











 text new page (beta)
text new page (beta) English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkIntroduction
Animal spatial organization ranges from widely overlapping home ranges to clearly defined exclusive territories owned by strong competitors. A home range can be described as the total area in which the animal lives and reproduces, though it may be undefended, whereas an area is a territory only if it is defended (Burt, 1943; Maher & Lott, 1995). Defended areas can vary greatly in their size and characteristics, ranging from small areas within the home range significant to habitation, reproduction, and feeding (e.g., nests or feeding places), to the entire home range area (Maher & Lott, 1995; Stamps, 1977).
Territorial animals enhance their chances of reproduction and survival by gaining access to mates and excluding potential competitors from their territory. In many lizard species, males are highly territorial (Baird, 2013) and defend their entire home range (Stone & Baird, 2002). In lizards, access to possible mates is often considered the primary motive for defending a territory (Baird, 2013). In such cases males may defend territories that include the home ranges of several females. These females typically mate with the current territory holder (Kiester, 1979; Stamps, 1983, 1987; Tokarz, 1998) and territorial males often have higher reproductive success than non-territorial males within the same species (Baird, 2013; Zamudio & Sinervo, 2003). As such, to ensure paternity of the female's offspring, male lizards will have to exclude potential competitors from their territory. Defense of a territory is costly in terms of both time and energy, and fights can result in injuries (Baird, 2013; Marler & Moore, 1988; Olsson, 1994). Individuals that are stronger, faster and/or healthier have higher resource-holding potential (RHP) and obtain better territories (Andersson, 1994). Often performance traits that determine RHP are related to morphological traits, like size, leg length, head width, etc. (Baird, 2013). To reduce fighting costs, in some species individuals signal their RHP and assess winning chance before engaging in a fight. Signals of high RHP include armaments (Berglund et al., 1996), but can also include ornaments like color (Baird, 2013; Lappin et al., 2006; Whiting et al., 2003).
In this study, we examined the relation between morphological traits and territory quality in Dickerson’s collared lizard, Crotaphytus dickersonae. This is a desert-dwelling lizard, endemic to Mexico, and can only be found in the rocky hills of Isla Tiburón in the Sea of Cortez and in the adjacent mountainous coastline of western Sonora (McGuire, 1996). Males of this species are highly territorial. They emerge from hibernation in March and immediately form territories, as breeding season starts in April (McGuire, 1996), and which they likely maintain during the entire reproductive season (Baird, 2013). This polygynous lizard species displays a strong sexual dimorphism; males are larger, heavier and have wider heads than females (McGuire, 1996; Plasman et al., 2007). Sexual dimorphism in these traits is often explained by sexual selection through male-male competition (Baird, 2013). Crotaphytus dickersonae also displays a strong sexual dimorphism in coloration: Males display a deep cobalt blue color over their entire body while females are a dull brown (McGuire, 1996). This bright coloration over the whole body is exceptional because most animals only display color patches, which in lizards are often only exposed during displays (Andersson, 1994; Whiting et al., 2003). The striking blue coloration of C. dickersonae males makes them highly conspicuous in the open habitat, possibly facilitating assessment of opponents if color functions as an honest signal of RHP. Indeed, an earlier study showed that bluer males had higher running speed and higher bite force and thus likely have higher RHP (Plasman et al., 2015). The exceptional coloration and strong sexual dimorphism of C. dickersonae makes it an excellent model for sexual selection studies. Yet, the species is poorly studied. Here, we examined whether the male's color and several morphological measures of Dickerson’s collared lizard are related to the quality of the territory obtained by the male.
