











 text new page (beta)
text new page (beta) English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkIntroduction
Since the first development of the island biogeography theory (MacArthur & Wilson, 1967), there have been numerous studies analysing the origins of the faunas of continental areas. In particular, isolated mountainous ranges and the so-called sky islands were attractive for biogeographers who assumed that the processes of immigration, as well as extinction and radiation taking place on “Darwinian islands” could well apply to these continental areas (Brown, 1971; Holland, 2009; Sanmartín et al., 2010; Vuilleumier, 1969). The island biogeography perspective was adopted readily on the Andean ranges in an attempt to explain the patterns of distribution and diversity among high elevation birds by dispersal from large source areas situated in the central Andes to the smaller isolated ranges in the northern Andes, and subsequent allopatric speciation (Adams, 1985; Vuilleumier, 1969; Vuilleumier & Monasterio, 1987).
According to the classical theory of island biogeography, the most important variable of the immigration / colonization process is the distance from the source area (MacArthur & Wilson, 1967). The effectiveness of a barrier is defined by the area unsuitable for free dispersal of individuals between hypothetical areas of origin and destination. In the case of terrestrial organisms established in oceanic islands such a barrier can be quantified as the distance over water separating them from a continent or another mainland. In the case of continental “islands”, however, there is no clear-cut barrier (Nekola & White, 1999). Lower elevation defined by valleys or flatlands separating 2 mountainous ranges might be a sort of barrier for montane species, but not totally impenetrable (Holland, 2009). Water barriers are generally constant over geological time, at least in the case of most oceanic islands of volcanic origin, whereas habitat barriers may appear and disappear over shorter periods.
In this study we evaluate the most important geographic variables in shaping the faunas ofthe Pronophilina butterflies in the northernmost Andean ranges, as the Northern Andes include the 3 cordilleras of Colombia (Bayona, 2018), being isolated by distance and topographic prominence (maximum elevation touch point). The concept of topographic prominence (called the key saddle) originated from the earth sciences (Choi et al., 2017; Llobera 2001), where it is applied in discriminating mountain peaks or hills. Topographic prominence characterizes the height of a mountain or hill’s summit as the relative height of a peak above the lowest contour that surrounds itself without encircling any higher peak (Choi et al., 2017; Rana, 2004). We also use this concept of topographic prominence for comparative purposes in this study.
This is a follow up of a preliminary biogeographical analysis carried out by Adams (1985), who compared the faunas of 3 Colombian cordilleras (Eastern, Central and Western), Cordillera de Mérida, Serranía de Valledupar (Sierra de Perijá) and Sierra Nevada de Santa Marta. Adams (1985) concluded about a positive species-area relationship (for land above 2,000 m asl) between relative isolation, species richness and percentage of endemic species. Adams (1985, p. 35.), however, did not speculate about the area of origin of the Pronophilina, nor did he compare the structure of the faunas of the above specified areas. Species-area correlation analysis brings very little information on regional or local biogeographical scale, and is of little relevance if other variables, such as the distance from the alleged source area, are not considered. Also, the gamma index of diversity applied by Adams (1985) is too coarse for regional studies dealing with the northern Andes, which are characterized by complex landscape. Area-independent diversity measures, such as species richness calculated along potential altitudinal transects, are considered to be more informative for assessing diversity at local and regional scales (Pyrcz, 2004; Pyrcz & Garlacz, 2012; Pyrcz, Wojtusiak et al., 2009).
The Neotropical subtribe Pronophilina (for some authors: tribe Pronophilini) within the cosmopolitan tribe Satyrini is a particularly suitable group for the studies of Andean biogeography. It is the most species rich group of Andean butterflies in the cloud forests and paramo habitats, with approximately 550 known species (Lamas et al., 2004). Adult Pronophilina are highly sedentary butterflies, similarly to most Satyrinae, particularly of Satyrini (Verovnik et al., 2013). They move little vertically and horizontally and keep near their host plants, mostly of the genus Chusquea spp. (Mahecha-Jiménez et al., 2011; Pyrcz, Casner et al., 2009).
Material and methods
Published distribution and taxonomic data are based on Adams & Bernard (1977) and Pyrcz (1999a, b) for the Sierra Nevada de Santa Marta; Adams & Bernard (1979), Pulido & Andrade (2007) and Viloria (1990) for the Sierra de Périja; Pyrcz & Viloria (2007) for El Tamá; Adams & Bernard (1981), Pyrcz & Viloria (2007, 2009) and Viloria (1994) for the Cordillera de Mérida; and Viloria et al. (2010) for the Cordillera de la Costa. Unpublished data come from C. Prieto, G. Rodríguez, J. Pulido, M. Costa, O. Mahecha, V. Díaz. Relationships between the species of Pronophilina have been inferred from previous phylogenetic studies using morphological and/or molecular data, in particular Casner & Pyrcz (2010), Pyrcz et al. (2010) and Pyrcz & Boyer (2011) for Lymanopoda Westwood, 1850; Pyrcz (2008) and Pyrcz & Viloria (2009) for Pedaliodes Butler, 1867; Pyrcz et al. (2013) for Corderopedaliodes Forster, 1964; Pyrcz et al. (2017) for Steromapedaliodes Forster, 1964; and Pyrcz, Lorenc-Brudecka, Zubek et al. (2018) for Arhuaco Adams & Bernard, 1977, as well on new original data.
