis de que los resultados de los diferentes tipos de individuos que resultan en una progenie se ajustan a una relación (3:1, 9:3:3:1, etc). El valor calculado de la ji-cuadrada (X2c) es función del número observado de individuos del tipo i (Oi), y de los números esperados (Ei), bajo el supuesto de que la relación hipotética es cierta. La estadística de prueba es
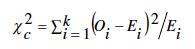
donde k es el número de tipos que pueden aparecer en la progenie. Entre más pequeño sea el valor de X2c más evidencia habrá de que la hipótesis es cierta, y la probabilidad (p) de no rechazar indebidamente la hipótesis nula será mayor.
¿Era empírico el enfoque de Mendel?
Antes de analizar los argumentos de los defensores de Mendel, conviene revisar lo que sus partidarios y antagonistas suponían respecto a su actitud ante la herencia. Monaghan y Corcos (1985a) creen que Fisher (1936), Dunn (1965) y Orel (1971), conocedores del trabajo de Mendel, podían pensar que éste era un teórico de la herencia, es decir, que sabía de antemano los resultados experimentales que obtendría, por lo que los datos podrían ajustarse acercándolos más a lo esperado. (A este respecto se piensa que quien ajustó los datos fue un ayudante de Mendel). Por el contrario Olby (1949) dice que Mendel tenía más interés en estudiar el comportamiento de los híbridos, y que ésto no tenía que ver con la naturaleza de la herencia. En este contexto Mendel habría sido un empírico, al ver primero qué obtenía de su experimentación y después formular alguna teoría. Sobre ésto, Glass (1963) señala que "...era bastante claro que en el caso Mendel, fue la existencia del inalcanzable e insoluble problema hasta ese entonces, lo que lo encaminó a idear los experimentos que diseñó, y no ninguna construcción conceptual (teoría) con la que se hubiera comprometido o hubiera deseado apoyar", es decir, que no hubiera teorizado al respecto. Sin embargo esto no significa que no tuviera idea, al observar sus resultados, de lo qué estaba sucediendo. Como Mendel mismo señaló, él iba encontrando evidencia de algunas regularidades consistentes. Así, en la introducción de su artículo (Mendel, 1866, traducido en Stern y Sherwood, 1966) establece que el propósito de sus experimentos era seguir el desarrollo de la progenie de los híbridos, ya que no se había formulado una ley aplicable a la formación y desarrollo de éstos. Monaghan y Corcos (1985a) añaden que la pregunta clave, bastante probable, era si los híbridos producirían progenies como éstos (to breed true) o no. Siguiendo en su artículo una amplia presentación y discusión sobre la naturaleza anual estacional de los experimentos de Mendel, sobre la notación empleada en la representación de los genes, dichos autores terminan dándole todo el crédito a Mendel, es decir, que éste no tenía una teoría preconcebida sobre la naturaleza de la herencia para poder ajustar sus datos, sino que simplemente era un investigador empírico sobre los híbridos y su descendencia.
Los defensores de Mendel
En un trabajo posterior, Monaghan y Corcos (1985b) discuten la naturaleza estadística de los resultados de Mendel, que fueron luego analizados por Fisher. Si los datos de Mendel estuvieran manipulados los valores de ji-cuadrada mostrados por Mendel tenderían a ser bajos; ellos muestran datos de siete estudios (dos en la generación F2 y cinco en la generación F3. En ésta, una combinación de los dos caracteres en F2 y cinco caracteres individuales más) y, aunque para el total la ji -cuadrada para la relación fenotípica 3:1 tiene un alto valor con probabilidades (todos los valores probabilísticos que se asientan en lo sucesivo se refieren al nivel observado de significancia de la estadística de prueba, suponiendo que la hipótesis nula es cierta) entre 0.95 y 0.99 (lo que asombró a Fisher), examinando los resultados en cada carácter no encuentran la tendencia a que fueran más bajos. El valor más bajo de ji-cuadrada fue 0.015 (0.90<p<0.95), y el más alto 0.606 (0.30<p<0.50). Al final de los dos experimentos en F2 parece que Mendel ya tenía un conocimiento significativo de que se obtenía la relación 3:1, por lo que los autores de ese artículo señalan que Mendel quería saber si las plantas permanecían constantes en su progenie (to breed true), lo que reafirmaría la idea del estudio de los híbridos que Mendel deseaba hacer. Por ello autofecundó las plantas y estudió la relación genotípica de los progenitores que exhibían el carácter dominante la cual, como sabemos ahora, sería 2:1, es decir, dos tercios de las plantas segregarían 3:1 y el resto no. Los datos de este experimento muestran ji-cuadradas que varían al azar, con el valor más bajo de 0.00 (p>0.99) y el más alto de 2.01 (0.1 0<p<0.20); por esta razón, de nuevo, sus resultados fenotípicos producen una probabilidad alta, entre 0.90 y 0.95.