Materials and methods
We studied adult Crotaphytus dickersonae in the coastal hills close to the village Bahía de Kino in Sonora, Mexico. Our study took place from May 2011 until September 2011. The focus of our study involved 2 sites. The sizes of the study sites were approximately 9.0 and 2.7 hectares. Both study sites were situated at a slope of a hill and consisted of granite rock, with some big rocks and many stones, and with little vegetation (i.e., Jatroph cuneata, Fouquieria splendens, Pachycereus pringlei). We caught 95% of all observed collared lizards, 21 lizards (10 males and 11 females) at study site 1 and 15 lizards (4 males and 11 females) at study site 2. We assumed the lizards of these study sites to be of the same population, given that the study sites were located in proximity (< 400 m) and only separated by a piece of flat desert land without geographic interruptions (i.e., no road, river, building, etc.). Additionally, habitat measures of both study sites were similar, giving us the confidence to combine relevant data regarding lizards at both study sites.
Lizards were caught using a pole and a noose made of fishing line. After capture we took a photo of the lateral sides of each lizard’s body to allow for photo identification. Crotaphytus dickersonae have white dots on their body and the pattern and distribution of these dots is unique for each individual lizard (Faber et al., 2013). To allow identification from a distance, we gave each lizard 4 small dots of paint on the dorsal base of the tail in a unique combination of green, red, and yellow non-toxic acrylic paint. When lizards had shed their skin, and thus the paint, they were caught again, identified by their photos, and repainted.
Phenotypic traits were measured on the event of the lizard’s first capture. We measured snout-vent length (SVL; ± 1 mm) and tail length (from the cloaca to the tip of the tail; ± 1 mm) with a flexible ruler. Body mass was measured with a Pesola scale (± 0.5 g). We measured the length of the left lower hind limb (from the knee joint to the ankle joint) with a caliper (± 1 mm). Head width of each lizard was measured with a caliper (± 1 mm) at the widest point of the head where the lateral extent of the jaw adductor muscles was largest. Body condition index (BCI), often used as an indication of the general physical status of the lizard, was estimated as the standardized residuals of the linear regression of body mass on SVL (Schulte-Hostedde et al., 2005).
We took a measurement of the blue color on both lateral sides of the dorsum of the male lizards with a portable spectrophotometer (MINOLTA CM-2600d, Minolta Co. Ltd., Osaka Japan). Blue chroma was calculated as reflection from 400-480 nm divided by the total reflection measured by the spectrophotometer (360-740 nm). In subsequent analyses we used the average blue chroma of both measurements. Blue chroma was determined for 7 males. For 6 of them we had information on territory variables. For these 7 males, color was measured in the same week and around the same time (16:00 h) after the males had been allowed to sunbathe in an enclosure consisting of a wooden frame covered with mesh (60 × 60 × 60 cm).
One of the male lizards we caught had lost one of its forelimbs and part of his tail. We decided to include this lizard in our study because we considered its short tail would affect his quality similarly as a naturally shorter tail would and his SVL, body mass, length of the hind leg, head width, and blue chroma did not fall outside the range of the measurements in other males.
Stone and Baird (2002) found that male collared lizards defend their entire home range against conspecific males. We therefore treated the home ranges of the males in our study as their territory, as has been done in other studies on collared lizards (Baird et al., 2007; Husak et al., 2008). Territories of males and home ranges of females were determined based on point sightings collected from May 3 until August 23 during daily walks through the study sites in a routine way between 8:00 and 17:30 h. We recorded the exact location of lizards with a Garmin GPSmap 62s (< 3 m accuracy). When a lizard was encountered twice during a day, we only recorded the second sighting when at least 2 hours had passed since the first sighting. We used a minimum of 15 sightings to calculate territory and home range size with the minimum convex polygon technique (Rose, 1982), the only exception to which was a lizard for which we used only 12 sightings to calculate territory size. This lizard had not been seen after July 16 and we assumed that he had died. This was the same lizard that had lost his forelimb and part of its tail. We collected enough sightings to estimate territory size for 6 males and home range size for 10 females.
We determined the home range of females for which we had a minimum of 15 sightings. We recorded 1) the number of overlapping female home ranges for each male territory, and 2) the total number of females that were seen in a male’s territory, including those females for which we did not calculate home range size.