Mountain range relative isolation was estimated by the shortest separating distance at the elevation of 2,000 m asl, and by key saddle (the highest touch point) in meters above sea level. Species diversity was expressed as the gamma index which corresponds to the total number of species occurring in a given mountain range, and in the “potential altitudinal transect index” (PAT) which corresponds to the number of sympatric and parapatric species occurring along an elevational gradient (Pyrcz et al., 2013; Pyrcz & Wojtusiak, 2002). Endemism ratio was estimated as the percentage of endemic species and subspecies (PE) within a given mountain range. The variation of the number of endemic species in relation to Prominence was also evaluated. Faunal similarity was calculated by means of the Jaccard coefficient J = A+B/A-B. A linear regression analysis was performed for 2 independent variables (distance and maximum contact elevation) with the species richness between isolated mountain ranges.
The following abbreviations and collection acronyms are applied. FW: Forewing; HW: hindwing; V: ventral surface; D: dorsal surface; SP: Sierra de Perijá, SNSM: Sierra Nevada de Santa Marta, LC: Cordillera de la Costa in Venezuela, CM: Cordillera de Mérida, ST: Sierra de Turimiquire, ET: El Tamá range, EC: Colombian Eastern Cordillera.
The main Andean chain splits in southern Colombia in an area known as the Nudo de los Pastos, into 3 roughly parallel ridges known as Western, Central and Eastern Cordilleras. The latter is the largest of all, with approximately 130,000 km2 above 2,000 m, extending in SN direction over nearly 1,000 km in its northern portion it splits into the Sierra de Perijá following northwards, and the Cordillera de Mérida which detaches into NE direction. The Sierra de El Tamá is geographically a part of the Colombian Eastern Cordillera of the Andes. It is a small compact range somewhat set apart from the main cordilleran stem, in its NW extremity, on the Venezuela - Colombia border. The highest elevation is the Pico El Cobre at 3,615 m asl.
The Sierra del Perijá, comprising the Serranía de Valledupar and the Serranía de Motilones, extends roughly over 300 km in NS direction, and in the widest part it is only 70 km wide. It starts near Ocaña and marks the limit between the Colombian departments of Norte de Santander and Cesar, and then as the range progresses north, it forms the international boundary between Venezuela (Zulia State) and Colombia (Department of Cesar). The highest point is Cerro Tétari at 3,630 m asl, followed by Cerro Irapa at 3,540 m asl.
The Cordillera de Mérida is separated from the El Tamá and the Cordillera Oriental by the valley of the Uribante River, an affluent of the Apure and the Táchira rivers, this last belongs to the Lake Maracaibo basin, in the area called the Táchira Depression. It extends over 450 km in the SW-NE direction, and is 70-90 km wide. It is composed of 3 higher massifs, the southern, Batallón reaching 4,000 m asl; the northern Cendé, which culminates at 3,650 m asl; and the central high block split into 2 parallel ranges by the valleys of Chama and Santo Domingo rivers, the northern La Culata culminating at the Paramo de Piedras Blancas at 4,650 m asl; and the southern Sierra Nevada, with the Pico Bolívar at 4,981 m asl (according to some sources 5,007 m asl), and other peaks exceeding 4,500 m asl. The northernmost extension is the somewhat isolated Sierra de Aroa, culminating at 1,980 m asl.
The Cordillera de La Costa is the northeasternmost extension of the Andes, also called the Maritime Andes, even if most scientists do not consider it as part of the Andes due to a different geological history. It extends for about 450 km and runs roughly parallel to the Caribbean coast. It is separated from the Cordillera de Mérida by the Valley of Chivacoa in the Venezuelan Yaracuy State. It splits into 2 parallel ranges, the higher but narrower, Serranía del Litoral, culminating at 2,710 m asl at the Pico Naiguatá, extending over some 25-30 km NS, and the lower but also wider; and Serranía del Interior, with only 1 peak exceeding 1,500 m, the Cerro Platillón reaching 1,950 m asl, extending some 40-45 km NS. The Cordillera de La Costa collapses to nearly sea level in the Cariaco Basin, and then re-emerges as the Sierra de Turimiquire-its eastern extremity.
The Sierra Nevada de Santa Marta is located west of the Sierra de Perijá across the Cesar Valley. It is a small, encompassing about 17,000 km2, compact, but very high range, extending over 130 km in the N-S and E-W direction. The highest point of the Sierra Nevada is the Pico Cristóbal Colón at approximately 5,700 m asl, lower than the 5,775 m asl, elevation that is often quoted, in the central part of the range (Table 1).
Table 1 Orographic data for each Massif. El Tamá (ET), Cordillera de Mérida (CM), Sierra de Perijá (SP), La Costa (LC) and Sierra Nevada de Santa Marta (SNSM).
| Massif | Maximun altitude (asl) | Saddle (asl) | Prominence (asl) | Surface (km2) |
| SNSM | 5,775 | 191 | 5,584 | 17,000 |
| CM | 5,007 | 1,050 | 3,957 | 32,500 |
| LC | 2,765 | 310 | 2,455 | 40,000 |
| ET | 3,450 | 1,217 | 2,233 | 480 |
| SP | 3,630 | 1,449 | 2,181 | 46 |
The Cordillera de La Costa and the Sierra Nevada de Santa Marta are isolated from other north Andean ranges by elevations below 500 m asl. The Sierra Nevada is separated from the Sierra de Perijá by the Valle del César River with the highest touchpoint at 180 m asl, and the Cordillera de La Costa is separated from the Cordillera de Mérida in the Abra de Sarare at 350 m asl. The Cordillera de Mérida and the Sierra de Perijá are, on turn, separated from the Cordillera Oriental below 1,500 m asl, with the highest touch points at 1,380 m asl and 1,240 m asl, respectively; the latter west of San Cristóbal in El Alto Pass. The Cordillera de La Costa has no area above 3,000 m asl (Fig. 1).