En los experimentos con dos caracteres, (forma y color del cotiledón), Mendel nunca mencionó la ahora famosa relación 9:3:3:1 (Monaghan y Corcos, 1985a), ya que lo único que quería saber era la estructura genética de las plantas F2 de sus híbridos. Lo mismo sucede con el estudio para tres caracteres, donde obtiene una probabilidad entre 0.80 y 0.90. En el siguiente estudio Mendel retrocruzó el híbrido doble con el probador recesivo directa y recíprocamente, obteniendo la relación 1/4:1/4:1/4:1/4. La probabilidad de la ji-cuadrada del primer experimento (AaBb X aabb) fue alta (0.80<p<0.90), y la de su cruza recíproca (aabb X AaBb) también (0.90<p<0.95), por lo que tal vez, consciente o inconscientemente, Mendel cometió algún sesgo, pero continuó con sus trabajos para confirmar su hipótesis. Por esta razón sembró la siguiente generación de sus cruzas del dihíbrido con el probador doble dominante (cruza directa y recíproca) obteniendo 0.90<p<0.95 y 0.80<p<0.85. En experimentos posteriores añadió dos caracteres más, el color de la flor y la altura del tallo. Con todos los estudios: para uno, dos y tres caracteres, la probabilidad conjunta fue de 0.95 a 0.99, lo que nuevamente es una probabilidad muy alta, como en el caso de la relación fenotípica para un carácter. Con objeto de probar que pudo haber un sesgo inconsciente para el estudio de dos caracteres (forma y color del cotiledón), los autores compararon los resultados de Mendel con los de otros siete investigadores; encontraron que el porcentaje de recesivos era muy parecido al hallado por Mendel, lo que indica que el problema de mala clasificación no era sólo de éste, y por otra parte, que en el caso de dos investigadores sus resultados probabilísticos diferían bastante de los hallados por los otros.
Para reforzar la defensa de los resultados de Mendel, Corcos y Monaghan (1985) introdujeron otros dos argumentos: el de la reducción del número de plantas usadas en la F3 y el cálculo de Fisher de la homocigosis lograda en esta generación. En la generación F2 Mendel usó alrededor de 500 plantas; sin embargo, en la F3 sólo usó 100, sembrando 10 semillas por planta. Ésto fue porque Mendel ya presumía que la relación en F2 era 3:1, por lo que no consideró necesario tener más plantas. Fisher señalaba que tal reducción causaba que Mendel no obtuviera la relación de heterocigotes a recesivos 2:1, sino 1.8874:1.1126 (relación que se obtiene con base en la probabilidad de que un caracter sea dominante, calculada con la distribución binomial). Por tanto, Corcos y Monaghan calcularon los respectivos números de homocigotes de los cinco experimentos mencionados, según las relaciones propuestas por Mendel y Fisher. De acuerdo con los resultados de este último, los datos de Mendel, según la relación 2:1 se ajustan más a lo esperado, pero no sus resultados más sofisticados, basados en la relación 1.8874:1.1126. De acuerdo con la hipótesis de Fisher los valores de ji-cuadrada variaron de 0.03 a 3.47, para Mendel de 0.00 a 2.01, y para Fisher variaron al azar tanto como se esperaba de la hipótesis de Mendel y, en algunos casos, fueron tan bajos como sus resultados.
En esta serie de estudios no es extraño que Monaghan y Corcos (1985a) concluyan su artículo con la siguiente aseveración de Galileo: "Pero he logrado probar éste y otro hechos, no pocos, ni menos relevantes: y lo que considero más importante, que se ha abierto a esta vasta y más excelente ciencia, de la cual mi trabajo es sólo el inicio, modos y medios con los cuales otras mentes más agudas que la mía, explorarán sus confines más remotos''.