We determined the exclusivity of a territory by 1) the number of overlapping male territories, 2) the percentage of overlap with territories of other males, and 3) the number of other males seen in the territory that were observed fewer than 15 times.
A higher number of refuges and basking sites may indicate higher territory quality. Collared lizards may hide below trees and shrubs from possible predators. Furthermore, the shade of these plants allow for thermoregulation opportunities. Large rocks are the preferred basking sites of males (McGuire, 1996). For each territory, we counted the number of trees and shrubs, and rocks taller than 30 cm to calculate the number of refuges and basking sites per m2, respectively.
We recorded all sightings of potential predators of C. dickersonae for our assessment of territory quality. The most dangerous terrestrial predators of Crotaphytus species are considered to be the roadrunner (Geococcyx californianus, a terrestrial bird) and the coachwhip (Masticophis flagellum, a diurnal snake) (Husak, Macedonia et al., 2006). Crotaphytus species are also predated by flying avian predators, but given that their hunting areas easily encompass the total area of both the study sites, making them an equal threat to all of the lizards in our study, they were not included in our consideration of territory quality. Furthermore, during our study we observed only 1 potential terrestrial predator of C. dickersonae (a rattlesnake species), therefore we excluded predator abundance of the territory quality assessment.
Territory and home range analyses (size and overlap) were performed with GIS and ArcView 3.2a with the Convex Hulls v. 1.24 extension by Jenness Enterprises (Jenness, 2008). Statistical analyses were performed in R, version 3.3.3 (R Core Team, 2017). We used non-parametric tests: Mann Whitney U tests to evaluate differences between males with and without territories, and Spearman rank correlation tests to assess correlations between morphological traits and aspects of their territories.
Results
At study site 1, we collected enough sightings to plot the territories of 5 males and the home ranges of 5 females (Fig. 1). We saw 5 additional males (each observed between 1 and 6 times) and 6 additional females (with sightings ranging from 1 to 10).
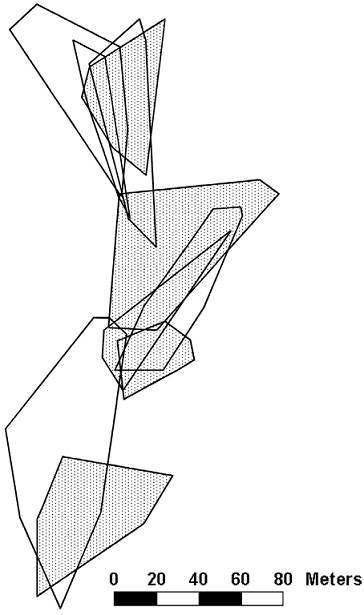
Figure 1 Territories of male (open) and home ranges of female (dotted) Crotaphytus dickersonae at study site 1.
Additionally, we observed a bout of interactions between 2 males at study site 1 on June 4, resulting in the apparent expulsion of one male from its territory. One of the males ran from a distance of > 30 m to another male and chased him away repeatedly. After June 4, the lizard that had been chased away was not seen again at the coordinates at which the chase had taken place, though he did establish a new territory within our study site (Fig. 2). For our calculations of territory quality and territory overlap of this lizard, we only used the sightings that we collected after June 4.