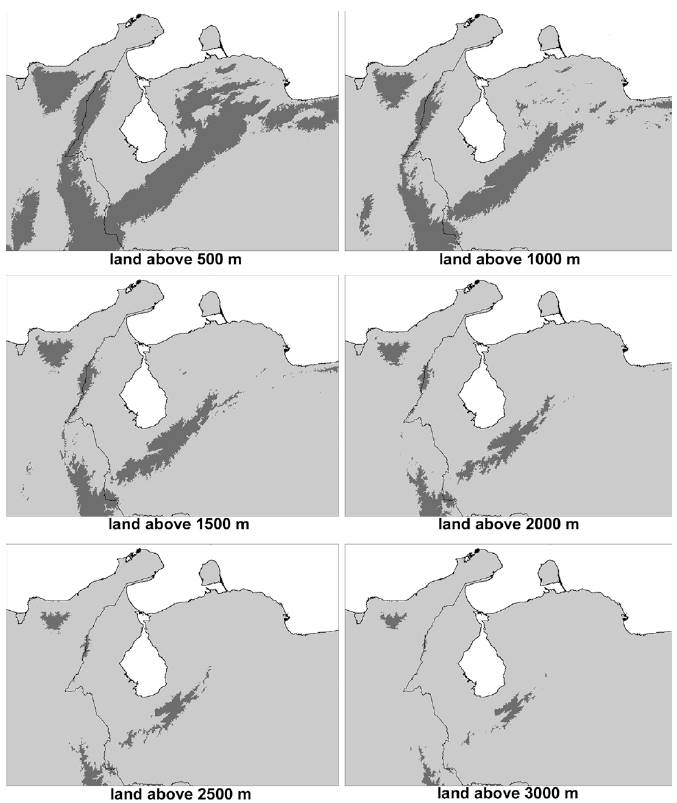
The central Andes (~13° S, 28° S) started to uplift in the Eocene or Oligocene, and in the north the mountains first reached high elevations, progressively extending northwards (Gregory-Wodzicki, 2000; Picard et al., 2008). The initial uplift of the Sierra de Perijá took place in the late Oligocene (ca. 25 mya), almost simultaneously with the orogenesis of the Cordillera de Mérida and the Eastern Cordillera of Colombia (Irving, 1975). The uplift continued in the Miocene and accentuated in the Pliocene, approximately 2 mya (Kellog, 1984) when the range reached elevations even higher than today. During the Pleistocene, the Sierra de Perijá got covered by snowcaps at least in one occasion, especially during the latest Würm glaciation 20,000-23,000 ya (Schubert, 1976). Finally, during the Holocene there was strong erosion coupled with a mild uplift. The Eastern Cordillera only reached half of its present elevation, some 2,000 m asl, at approximately 10 mya. A strong uplift of some 2,000-3,500 m asl has occurred in the Eastern Peruvian Cordillera and the Altiplano in the last 10 million years (Sobolev & Babeyko, 2005). The uplift of the northern Andes was very recent, with these mountains estimated to have been at no more than 40% of their current elevation, in some areas, at 4 mya (Gregory-Wodzicki, 2000). Palynological data indicate that high elevation proto-paramo habitats have appeared in the Cordillera de Mérida only in Late Pliocene or Early Pleistocene (2.5-1.8 mya; Salgado-Labouriau, 1980).
The vegetation areas above 1,000 m asl in the northernmost Andes are covered predominantly by montane vegetation, varying from subdesert to humid forests depending on the slope. Cloud forests make up an extremely varied life zone which occupies a wide elevational band between 1,200/1,400 m asl and 2,800/3,000 m asl. Dryer areas are located on some leeward slopes of some higher mountains. Such dry vegetation pockets are known from the central part of the Cordillera de Mérida in the Timotes and Chama valley, and in the northern section of the transitional area between the Cordillera de Mérida and the Sierra El Tamá, the San Antonio valley (also called Táchira Valley or Táchira Depression). The southern slopes of the Cordillera de La Costa are also considerably dryer, and particularly the Serranía del Interior which is mostly covered by deciduous forests and lower montane shrubland.
The vegetation covering the lands above 2,800 m asl in 4 of the 5 ranges discussed here, form the oreal biome called paramo (Rangel, 2007): Andean subparamo (2,800-3,200 m), the ecotone between the high Andean forest and the open vegetation of the paramo; low paramo (3,200-3,500 m) characterized by shrubs and grasslands; paramo (above 3,400 m asl) characterized by landscapes dominated by Poaceae (Calamagrostis), "frailejonales" (Espeletia), and "chuscales" (Chusquea bamboo) where the community diversification is the highest. In Perijá the paramo strip is much reduced, present only in Cerro Las Tres Tetas and Sabana Rubia. Large extensions of paramo are found through the Cordillera de Mérida, and the Sierra Nevada de Santa Marta. The Cordillera de La Costa is not high enough and is the only range discussed here which lacks altogether the paramo life zone. The top elevations of its highest peaks, the Naiguatá and Oriental, marginally contain pockets of subparamo vegetation with Libanothamnus (Asteraceae), a genus characteristic of this biome.