Pilgrim (1986), experto en el manejo e interpretación de datos experimentales, considera que la conclusión de Fisher (que los datos de Mendel eran falsos) es errónea.
Este autor cuestiona los métodos utilizados por Fisher, que luego fueron adoptados por otros autores. Monaghan y Corcos (1985a) concluyeron que a pesar de que tales métodos estadísticos fueron los apropiados, sus resultados (que coincidían con los de Fisher) no eran lógicos, particularmente los que se derivan de la combinación de información obtenida en varios experimentos independientes, no obstante que en lo individual los datos de cada experimento no mostraban sesgo alguno. Ante esta situación, lo que habría que investigar era si el método utilizado por Fisher, basado en la ji-cuadrada, era adecuado para detectar datos falsificados, y si los datos de Mendel habían sido manipulados. Pilgrim (1986) argumenta que el método que se basa en la ji-cuadrada sólo produce, sobresalientemente bien, la probabilidad de que los datos experimentales sean compatibles con la hipótesis, y que Fisher la usó para otro propósito: sabiendo que la hipótesis era correcta y que 105 datos se ajustaban a ella (puesto que consideraba que Mendel sabía lo que debía esperar), Fisher usó la ji-cuadrada para determinar si los datos de Mendel constituían una muestra aleatoria o estaban manipulados. También investigó la limpieza de los datos de Mendel; los métodos de Pilgrim, sin embargo, fueron muy diferentes: calculó la probabilidad exacta de que los datos obtenidos por Mendel fueran los obtenidos. El cálculo lo hizo con la distribución binomial con parámetros n (número de plantas) y p (probabilidad de la ocurrencia de un genotipo o fenotipo, determinado según la hipótesis particular). Así, para el caso de la proporción de dominantes heterocigóticos en la F2 de un monohíbrido, los parámetros fueron n=100 y p=proporción de los fenotipos dominantes genéticamente heterocigóticos (2/3). La media de esta distribución es np=100(2/3)=66.666 y la varianza es np(1-p) = 100 (2/3)(1/3)=100(2/9)=22.22, con una desviación estándar de 4.71. De acuerdo con el teorema de De Moivre-Laplace, aproximadamente dos tercios de todos los valores (cada valor es el número de heterocigóticos resultantes en una muestra aleatoria de tamaño 100) deberán estar en el intervalo 66.666+4.71; es decir, redondeando, en el intervalo [62, 72]. Los valores que obtuvo Mendel en los cinco experimentos en que usó 100 como tamaño de muestra fueron 64, 71, 60, 67 y 72, números muy congruentes con el razonamiento de Pilgrim. Para que los datos de Mendel hubieran sido demasiado buenos, afirma Pilgrim, los cinco valores deberían haber estado en el intervalo [65, 69] o quizá en el [66, 68]. Con los valores obtenidos, Pilgrim concluyó que no hay razón para sospechar que los resultados no corresponden a otra cosa que no sea una muestra aleatoria.
Para evaluar el resto de los datos de Mendel Pilgrim calculó la probabilidad (Pd) de que cada valor estuviera tan o más cercano a la media [100(2/3)] que en los experimentos de Mendel. Por ejemplo, para el valor 64, como la media es 66.667, Pd =P(X=64) + P(X=65) + P(X=66) + P(X=67) + P(X=68) + P(X=69) + P(X=70). De los tres primeros experimentos que involucraron la relación 3:1, en el primero [semilla redonda (R) vs semilla arrugada (A)] ocurrieron los valores 5 474(R) y 1 850(A) para los que Pd=0.378, lo que significa que en 37.8% de los casos los valores de un muestreo aleatorio hubieran estado tan o más cercanos de lo esperado que los de Mendel. Para los dos experimentos restantes Pd=0.113 y Pd=0.481. Estos resultados, del todo normales en un experimento de este tipo, comenta Pilgrim, difícilmente pueden considerarse extraordinarios. Para dar una idea más clara del significado de estas probabilidades, el autor presenta un ejemplo muy sencillo: para Pd=0.481 esta probabilidad es aproximadamente la que corresponde a la ocurrencia de dos hijos del mismo sexo en familias con dos hijos. La media de Pd en 14 experimentos de Mendel fue 0.487, lo que significa que 48.7% de las veces los resultados hubieran estado así de cerca o más a su media, o valor esperado. Con esta información Pilgrim tuvo que concluir que no había razón para creer que los datos de Mendel habían sido obtenidos deshonestamente. Una vez que concluyó que los datos no habían sido falseados, quedaba por determinar qué fue lo que hizo que así pareciera desde la óptica Fisheriana.