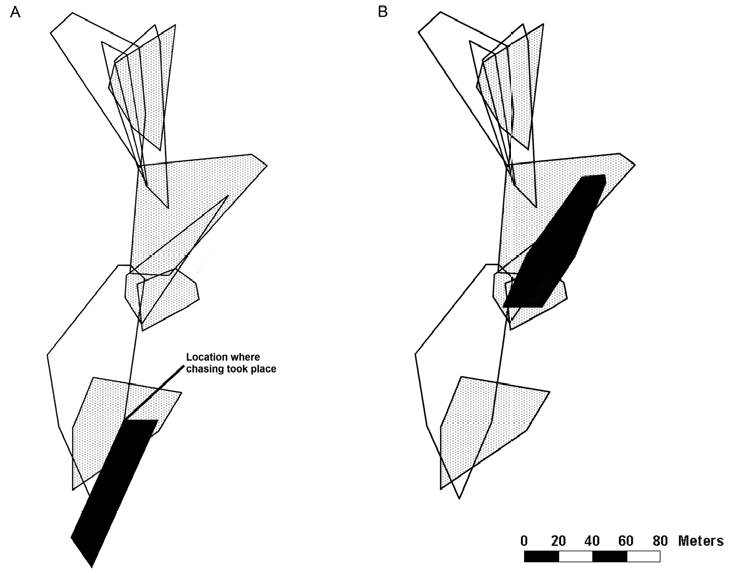
Figure 2 Change of territory (black) before (A) and after (B) the owner of the territory was chased away by the male of the overlapping territory. Territories of other males are indicated by open areas; home ranges of females are dotted.
At study site 2 we collected enough sightings to plot the territory of 1 male and home ranges of 5 females (Fig. 3). We saw 3 additional males (each observed between 1 and 3 times) and 6 additional females (each observed between 1 and 6 times).
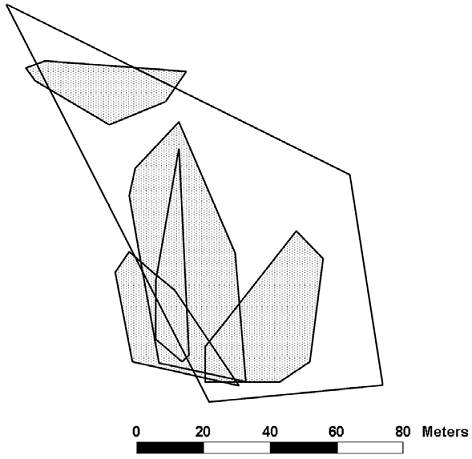
Figure 3 Territory of male (open) and home ranges of female (dotted) Crotaphytus dickersonae at study site 2.
Phenotypic traits did not differ between territorial males (> 12 sightings, n = 6) and other males (≤ 6 sightings, n = 10; Mann-Whitney U tests: U > 23.00, p > 0.44; Table 1). Males with higher SVL had longer tails (rs= 0.83, p= 0.04), wider heads (rs = 0.90, p = 0.01), and longer legs (rs = 0.99, p = 0.0003). Strangely, SVL did not relate with body mass (rs = 0.60, p = 0.21). Heavier males tended to have longer tails (rs = 0.77, p = 0.07) and wider heads (rs= 0.75, p = 0.08). When corrected for SVL, body mass, tail length, head width, and leg length were uncorrelated (rs < 0.66, p > 0.16). For further analyses we used morphology traits corrected for SVL. Bluer males had wider heads (rs= 0.93, p = 0.01), and tended to be heavier (rs = 0.77, p= 0.07), but blue chroma did not correlate with SVL (rs= 0.71, p = 0.11) or tail length (rs = 0.66, p = 0.16).
Table 1 Morphological traits of territorial and non-territorial males. Average ± SD are given. Differences were not significant.
| Males | ||
| with territory | without territory | |
| SVL (mm) | 84 ± 8 | 84 ± 8 |
| Weight (g) | 23 ± 8 | 28 ± 11 |
| Head width (mm) | 20 ± 5 | 22 ± 3 |
| Tail length (mm) | 187 ± 39 | 197 ± 14 |
| Hind limb length (mm) | 29 ± 5 | 30 ± 3 |
Male territory size averaged 2,912.9 m2, ranging from 693.5 m2 to 6,236.0 m2 (n = 6) (Figs. 1, 3). We found no significant correlations between phenotypic traits and the size of the territory (Table 2).