The northernmost Andes are arguably the best known as far as the systematics of the Panandean subtribe Pronophilina is concerned, as well as the geographic distribution patterns, including altitudinal. Its study began in the 19th century and an important number of faunal monographs and taxonomic revisions have been published in the last 4 decades. First montane South American Satyrinae of the subtribe Pronophilina were probably collected and described from the Venezuelan Cordillera de La Costa. It wasn’t, however, until recently that the fauna of this range was monographed comprehensively (Viloria et al., 2010). This is a relatively species-poor area. It is well known, particularly the Serranía del Litoral. Some isolated areas of the Serranía del Interior require more sampling, which is however not expected to alter the catalogue of species.
The Cordillera de Mérida is more complex both biogeographically and ecologically than any other north Andean range. The first faunal monograph of Adams and Bernard (1981) was restricted in scope to its central part, the Sierra de la Culata and the Sierra Nevada. Numerous other papers followed, dealing with particular taxa, such as Diaphanos (Viloria, 1994), Steromapedaliodes (incl. Redonda; Pyrcz et al., 2017; Viloria & Pyrcz, 2001; Viloria et al., 2013), Lymanopoda (Pyrcz, Casner et al., 2009; Pyrcz et al., 2010) or Pedaliodes (Pyrcz, 2008; Pyrcz & Viloria, 2009), or with restricted geographical areas (Pyrcz & Garlacz, 2012; Pyrcz & Wojtusiak, 2002). The CM has been sampled extensively by the last author of this paper and collaborators from 1991 onward. Despite the many papers dedicated to the Pronophilina of the CM, and a respectable general knowledge, undoubtedly superior to that in the case of any other Andean range, its fauna has not been monographed so far.
The Sierra El Tamá has been sampled for Pronophilina butterflies from the 1980´s onward, more frequently at the break of XXI century, particularly from the northern, Venezuelan side (Pyrcz, 1995). There is only 1 published paper dedicated to its fauna which is a comprehensive monograph (Pyrcz, 1995; Pyrcz & Viloria, 2007). Posterior sampling did not reveal the presence of additional taxa. It is however plausible that other species will be found to occur with more intense sampling.
The monumental snow peaks of the Sierra Nevada de Santa Marta visible form the coast had been attracting a great deal of interest and were fairly accessible from the ports of Santa Marta and Cartagena. As a result, butterfly checklists were available in the 19th century and the first group of species of Pronophilina were described by Godman & Salvin (1880). In the first decades of the 20th century, the SNSM remained in the focus of lepidopterists. It is worth mentioning the research by Eugen Krüger (1924, 1925), who described numerous new endemic taxa of Pronophilina including the paramo inhabitants Paramo oculata (Krüger, 1924) and Lymanopoda nevada (Krüger, 1924). The fauna of the SNSM was monographed by Adams and Bernard (1977) and since then, despite rather frequent sampling (C. Prieto, F. Montero, G. Rodríguez, O. Mahecha, and others), no other new species have been found to occur in this range.
Compared to the neighbouring mountain ranges, the Sierra de Perijá has received little or virtually no attention from the early naturalists. It wasn´t until the long-term research project on Colombian and Venezuelan Pronophilina that the butterflies of the SP remained almost completely unknown (Adams & Benard, 1979). In recent decades, the research on the fauna of SP has been seriously hampered by the social instability of this region related to intense smuggling, guerilla warfare and notorious borderline problems. It was in the early 1990’s that Angel Viloria (1990) undertook for the first time the study of cloud forest butterflies of the SP from the Venezuelan side, with a focus on Satyrinae. His study resulted in the finding of 10 out of previously reported species and discovering 1 new taxon of the genus Oxeoschistus Butler, 1867. At the break of this century, the SP was sampled repeatedly from the Colombian side (Pulido & Andrade, 2007). As a result, a first tentative checklist of butterflies of the SP was published. It was comprised of 86 species including 26 Pronophilina, some of which were new for the range, including 5 species of Pedaliodes which were not identified. Finally, in 2011 a sampling expedition was organized by the IVIC Caracas lead by the last author of this paper. It focused in the northern part of the range west of the locality of Villa del Rosario and was limited to an altitudinal band comprised between 1,400-2,000 m asl.
Results
The robustness of faunal data presented here is to be estimated as high owing to thorough and long-term sampling, and exhaustive systematic research of Pronophilina in the 5 ranges herewith studied. We estimate that the chances of detecting any new species belonging to this group in the LC, SNSM and CM are low. The species inventory of the SP and ET is possibly not complete, and we speculate, based on the knowledge of the neighbouring Colombian Eastern Cordillera, there are possibly 2-4 further species to be found in each range, which however would not alter significantly the results of this study.
Species diversity in the mountain ranges researched in this study, irrespectively if expressed by within-areas measure -gamma diversity- or along potential altitudinal transects (PAT), brings similar results except for the CM (40 and 46 respectively; Table 2). The CM is a significantly larger mountainous chain, compared to the SP, SNSM and LC, with a complex zoogeographical pattern of closely related allopatric species replacing each other in different parts of the range including 9 species of Steromapedaliodes and 4 of Diaphanos. A proportion of genera in the faunas of the researched ranges are closely similar, which is reflected in the ratio of the most speciose genera in the sample. Particularly, compared to ET, which is considered as the “continental source area”, the faunal structure is balanced indicating that the faunas of the 4 peripheral “island” areas are at equilibrium (Appendices 1, 2).