Para analizar qué tan apropiada fue la metodología utilizada por Fisher, Pilgrim, considerando que si bien los datos de Mendel no fueron ''demasiado buenos para ser ciertos'', se preguntó ¿por qué la ji-cuadrada usada por Fisher los mostró como tales? Usando esta distribución, sucede que si los datos son compatibles con la hipótesis las probabilidades altas (p) no son raras. También se preguntó Pilgrim ¿cuándo los datos aparecen como demasiado buenos? Al parecer Fisher nunca reflexionó sobre ésto; sin embargo, consideró que a medida que p era mayor los datos se hacían más sospechosos. Por lo anterior, Pilgrim creyó necesario obtener información que diera respuesta a estas preguntas. Para ello analizó con ji-cuadrada cuatro grupos de datos de cinco muestras de tamaño 100 cada uno. Los datos serían probados vs la hipótesis de que una mitad serían de un tipo y la otra mitad de otro tipo. Los grupos de datos fueron: 1) datos honestamente derivados (por muestreo aleatorio); 2) datos con una variación excesiva (descritos por Pilgrim como los de un investigador con mala suerte); 3) datos ostensiblemente cargados a un lado de la curva normal, 4) datos demasiado buenos. En el primer grupo obtuvo un valor de X2=2.32, con 5 grados de libertad, para una p=0.80; en el segundo X2=10 y p=0.5. Respecto a datos observados (Oi) ostensiblemente cargados a un lado de la normal (grupo 3), que no se ajustan a la hipótesis, la prueba no discrimina entre datos a un lado de la media (o valor esperado, Ei) o al otro; debido a que la estadística usada es 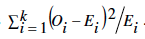 . En este caso los resultados fueron: X2=2.32 y p=0.8, ¡igual al grupo 1 de datos honestos! Finalmente, en el caso de datos demasiado buenos se obtuvo X2 = 0.32 con p>0.99.
. En este caso los resultados fueron: X2=2.32 y p=0.8, ¡igual al grupo 1 de datos honestos! Finalmente, en el caso de datos demasiado buenos se obtuvo X2 = 0.32 con p>0.99.
Para investigar qué pasa cuando se analiza conjuntamente los datos de varios experimentos, como lo hizo Fisher con los datos de Mendel, Pilgrim ideó varios escenarios. Con dos conjuntos de datos como los que hubieran sido obtenidos en muestras aleatorias (como los del grupo 1) se obtuvo: X2=4.62 y p=0.9. Con tres de estos conjuntos obtuvo un valor de X2=6.96 y p=0.96; es decir, cuando los datos buenos se repiten, es más probable que los datos confirmen la hipótesis. Pilgrim concluye que así es como se debe interpretar el uso de la ji-cuadrada, y que cuando se usa, como Fisher lo hizo, para detectar datos falsificados, cualquier conjunto de datos aceptable, si es congruente con la hipótesis, parecerá demasiado bueno cuando se combine.