Table 2 Spearman correlations between territory aspects and morphological traits of the male lizards. Results are given with and without correcting for territory size. Tail length and head width were corrected for SVL. Body mass and hind limb length were excluded from the table, as they did not correlate with any of the territory aspects. · indicates a trend (0.10 < p > 0.05), * indicates p < 0.05, and ** indicates p < 0.01.
| Territory quality | SVL | Tail length | Head width | Blue chroma | ||||
| rs | p | rs | p | rs | p | rs | p | |
| Territory size: | 0.54 | 0.27 | 0.60 | 0.21 | 0.60 | 0.21 | 0.66 | 0.16 |
| Female presence: | ||||||||
| Nr of female's home ranges overlap | 0.76 | 0.08˙ | 0.94 | 0.01** | 0.76 | 0.08˙ | 0.76 | 0.08˙ |
| corrected for territory size | -0.03 | 0.70 | -0.26 | 0.62 | -0.26 | 0.62 | -0.26 | 0.62 |
| Additional females | 0.83 | 0.14 | 0.93 | 0.01** | 0.74 | 0.09˙ | 0.74 | 0.09˙ |
| corrected for territory size | 0.94 | 0.36 | 0.76 | 0.08˙ | 0.76 | 0.08˙ | 0.76 | 0.08˙ |
| Total nr of females | 0.76 | 0.08˙ | 0.94 | 0.01** | 0.76 | 0.08˙ | 0.76 | 0.08˙ |
| corrected for territory size | 0.60 | 0.40 | 0.49 | 0.33 | -0.30 | 0.96 | 0.14 | 0.79 |
| Exclusivity: | ||||||||
| Nr of male territories | -0.68 | 0.14 | -0.93 | 0.01** | -0.74 | 0.09˙ | -0.74 | 0.09˙ |
| corrected for territory size | -0.60 | 0.21 | -0.77 | 0.07˙ | -0.77 | 0.07˙ | -0.77 | 0.07˙ |
| % Overlap male territories | -0.54 | 0.27 | -0.83 | 0.04* | -0.83 | 0.21 | -0.66 | 0.16 |
| Additional males | 0.77 | 0.07˙ | 0.83 | 0.04* | 0.93 | 0.01** | 0.93 | 0.01** |
| corrected for territory size | 0.52 | 0.30 | -0.60 | 0.12 | 0.88 | 0.02* | 0.88 | 0.02* |
| Habitat: | ||||||||
| Refuges (per m2) | -0.49 | 0.33 | -0.43 | 0.40 | -0.60 | 0.21 | -0.54 | 0.27 |
| Basking sites (per m2) | 0.14 | 0.79 | 0.31 | 0.54 | -0.20 | 0.70 | -0.14 | 0.79 |
The average number of overlapping female home ranges for each male territory was 3 and ranged from 2 to 5 (Figs. 1, 3). Males with longer tails had more overlapping female home ranges (rs = 0.94, p = 0.005, Fig. 4). Males with longer SVL, wider heads, and higher blue chroma tended to have more overlapping female home ranges (Table 2). However, when correcting for territory size all these correlations became non-significant (rs < 0.26, p > 0.623). There was no correlation between hind limb length and the number of overlapping female home ranges.
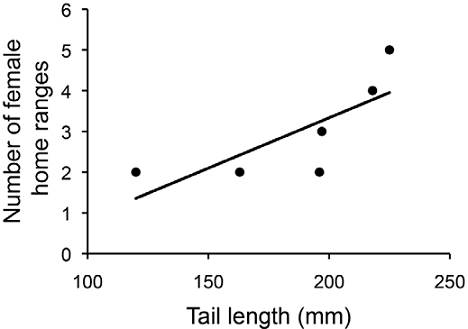
Figure 4 Relationship between the number of overlapping female home ranges and the tail length in male Crotaphytus dickersonae.