Endemism ratio is higher in the most isolated areas. In the SNSM, it is considerably higher than in other north Andean ranges (PE: 0.59) and marked by the presence of 1 endemic genus (Paramo) and 1 nearly endemic genus (Arhuaco). The endemism ratio at the species level is slightly higher in the SP (PE: 0.32) than in CM (PE: 0.25). However, there are 4 endemic genera in the CM (Steromapedaliodes, Cheimas, Diaphanos), compared to only 1 in the SP (Dangond). The endemism ratio in the LC (PE: 0.25) is similar to that of the CM, but all the endemic species occurring there are closely related, geographical vicariates of the species found in the CM, and there are no endemic genera in the LC (Table 2).
Table 2 Gamma diversity, species richness along potential altitudinal transects (PAT) and endemism (PE) expressed as the proportion of endemic species and subspecies (El Tamá (ET), Cordillera de Mérida (CM), Sierra de Perijá (SP), La Costa (LC) and Sierra Nevada de Santa Marta (SNSM).
| ET | SP | CM | SNSM | LC | |
| S (gamma) | 55 | 39 | 46 | 22 | 24 |
| PAT | 55 | 40 | 40 | 22 | 24 |
| PE sp. | 0.13 | 0.32 | 0.25 | 0.59 | 0.25 |
| PE sp. + ssp. | 0.36 | 0.35 | 0.40 | 0.72 | 0.54 |
The decrease in species richness between the ET and the SP (29%) is comparable with that between the ET and the CM (27%), despite the 3 time longer distance separating the former 2 ranges. The highest contact point of the ET and the SP is however some 300 m higher than that of the ET and the CM. Compared with the CM, the faunal loss of the LC is 40%, slightly lower than that between the SP and the SNSM (44%), despite the 3 times longer distance between the former ranges. Again, the highest touch-point is 200 m higher between the CM and the LC than between the SP and the SNSM. In the latter case, we need to consider the lower ecological diversity of the LC, and the lack of higher elevations (above 2,800 m asl), and thus, the absence of paramo habitats. This restricts the chances of current successful colonization for high elevation species. When subparamo/paramo specialists (combined 11 species) are removed from the analysis, we obtain a significantly lower faunal loss between the CM and the LC, by only 17% (Table 3).
Table 3 Main geographical variables and faunal loss relative to source area (*corrected, species occurring above 2,800 m removed). El Tamá (ET), Cordillera de Mérida (CM), Sierra de Perijá (SP), La Costa (LC) and Sierra Nevada de Santa Marta (SNSM).
| Highest touch point / m | Distance at 2000 m / km | Fauna loss in % | |
| ET - SP | 1380 | 130 | 29 |
| ET - CM | 1250 | 40 | 27 |
| CM - LC | 350 | 180 | 40(17*) |
| SP -SNSM | 190 | 60 | 44 |
Twenty-three species are shared by the SP and the ET. All of them occur at 2,000 m asl or below, and except for C. Corderopedalioides luzangelae Pyrcz & Warren-Gash, 2013, they are also found in the CM. None of the species occurring in the SP at or below 1800 m asl is endemic. They are all widespread Andean species, and only a few differentiated into local endemic subspecies. On the other hand, only 4 species whose lowest altitude range is 2,000 m asl are not endemic in the SP. Two species, however, developed endemic subspecies. Furthermore, all the species which fly at or above 2,400 m asl in the SP are endemic. Some of the endemic species occurring in the SP and in the CM at 2,400-2,600 m asl have corresponding closely related allies in the ET (Adams & Bernard, 1979). Pedaliodes cesarense Adams & Bernard, 1979, Pedaliodes vallenata Adams & Bernard, 1979, and Pedaliodes zuleta Adams & Bernard, 1979 in the SP, and Pedaliodes ornata Grose-Smith & Kirby, 1895, in the CM are morphologically and genetically (Pyrcz, unpublished COI data) closely related to Pedaliodes reyi Pyrcz & Viloria, 2007 in the ET. All these species are part of a possibly monophyletic P. phaea group (Pyrcz et al., 2013), which is characterized by presenting high cryptic diversity (Díaz et al., unpublished COI data; Mahecha-Jiménez et al., 2011). Moreover, this group is found in high elevation cloud forests throughout the northern Andes but is particularly diverse in the Colombian Eastern Cordillera, where in addition to the polytypic Pedaliodes phaea (Hewitson), P. phaeina Staudinger, P. fuscata (Felder & Felder), P. valencia Pyrcz & Viloria, P. haydoni Pyrcz & Viloria and P. bernardi Adams also occur (Pyrcz et al., 2013).
According to our studies, below 2,600 m asl there are no endemic species neither in the SP nor in the CM. However, there are several, apparently paleoendemic elements in the SM which are not related to any species from the latter 2 ranges and presuppose a more remote origin as Paramo oculata (Krüger, 1924) and Lymanopoda nevada E. Krüger, 1924 (Pyrcz et al., unpublished data). Particularly, Arhuaco ica Adams & Bernard (1977) is interesting as it is placed in a genus alongside 1 Central American species, Arhuaco dryadina (Schaus, 1913) (Lamas et al., 2004). However, its phyletic relations remain a matter of controversy due to the incongruence of morphological and partial molecular (COI) data Pyrcz, Lorenc-Brudecka, Zubek et al., 2018). On the other hand, all local endemic species are closely related, sister-species of the species occurring in the CM. From among 11 species all endemic to the SP and 10 endemic to the CM, none is known to occur below 2,400 m asl. Also, none of the 4 endemic species in the SP and 6 in the CM (except for a local subspecies Cheimas opalinus cristalina Pyrcz & Boyer, 2018) with, apparently, no close relatives in the ET occurs below 2,800 m asl (Appendices 1, 2).