Respecto a la crítica de Fisher de que los datos de Mendel están dentro de una desviación estándar, Pilgrim argumenta, con base en la aproximación de la binomial a la normal, que aproximadamente dos terceras partes de los datos deben estar dentro del error estándar. Él, como cualquier conocedor de la distribución normal, encuentra incomprensible la posición de Fisher, y concluye que la ji-cuadrada no es apropiada para detectar datos falsificados, y que no hay razón para que se cuestione la honestidad de Mendel. Pilgrim no fue el único que cuestionó los métodos estadísticos utilizados por Fisher para dictaminar sobre la honestidad de Mendel. Weilling (1986) concluyó que, como la varianza de las segregaciones genéticas es menor que la del tipo binomial supuesto, la ji-cuadrada no es una estadística adecuada para el propósito que persiguió Fisher. También concluyó que, como las varianzas de las diferentes segregaciones no son homogéneas, no se pueden combinar los datos de diferentes experimentos para ser analizados, como lo hizo Fisher. A continuación se presenta un resumen de los análisis de Weilling para obtener sus conclusiones. Con respecto a la magnitud de las varianzas de las segregaciones genéticas, consideró que en el ovario de una flor de la especie que utilizó Mendel para su estudio de híbridos debe haber cuando mucho 12 óvulos, pero no todos producen semilla. Si la hembra es de genotipo aa y el macho es Aa, y si el número de óvulos fertilizados fuera W, esta variable (números de cigotes resultantes) no seguiría una distribución binomial sino una hipergeométrica. La varianza de la Hipergeométrica es menor que la de la Binomial. Por esta razón concluyó que la distribución binomial que Fisher usó para determinar la relación 1.8874:1.1126, en lugar de la supuesta relación 2:1, no fue adecuada, y en consecuencia la crítica a Mendel tampoco lo fue. Esta relación 2:1 fue utilizada para determinar en la F2 de un monohíbrido la proporción de fenotipos dominantes cuya prole, generada por autofecundación, era homogénea o heterogénea.
También es interesante cómo Weilling (1986) analizó el supuesto de homogeneidad de varianzas requerido para el uso correcto de la ji-cuadrada para analizar datos provenientes de varios experimentos. Él consideró que en los caracteres de forma y color de semilla la segregación resultante se observa en las semillas que darán lugar a las plantas F2, mientras que en los de planta la segregación se observará en las que generen las semillas. Como en los procesos de germinación de semillas y desarrollo de plantas pueden ocurrir pérdidas, la varianza para estos caracteres debe ser mayor que para color y forma de semilla. Esto da como resultado que, por la heterogeneidad de varianzas, el uso de la ji-cuadrada para el análisis combinado de datos de diferentes experimentos, como el que hizo Fisher con los datos de Mendel, no sea apropiado.
Conclusiones
De acuerdo con la revisión de los datos de Mendel y de los argumentos que Fisher empleó para descalificar sus métodos, los autores que han opinado al respecto, cada uno con su particular aportación, llegan a la conclusión (sostenida por nosotros) de que los datos mendelianos fueron correctamente analizados. Vale la pena recalcar que en este respecto Fisher pudo estar equivocado.
Literatura Citada
Corcos, A., and F. Monaghan. 1985. More about Mendel's experiments: where is the bias? The Journal of Heredity 76: 384. [ Links ]
Dunn, L. C. 1965. Mendel, his work and his place in history. Proc. Am. Phil. Soc: 109:189-198. [ Links ]
Fisher, R. A. 1936. Has Mendel's work been rediscovered? Am. Science 1: 115-137. [ Links ]
Glass, B. T. 1963. The establishment of modern genetical theory as an example of the interaction of different models, techniques, and inferences. In: Scientific Change. A. C. Crombie, Ed. Basic Books Inc. Pt. 6, No. 17: 251-541. [ Links ]
Mendel, G. 1866. Versuch über Ptlanzen-Hybriden. Verhandlugen des Naturforschenden Vereines in Brünn 4(1865) Abhandlugen. pp: 3-47. [ Links ]
Monaghan, F., and A. Corcos. 1985a. Mendel, the empiricist. Heredity 76: 49-54. [ Links ]
Monaghan, F., and A. Corcos. 1985b. Chi square and Mendel's experiments: where's the bias? Heredity 76: 307-309. [ Links ]
Olby, R. 1949. Mendel no Mendelian. Hist. Science 17: 53-72. [ Links ]
Orel, V. A. 1971. A reconstruction of Mendel's Pissum experiments and an attempt at an explanation of Mendel's way of presentation. Folia Mendeliana no. 6. pp: 41-60. [ Links ]
Pilgrim, I. 1984. The too-good-to-be-true paradox and Gregor Mendel. The Journal of Heredity 75: 501-502. [ Links ]
Pilgrim, I. 1986. A solution to the too-good-to-be-true paradox and Gregor Mendel. The Journal of Heredity 77: 218-220. [ Links ]
Stern, C., and E. R. Sherwood. 1966. The Origins of Genetics: A Mendel Source Book. W. H. Freeman. [ Links ]
Weilling, F. 1986. What about R. A. Fisher's statement of the too-good data of Mendel' s pisum paper? The Journal of Heredity 77: 281-283. [ Links ]