The average of the total number of females observed in a male’s territory (including the females for which we did not calculate home range size) was 3.8 and ranged from 2 to 8. The total number of females observed in a male’s territory was significantly higher for males with longer tails (rs = 0.94, p = 0.005) and tended to be higher for males with longer SVL, wider heads, and higher blue chroma (Table 2). We found no correlation between the total number of females seen in a male’s territory and the hind limb length or body mass of the male. When correcting for territory size, no morphological measure bore statistical relation to the total number of females observed in the territory (p > 0.20). Interestingly, the additional females, with too few sightings to calculate their home range, were only seen in the territories of the males with the widest heads and the longest tails (Fig. 5).
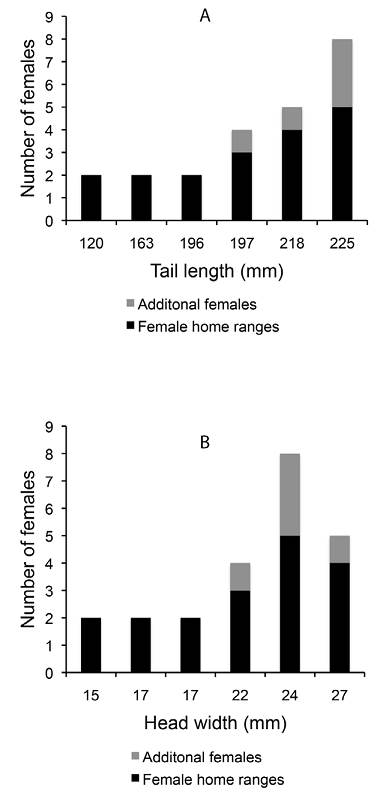
Figure 5 The relationship between the total number of females that was seen in a male’s territory and the tail length (A) and the head width (B) of the male in Crotaphytus dickersonae.
The number of overlapping male territories averaged 1.33 and ranged from 0 to 2. Males with longer tails had less overlapping male territories (rs = -0.93, p = 0.008). There was a trend that males with wider heads and higher blue chroma had less overlapping male territories (Table 2). We found no significant correlation between the SVL and the hind limb length of the male and the number of overlapping male territories (Table 2).
The percentage of overlap with other male territories averaged 31.5% and ranged from 0% to 98.7%. Males with longer tails had significantly less overlap with other male's territories (Table 2; Fig. 6). There was no significant correlation between the other phenotypic traits and the percentage of overlap with other male territories (Table 2).
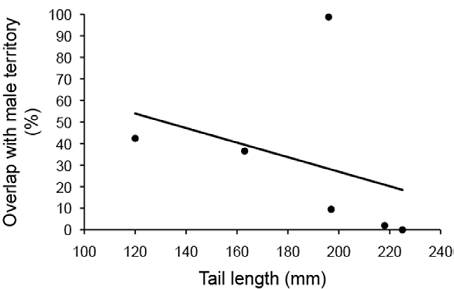
Figure 6 The relationship between the percentage of overlap with the territories of other males and the tail length of male Crotaphytus dickersonae.
Males with longer tails, wider heads, and higher blue chroma had a higher number of additional males (males for which we did not calculate territory size) in their territories (Table 2). Males with higher SVL also tended to have more incidences of additional males in their territories (Table 2). When correcting for territory size, head width and blue chroma still related positively with the number of additional males (Table 2). Interestingly, the number of additional males was correlated with the number of females in the territory (rs = 0.89, p = 0.02).
The number of trees and shrubs in a territory ranged from 150 to 456 and averaged 248. The territories had an average of 0.107 trees and shrubs per m2; this ranged from 0.069 to 0.216. The total number of rocks in a territory ranged from 75 to 526 and averaged 224.5. The number of rocks per m2 averaged 0.080 and ranged from 0.042 to 0.108. When corrected for territory size, the number of refuges or basking sites did not correlate with any of the phenotypic traits evaluated (Table 2). Furthermore, the number of refuges or basking sites did not correlate with the number of females present in the territory, whether considering total number of females, percent of overlap with female home ranges, or additional females (p > 0.11).