Species diversity is higher by 25-30% in the ET than in the SP and the CM. In the SP and the CM there was a local radiation among the taxa occurring at higher elevations, above 2,600 m asl, particularly intense in the CM, which contributed to the local diversity. It resulted in the evolution of several allopatric, sympatric and parapatric sister-species (along an elevational gradient), for example: Corades chelonis Hewitson, 1863 - Corades pax Watkins, 1939 and Steromapedaliodes chiquinquirana- Steromapedaliodes albarregas are distributed parapatrically in the main central ranges of the Cordillera de Mérida (Pyrcz & Garlacz, 2012; Pyrcz et al., 2017). Interestingly, there was a local radiation in the SP, which resulted in the speciation of 3 parapatric species of the Pedaliodes reyi stock (P. zuleta, P. cesarense, and P. vallenata; Diaz et al., unpublished COI data). There was probably also a restricted parapatric speciation in the SNSM: P. tyrrheus Godman & Salvin, 1880 and P. cebolleta Adams & Bernard, 1977, as well as allopatric speciation in P. tyrrheus and P. tyrrheoides Adams & Bernard (Adams, 1985, 1986; Mahecha et al., unpublished data). Eventually, distributional data indicate a parapatric speciation in the LC: E. encycla (Felder & Felder, 1867) and E. neocycla Pyrcz & Viloria, 2010 (Viloria et al., 2010).
There is significant correlation between faunal loss and source area (Fig. 2) and the highest touch point between the ranges (Fig. 3). There seems to be no correlation between the distances from the source area among the different ranges included in this test (Fig. 4). Likewise, there is a variation between the PE index and the prominence massif (Fig. 5), as also between species richness with the prominence, and among the surface area of mountain chains with the species richness (Fig. 6 A, B). Finally, there is higher similarity at the species level between the CM-LC and the SP-ET (Table 4).
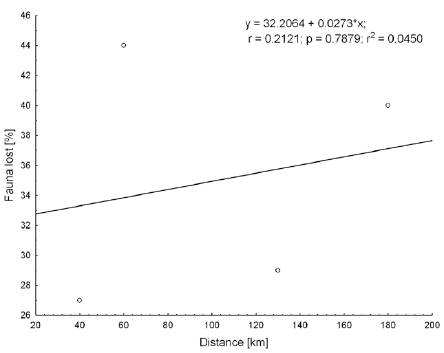
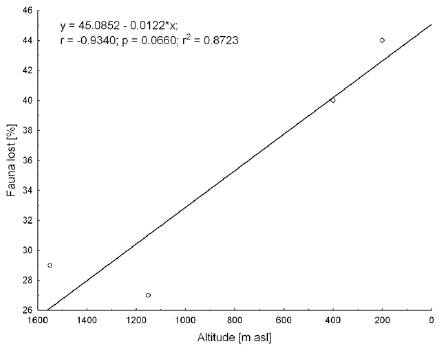
Figure 3 Relationship between faunal loss and maximum touch-point elevation between source and destination area.
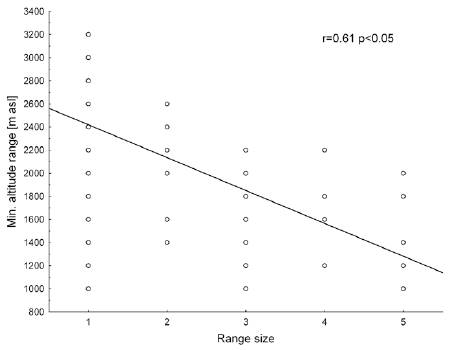
Figure 4 Relationship between range size, expressed as presence in 1-5 mountain ranges of the northern Andes, and altitudinal range.
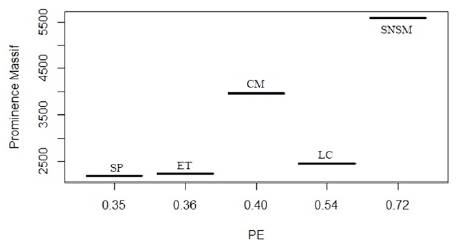
Figure 5 PE index in each Prominence Massif. SNSM (5584), CM (3957), LC (2455), ET (2233), SP (2181).
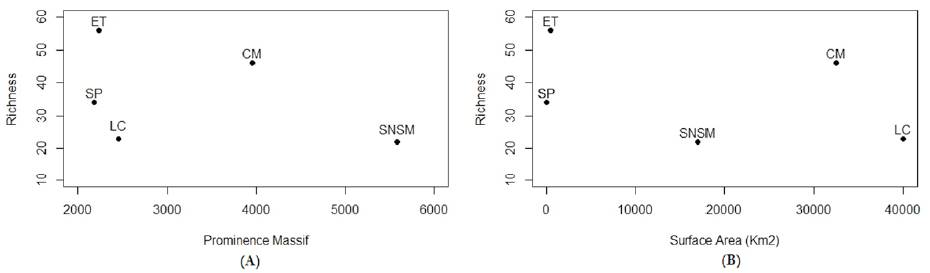
Figure 6 (A), Richness in each Prominence Massif; (B), richness vs. surface (km2). El Tamá (ET), Cordillera de Mérida (CM), Sierra de Perijá (SP), La Costa (LC) and Sierra Nevada de Santa Marta (SNSM).