Discussion
We studied the relation of phenotypic traits with territory quality in Crotaphytus dickersonae. Results show that males with longer tails had higher female presence in their territory and less overlap with other male territories. Additionally, bluer males and males with wider heads tended to have territories with more females and less overlapping male territories, yet these males had more additional males in their territories.
Tail length and head width are morphological features that can be related to RHP. Longer tails increase agility, allowing the animal to run faster without losing stability (Wheatley et al., 2015; Wynn et al., 2015). Tail length is related to sprint speed in lizards. Experimental reductions in tail length decreased sprint speed in some lizard species (Ballinger et al., 1979; Downes & Shine, 2001; Formanowicz et al., 1990), but increased sprint speed in others (Daniels, 1983; Li et al., 2011). Garland (1985) suggested that longer tails would be especially beneficial for lizards that run bipedally. Crotaphytus species are well known for their bipedal gait (McGuire, 1996), and longer tails may allow them to run faster. Lizards that can run faster have larger territories (Peterson & Husak, 2006) and are more likely to achieve dominance (Garland et al., 1990; Robson & Miles, 2000). This suggests that running speed is important in male-male competition. Accordingly, Husak and Fox (2006) found that male C. collaris use near maximum sprint speed when they respond to other males in their territory. In the interactions between males that we observed, a typical pattern of behavior involved one of the males running quickly to the other male, with the latter subsequently running away. We observed several such interactions between 2 males, which even led to territory replacement of one of the males (see results). The lizard that had chased away the other male was the male with the widest head and longest tail of all males included in this study. Hence, faster and stronger males may be able to maintain exclusivity of their territory. These males might also be better able to expand the boundaries of their territory, possibly increasing the number of females in their territory while still being able to keep other males away from those females.
Biting is often used in agonistic encounters (Hall et al., 2010; Lailvaux & Irschick, 2006) and can be considered an important trait determining RHP (Lailvaux & Irschick, 2006). Male lizards that bite harder are more likely to win fights (Husak, Lappin et al., 2006; Huyghe et al., 2005; Lailvaux et al., 2004), have larger territories with more overlap with female home ranges (Lappin & Husak, 2005), and they have a higher reproductive success (Husak et al., 2009; Lappin & Husak, 2005). Head size and shape are strongly correlated with bite force (Anderson et al., 2008). In male lizards, head width is a predictor of bite force; males with wider heads bite harder (Herrel et al., 2006; Husak, Lappin et al., 2006; Lappin & Husak, 2005). The positive relation between head width and territory quality suggests that biting also plays a major role in territory acquisition and defense in C. dickersonae. Head width, however, correlated positively with additional males. Perhaps males are attracted by the higher territory quality of these males, in which more female encounters may be worth the risk of the attack of the strong territory owner.
We found that bluer males tended to have more exclusive territories. The intensity of the blue color relates to running speed and bite force in C. dickersonae and could thus be an honest signal of RHP (Plasman et al., 2015). Morphological features may not be easily distinguished from long distances, whereas bright coloration can be easily detected (Losos, 1985; Whiting et al., 2003), especially in the open habitat of the Sonoran desert. The blue color may facilitate the assessment of the opponent's RHP over long distances (preliminary opponent assessment), whereas morphological traits may be assessed in a later stage when animals are nearer (secondary opponent assessment; Kokko, 2013). The blue color may thus dissuade other males to invade their territory. Contrarily, however, we observed more additional males in the territories of bluer males. Likewise, more additional males were observed in territories of males with longer tails and wider heads. These additional males may temporarily sneak into these territories with the intention to steal copulations. This strategy is frequently seen in species where part of the population is not able to obtain a territory (Gross, 1996; López-Sepulcre & Kokko, 2005) and has been found to have equal fitness benefits as that of territory owners in the C. collaris (York et al., 2014). The big size of the territories of C. dickersonae prevent males from guarding the entire area simultaneously, leaving parts open to opportunistic males. Further, the number of additional males correlated with female abundance. Interestingly, the strong correlation of number of additional males with blue chroma of the resident male may suggest that the sneaker males perhaps uses the information broadcasted by the territorial male to estimate female presence.