Discussion
Two key assumptions of this study are that the area of origin of Pronophilina butterflies in the northernmost Colombia and Venezuela is the El Tamá range, and that their dispersal towards the most isolated mountains, the SP, CM, SNSM and LC, was mostly through ecological corridors by slow expansion rather than jump dispersal (Fig. 7). They are supported by the timing of the orogenesis of the northern Andes, especially the uplift of the Eastern Cordillera, paleoecological studies, faunal, and partial phylogenetical data. The northern Andes of Colombia and particularly the peripheral ranges of northern Venezuela and Colombia are considerably younger than the main Andean block (Gregory-Wodzicki, 2000). From Middle to Late Miocene the northern Andes had about half their present average elevation. Particularly, in Colombia the Western and Central Cordilleras already had rather high elevations at the Oligocene-Miocene boundary (24-21 mya; Gómez et al., 2005; Graham, 2009; Nattier et al., 2017). The uplift of the Eastern Cordillera began at that time (Nattier et al., 2017; Restrepo-Moreno et al., 2009), but the present height occurred between 6-3 mya (Graham, 2009) and they probably only attained cloud forest altitudes during the Pleistocene (Nattier et al., 2017). These uplifts and climate change history may have generated the formation of a complex topography expressed by depressions, valleys, rivers, etc., acting as biogeographical barriers for the dispersal of Pronophilinan butterflies through different geological periods, such as the Pliocene-Pleistocene.
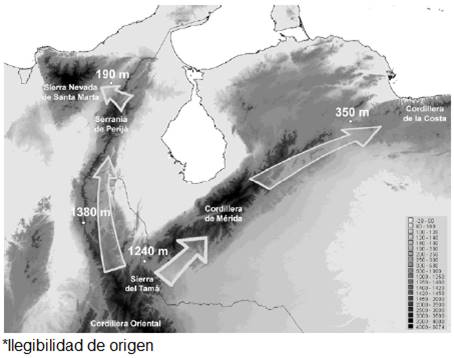
Figure 7 Hypothetical main ways of dispersal of montane Pronophilna butterflies from the main Andes to the peripheral ranges of northern Colombia and Venezuela.
Paleoecological data based on pollen and spore analysis document important climate fluctuations in the northern Andes, for example in the late early Eocene (or Eocene thermal maximum) which was the warmest interval of the Paleogene (Jaramillo & Dilcher, 2000). Furthermore, this event has been associated with increased plant diversity in middle to high latitudes. Increases in greenhouse gases and oceanic heat transport have been proposed to explain this event (Jaramillo & Dilcher, 2000; Sloan et al., 1995). These mechanisms would generate changes in global climate. Therefore, the first would produce more changes in tropical regions (Jaramillo & Dilcher, 2000; Sloan & Rea, 1995). On the other hand, at the Pliocene-Pleistocene (last 5.3 my) involved phases of considerable climate changes like glacial cycles, which produced cooling periods when cloud forests in this region were at lower elevations than today (Flantua et al., 2019; Schubert & Vivas, 1993). Further paleoecological studies in the CM in Venezuela and Eastern Cordillera of Colombia (SP and Norte de Santander) show that paramos underwent robust elevational shifts in vegetation zones during the Pleistocene (2.7-0.02 mya), resulting in extensive changes in surface area and connectivity, led to rapidly changing distributions (an expansion and contraction of the range) of montane species of the fauna and flora, which originated new linages (Flantua et al., 2019; Hazzi et al., 2018; Torres et al., 2013). Present day paramo of Colombia is only 5% of its former zone (Hazzi et al., 2018) and there are records of this ecosystem reaching altitudes as low as 1,900 m asl (Hazzi et al., 2018; Hooghiemstra et al., 2006). Hence, highland mountain ecosystems (e.g., cloud forests and paramos) were connected during glacial periods and successively during interglacial periods, warm climatic conditions fragmented these into isolated (sky) islands, separating the biota specific from them (Flantua et al., 2019; Hazzi et al., 2018). This would explain why the species occurring at approximately 2,000-2,200 m asl in the SP and the CM did not diverge from their relatives in the ET from which they are currently widely isolated and possibly the differentiating effect of the gene pool did not show as yet.
It is possible to identify some temporal patterns in the origin and diversification of Satyrini lineages, indicating that Satyrini originated around 42 mya (41.8 ± 6 mya) during the Eocene. Possibly, the Satyrini underwent a quick diversification in a considerably short span of time, since virtually all the subtribes of the Satyrini, including Pronophilina, appeared during the Oligocene, between 32- 24 mya (Peña et al., 2011). Otherwise, DNA sequences comparative analysis coupled with molecular clocks can be used as another tool for the setting of timing of possible divergence and climatic disturbances (Casner & Pyrcz, 2010; Garzón-Orduña et al., 2014; Marín et al., 2017; Matos-Maraví et al., 2013; Nattier et al., 2017; Peña et al., 2011; Pyrcz et al., 2016; Pyrcz, Lorenc-Brudecka, Zubek et al., 2018). For example, molecular phylogenies of the genus Lymanopoda indicate that the moment of divergence between sister-species Lymanopoda caeruleata Godman & Salving, 1880 and L. caucana Weymer, 1911 (SNSM and SP) took place in mid Pleistocene (~1 mya), whereas between L. dietzi Adams & Bernard, 1981 and L. lecromi Pyrcz & Viloria, 2007 (CM and ET) occurred in late Pleistocene (~0.2 mya; Casner & Pyrcz, 2010). Available faunal and partial phylogenetic data (Casner & Pyrcz, 2010; Pyrcz et al., 2017; Pyrcz, Lorenc-Brudecka, Boyer et al., 2018; Pyrcz, unpubl.) confirm that the faunas of Pronophilina of the Cordillera de Mérida and the Sierra de Perijá originated in El Tamá range, a part of the larger “continental” Cordillera Oriental of the Andes, where most species of the CM and the SP either also occur or have close relatives.
Supported by others studies (Benham et al., 2015; Castroviejo-Fisher et al., 2014; Fiedler & Strutzenberger, 2013; Nattier et al., 2017), we hypothesize that changes in altitudinal distribution of ecological belts associated to Pleistocene climatic variations (after 2.6 mya) may have favoured dispersal of Pronophilina butterfly species between the northernmost Colombian and Venezuelan ranges by creating temporarily existing ecological corridors. Consequently, the dispersal of cloud forest species from older ranges might have been controlled by climate-driven altitudinal shifts of habitats, like those that caused by the glacial cycles (Adams, 1985; Casner & Pyrcz, 2010; Nattier et al., 2017; Pyrcz et al., 2017; Pyrcz, Lorenc-Brudecka, Zubek et al., 2018).
Dispersal from the ET could have been 2 directional, towards the SP and the CM. At the same time, some of the Pronophilina species which succeeded to colonize the SP, might have continued their dispersal towards the eastern slope of the SNSM by overcoming the Cesar valley. A similar scenario is probable between the CM and the LC. The faunas of the SP and the CM are impoverished relative to the source area. There was however a considerable radiation of such genera as Sterompedaliodes and Diaphanos in the CM, and, to some degree, of the genus Pedaliodes in the SP. On their turn, the CM and the SP acted as stepping-stones for the colonization of the LC and the SNSM. In the SNSM there is however, at least 1 taxon (Arhuaco Adams & Bernard, 1977) of different geographic affinity (Central American), suggesting that the fauna of the SNSM is of mixed origin (Pyrcz, Lorenc-Brudecka, Zubek et al., 2018).
Our study shows that the touch-point elevation between the mountain ranges is a better predictor of species affinities and species richness of isolated “continental island” areas than distance. It implies that the dispersal of the studied group of butterflies is taking place through ecological corridors which existed in the past rather than through jump dispersal. Long distance dispersal and overcoming of barriers is a negligible factor in this biogeographic analysis and there is no correlation between distance and the chances of colonization. Worth noting here, is that the above statements refer to this particular study and certainly may not be applied to other biogeographic events involving long distance dispersal.
The analysis of current distribution patterns of the Pronophilina can be a useful tool for identifying the extent of these vertical movements of the vegetation zones and their timing (Casner & Pyrcz, 2010). For example, our study provides indirect proof that the bottom range of cloud forests in most areas of northern Colombia and Venezuela was situated lower, some 600-1,000 m, because none of the species occurring in the SP and the CM above 2,400 m asl is widespread. It appears that the vertical movements of the cloud forests in northern Venezuela (Lara, Trujillo, Aragua, Carabobo) reached some 800-1,000 m lower than today. In fact, several common species to the LC and the CM do not occur currently below 1,400- 1,600 m asl. Such a vertical movement of the cloud forest belt also apparently allowed a very limited dispersal from the SP to the SNSM. These vertical movements, either upwards (canopy/high elevation) or downwards (ground/ low elevation) may afford more plastic ecologies or could provide fitness advantages in the face of environmental instability (Oliveira & Scheffers, 2019; Scheffers et al., 2017). Hence, whereas vertical movements provide opportunities for species to persist by moving within multidimensional climate-habitat space (Oliveira & Scheffers, 2019; Scheffers et al., 2017), higher exposure of regional species pools to climate instability would increase the relative proportion of species in communities towards higher latitudes/altitudes (Oliveira & Scheffers, 2019). Moreover, one possible consequence of steep vertical gradients in forest microclimate is that individuals moving over the vertical gradient on a short-term basis experience a large variation in microclimate as they move. This regular exposure to rapidly varying microclimate should select for greater physiological resilience to short-term climatic variation (Scheffers et al., 2017).
The only species whose distribution does not follow the general pattern observed in this study is Panyapedaliodes panyasis (Hewitson, 1862). This species occurs at elevations above 1,800 m asl, and is nevertheless, distributed throughout all the studied mountain regions without any visible phenotypical variation. It is also widely distributed throughout the Andes (Pyrcz, 2004; Pyrcz & Rodríguez, 2007; Pyrcz & Viloria, 2005). Such a pattern could indicate that in the past the vertical distribution of this species was different, and it was found at lower elevations. This is just a hypothesis which needs to be tested using extensive molecular data for P. panyasis from its entire range to show the patterns of distribution, similar to the phylogeographical study of arachnids by Salgado-Roa et al. (2018). This species, nonetheless, is interesting as it is an exception to a biogeographic pattern presented by other Pronophilina. Interestingly however, P. panyasis does not penetrate to Central America where several north Andean species are found, for example Praepronophila perperna (Hewitson, 1862), Pedaliodes manis (C. Felder & R. Felder, 1867) and Pseudomaniola phaselis (Hewitson, 1862).
Finally, our study indicates that classic island biogeography methodology could be used in the study of the colonization of mountains with, however, some important restrictions because the barriers separating them are not historically permanent in the time. They might “appear” and “disappear” due to changing climatic and ecological factors. The analysis of species distributions allows concluding indirectly on the extent of vertical movements of vegetation zones in the past. It is therefore a valuable tool for paleoecologists.