More females were found in the territories of males with longer tails, wider heads and higher blue chroma. Higher female abundance may occur due to high territory quality. Although we did not find correlations with abundance of basking sites and refuges, or the presence of predators, we cannot exclude other territory characteristics that were not measured here (e.g., food abundance). On the other hand, female abundance may be higher, because females prefer the territory owner as mate. Females may be attracted by traits used in male-male competition, as these traits may relate to general condition or quality (Berglund et al., 1996) and provide the female with good genes for her offspring (Hamilton & Zuk, 1982; Zahavi, 1975) or give direct benefits in the form of a high quality territory and protection from other males (Emlen & Oring, 1977). Female choice has rarely been documented in lizards (Olsson & Madsen, 1995; Olsson et al., 2013) and generally the female mates with the territory owner with which her home range overlaps (Kiester, 1979; Stamps, 1983, 1987; Tokarz, 1998). Nevertheless, females may temporarily leave their home range to mate with a high quality male (Calsbeek & Sinervo, 2002; Vitousek et al., 2007). In the collared lizard, C. collaris, almost 40% of the offspring was from a male other than the owner of the territory with which their home range overlapped (Husak et al., 2008) and multiple mating may be common (York & Baird, 2015). However, the lack of difference between offspring fitness from territorial and non-territorial males and the thermoregulation costs associated to fleeing from males, suggests that females may accept copulations to avoid male harassment (York & Baird, 2015). On the other hand, in an earlier study with C. collaris it was found that females discriminated against young males (Baird et al., 1996). Furthermore, we found that only males with long tails and wide heads had additional female visitors, which may suggest female choice. To what extent female choice determines overlap of male territories with female home ranges or additional female visitors remains unclear and is an interesting topic for future studies.
Although we assumed basking sites and refuges to be important factors for survival in C. dickersonae, their quantity did not relate to any male trait or to female presence. Possibly refuges and basking sites are not considered when selecting a territory, due to the high abundance of rocks and bushes in our study site. As such, territory quality was determined primarily by its size and the presence of conspecifics, and obtaining mates is likely the most important motive for territory defense in this species (Baird, 2013).
Our results indicate that, in male Crotaphytus dickersonae, head width (i.e., bite force) and especially tail length (i.e., running speed) are important traits for the intra-sexual contest for territory acquisition and determine access to females for mating. The blue coloration of the males probably signals male dominance status (Plasman et al., 2015) and helps to estimate the opponent's RHP over long distances and possibly attracts females. The conspicuous sexual signals can thus increase fitness benefits (e.g., repel opponents and attract mates). Yet, the higher number of additional males in territories of bluer males suggests that opponents may exploit the signal when intercepting the mates attracted by the signal (Perril et al., 1978). Further, our results indicate that territories of C. dickersonae males are large compared to those of other animals of similar size (Candolin & Voigt, 2001; McLoughlin & Ferguson, 2000), other lizards (Molnár et al., 2016; Stehle et al., 2017), or even other collared lizard species (Schwartz et al., 2007; although this may depend on population, see McCoy et al., 2003). Due to their size, the number of territories that can be established is probably limited, whereas costs for patrolling and defending this big area are likely high, and low female density may result in low reproductive payoff (Briffa & Sneddon, 2007; Lappin & Husak, 2005). Hence, it could be concluded that C. dickersonae is a species with relatively few territorial males and many floaters (López-Sepulcre & Kokko, 2005). It needs to be determined if these 2 mating strategies form an ESS (evolutionary stable strategy) with equal success or that being a floater is 'making the best of a bad job'.

